
Friend or Foe? The Roles of Antioxidants in Acute Lung Injury

 and
and 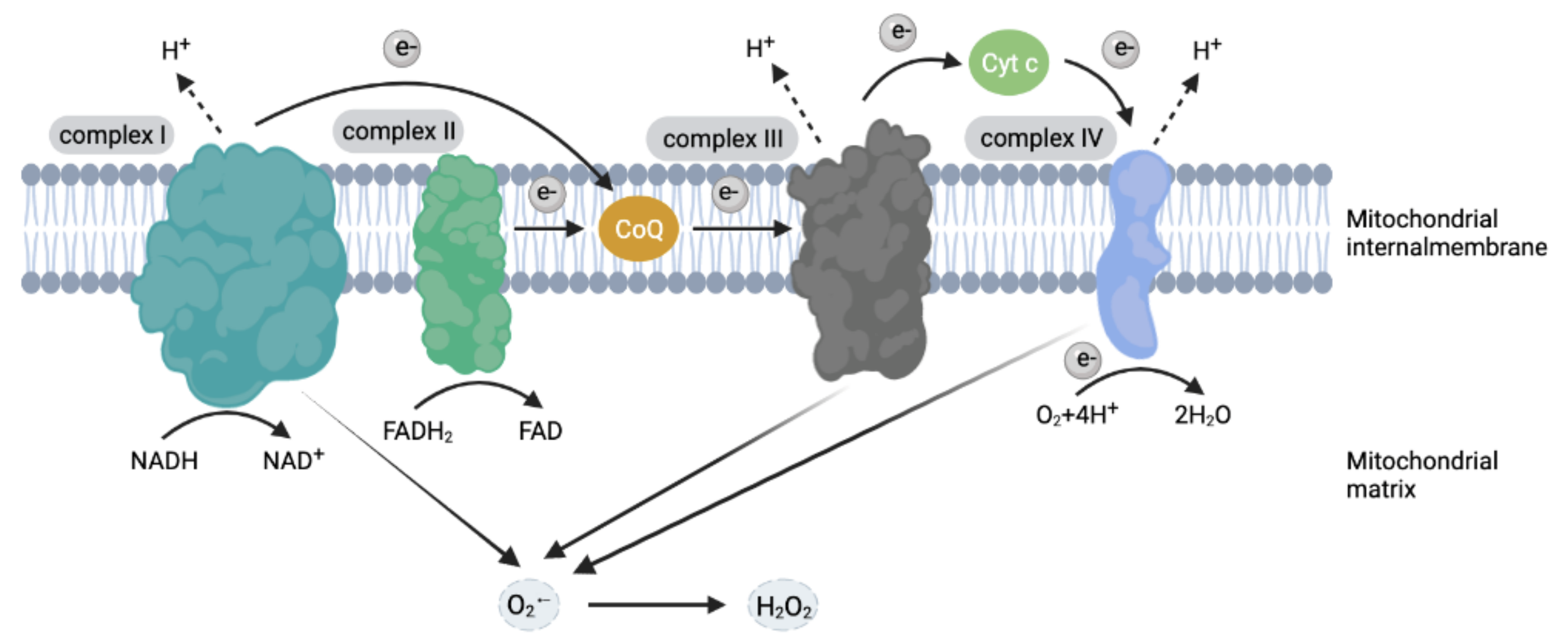{kind=link}
{kind=link}
Abstract
:1. Introduction
2. The Role of ROS in ALI
2.1. ROS Generation
2.2. The Oxidation Function of ROS
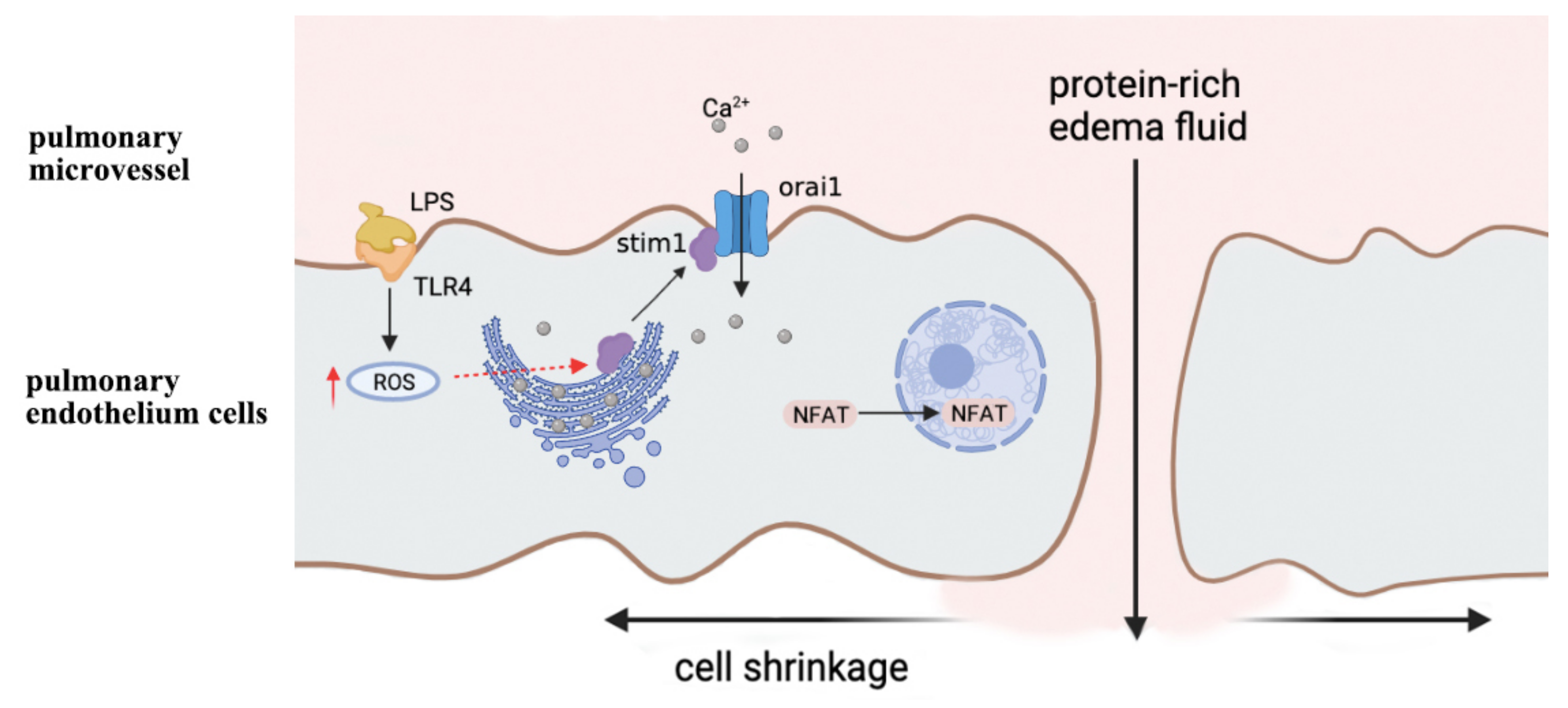
2.3. ROS Induces Calcium Channel Dysfunction
2.4. ROS Induces Chemotaxis of Neutrophils
2.5. Cell-Free Hemoglobin (CFH) and ALI
3. The Application of Antioxidants in ALI
3.1. Acetylcysteine (NAC)
3.2. Vitamins
3.3. Ambroxol
3.4. Natural Medicine
3.5. Micronutrients
4. The Possible Mechanisms of Antioxidants Failure in Severe Organ Injury
4.1. Possible Mechanism 1: The Dosage and Toxicity of Antioxidants
4.2. Possible Mechanism 2: Normal Physiological Function of Free Radicals and Hormesis Theory
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Gattinoni, L.; Tonetti, T.; Quintel, M. Regional physiology of ARDS. Crit. Care 2017, 21, 312. [Google Scholar] [CrossRef] [Green Version]
- Ashbaugh, D.G.; Bigelow, D.B.; Petty, T.L.; Levine, B.E. Acute respiratory distress in adults. Lancet 1967, 2, 319–323. [Google Scholar] [CrossRef]
- Ranieri, V.M.; Rubenfeld, G.D.; Thompson, B.T.; Ferguson, N.D.; Caldwell, E.; Fan, E.; Camporota, L.; Slutsky, A.S. Acute respiratory distress syndrome: The Berlin Definition. JAMA 2012, 307, 2526–2533. [Google Scholar]
- Bernard, G.R.; Artigas, A.; Brigham, K.L.; Carlet, J.; Falke, K.; Hudson, L.; Lamy, M.; Legall, J.R.; Morris, A.; Spragg, R. The American-European Consensus Conference on ARDS. Definitions, mechanisms, relevant outcomes, and clinical trial coordination. Am. J. Respir. Crit. Care Med. 1994, 149, 818–824. [Google Scholar] [CrossRef] [PubMed]
- Fan, E.; Brodie, D.; Slutsky, A.S. Acute Respiratory Distress Syndrome: Advances in Diagnosis and Treatment. JAMA 2018, 319, 698–710. [Google Scholar] [CrossRef] [PubMed]
- Suki, B.; Hubmayr, R. Epithelial and endothelial damage induced by mechanical ventilation modes. Curr. Opin. Crit. Care 2014, 20, 17–24. [Google Scholar] [CrossRef]
- Meduri, G.U.; Bridges, L.; Shih, M.C.; Marik, P.E.; Siemieniuk, R.; Kocak, M. Prolonged glucocorticoid treatment is associated with improved ARDS outcomes: Analysis of individual patients' data from four randomized trials and trial-level meta-analysis of the updated literature. Intensive Care Med. 2016, 42, 829–840. [Google Scholar] [CrossRef] [PubMed]
- Allen, T.C.; Kurdowska, A. Interleukin 8 and acute lung injury. Arch. Pathol. Lab. Med. 2014, 138, 266–269. [Google Scholar] [CrossRef]
- Tschopp, J.; Schroder, K. NLRP3 inflammasome activation: The convergence of multiple signalling pathways on ROS production? Nat. Rev. Immunol. 2010, 10, 210–215. [Google Scholar] [CrossRef]
- Zemans, R.L.; Matthay, M.A. What drives neutrophils to the alveoli in ARDS? Thorax 2017, 72, 1–3. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Wu, D.; Jiang, X. Icam-1 and acute pancreatitis complicated by acute lung injury. JOP 2009, 10, 8–14. [Google Scholar] [PubMed]
- Zhou, T.; Song, W.F.; Shang, Y.; Yao, S.L.; Matalon, S. Halogen Inhalation-Induced Lung Injury and Acute Respiratory Distress Syndrome. Chin. Med. J. 2018, 131, 1214–1219. [Google Scholar] [CrossRef]
- Conte, E.; Fagone, E.; Fruciano, M.; Gili, E.; Iemmolo, M.; Vancheri, C. Anti-inflammatory and antifibrotic effects of resveratrol in the lung. Histol. Histopathol. 2015, 30, 523–529. [Google Scholar] [PubMed]
- Panahi, Y.; Ghanei, M.; Hashjin, M.M.; Rezaee, R.; Sahebkar, A. Potential Utility of N-acetylcysteine for Treating Mustard Lung. Crit. Rev. Eukaryot. Gene Expr. 2017, 27, 247–266. [Google Scholar] [CrossRef]
- Sies, H.; Jones, D.P. Reactive oxygen species (ROS) as pleiotropic physiological signalling agents. Nat. Rev. Mol. Cell Biol. 2020, 21, 363–383. [Google Scholar] [CrossRef] [PubMed]
- Piotrowski, W.J.; Marczak, J. Cellular sources of oxidants in the lung. Int. J. Occup. Med. Environ. Health 2000, 13, 369–385. [Google Scholar]
- Konior, A.; Schramm, A.; Czesnikiewicz-Guzik, M.; Guzik, T.J. NADPH oxidases in vascular pathology. Antioxid. Redox Signal. 2014, 20, 2794–2814. [Google Scholar] [CrossRef] [Green Version]
- Kellner, M.; Noonepalle, S.; Lu, Q.; Srivastava, A.; Zemskov, E.; Black, S.M. ROS Signaling in the Pathogenesis of Acute Lung Injury (ALI) and Acute Respiratory Distress Syndrome (ARDS). Adv. Exp. Med. Biol. 2017, 967, 105–137. [Google Scholar]
- Carnesecchi, S.; Pache, J.C.; Barazzone-Argiroffo, C. NOX enzymes: Potential target for the treatment of acute lung injury. Cell. Mol. Life Sci. 2012, 69, 2373–2385. [Google Scholar] [CrossRef]
- Jin, H.Z.; Yang, X.J.; Zhao, K.L.; Mei, F.C.; Zhou, Y.; You, Y.D.; Wang, W.X. Apocynin alleviates lung injury by suppressing NLRP3 inflammasome activation and NF-kappaB signaling in acute pancreatitis. Int. Immunopharmacol. 2019, 75, 105821. [Google Scholar] [CrossRef]
- Jiang, J.; Huang, K.; Xu, S.; Garcia, J.; Wang, C.; Cai, H. Targeting NOX4 alleviates sepsis-induced acute lung injury via attenuation of redox-sensitive activation of CaMKII/ERK1/2/MLCK and endothelial cell barrier dysfunction. Redox Biol. 2020, 36, 101638. [Google Scholar] [CrossRef]
- Gielis, J.F.; Boulet, G.A.; Briede, J.J.; Horemans, T.; Debergh, T.; Kusse, M.; Cos, P.; Van Schil, P.E. Longitudinal quantification of radical bursts during pulmonary ischaemia and reperfusion. Eur. J. Cardiothorac. Surg. 2015, 48, 622–629. [Google Scholar] [CrossRef] [Green Version]
- Bortolotti, M.; Polito, L.; Battelli, M.G.; Bolognesi, A. Xanthine oxidoreductase: One enzyme for multiple physiological tasks. Redox Biol. 2021, 41, 101882. [Google Scholar] [CrossRef] [PubMed]
- Furuhashi, M. New insights into purine metabolism in metabolic diseases: Role of xanthine oxidoreductase activity. Am. J. Physiol. Endocrinol. Metab. 2020, 319, E827–E834. [Google Scholar] [CrossRef] [PubMed]
- Coussens, L.M.; Werb, Z. Inflammation and cancer. Nature 2002, 420, 860–867. [Google Scholar] [CrossRef]
- Chance, B.; Sies, H.; Boveris, A. Hydroperoxide metabolism in mammalian organs. Physiol. Rev. 1979, 59, 527–605. [Google Scholar] [CrossRef]
- Panov, A.V.; Dikalov, S.I. Cardiolipin, Perhydroxyl Radicals, and Lipid Peroxidation in Mitochondrial Dysfunctions and Aging. Oxid. Med. Cell. Longev. 2020, 2020, 1323028. [Google Scholar] [CrossRef] [PubMed]
- Dalle-Donne, I.; Giustarini, D.; Colombo, R.; Rossi, R.; Milzani, A. Protein carbonylation in human diseases. Trends Mol. Med. 2003, 9, 169–176. [Google Scholar] [CrossRef]
- Dupont, G.; Combettes, L.; Bird, G.S.; Putney, J.W. Calcium oscillations. Cold Spring Harb. Perspect. Biol. 2011, 3, a004226. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Bo, L.; Liu, Q.; Liu, W.; Chen, X.; Xu, D.; Jin, F. Activation of TRPV1-dependent calcium oscillation exacerbates seawater inhalation-induced acute lung injury. Mol. Med. Rep. 2016, 13, 1989–1998. [Google Scholar] [CrossRef] [Green Version]
- Seeley, E.J.; Rosenberg, P.; Matthay, M.A. Calcium flux and endothelial dysfunction during acute lung injury: A STIMulating target for therapy. J. Clin. Investig. 2013, 123, 1015–1018. [Google Scholar] [CrossRef] [Green Version]
- Gandhirajan, R.K.; Meng, S.; Chandramoorthy, H.C.; Mallilankaraman, K.; Mancarella, S.; Gao, H.; Razmpour, R.; Yang, X.F.; Houser, S.R.; Chen, J.; et al. Blockade of NOX2 and STIM1 signaling limits lipopolysaccharide-induced vascular inflammation. J. Clin. Investig. 2013, 123, 887–902. [Google Scholar] [CrossRef] [PubMed]
- Qiu, X.; Liang, X.; Li, H.; Sun, R. LPS-induced vein endothelial cell injury and acute lung injury have Btk and Orai 1 to regulate SOC-mediated calcium influx. Int. Immunopharmacol. 2021, 90, 107039. [Google Scholar] [CrossRef]
- Samapati, R.; Yang, Y.; Yin, J.; Stoerger, C.; Arenz, C.; Dietrich, A.; Gudermann, T.; Adam, D.; Wu, S.; Freichel, M.; et al. Lung endothelial Ca2+ and permeability response to platelet-activating factor is mediated by acid sphingomyelinase and transient receptor potential classical 6. Am. J. Respir. Crit. Care Med. 2012, 185, 160–170. [Google Scholar] [CrossRef] [PubMed]
- Tauseef, M.; Knezevic, N.; Chava, K.R.; Smith, M.; Sukriti, S.; Gianaris, N.; Obukhov, A.G.; Vogel, S.M.; Schraufnagel, D.E.; Dietrich, A.; et al. TLR4 activation of TRPC6-dependent calcium signaling mediates endotoxin-induced lung vascular permeability and inflammation. J. Exp. Med. 2012, 209, 1953–1968. [Google Scholar] [CrossRef]
- Granger, V.; Peyneau, M.; Chollet-Martin, S.; de Chaisemartin, L. Neutrophil Extracellular Traps in Autoimmunity and Allergy: Immune Complexes at Work. Front. Immunol. 2019, 10, 2824. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakai, J.; Li, J.; Subramanian, K.K.; Mondal, S.; Bajrami, B.; Hattori, H.; Jia, Y.; Dickinson, B.C.; Zhong, J.; Ye, K.; et al. Reactive oxygen species-induced actin glutathionylation controls actin dynamics in neutrophils. Immunity 2012, 37, 1037–1049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gwinn, M.R.; Vallyathan, V. Respiratory burst: Role in signal transduction in alveolar macrophages. J. Toxicol. Env. Health B Crit. Rev. 2006, 9, 27–39. [Google Scholar] [CrossRef] [PubMed]
- Iles, K.E.; Forman, H.J. Macrophage signaling and respiratory burst. Immunol. Res. 2002, 26, 95–105. [Google Scholar] [CrossRef]
- Millar, T.M.; Phan, V.; Tibbles, L.A. ROS generation in endothelial hypoxia and reoxygenation stimulates MAP kinase signaling and kinase-dependent neutrophil recruitment. Free Radic. Biol. Med. 2007, 42, 1165–1177. [Google Scholar] [CrossRef]
- Hattori, H.; Subramanian, K.K.; Sakai, J.; Luo, H.R. Reactive oxygen species as signaling molecules in neutrophil chemotaxis. Commun. Integr. Biol. 2010, 3, 278–281. [Google Scholar] [CrossRef] [PubMed]
- Miller, E.W.; Dickinson, B.C.; Chang, C.J. Aquaporin-3 mediates hydrogen peroxide uptake to regulate downstream intracellular signaling. Proc. Natl. Acad. Sci. USA 2010, 107, 15681–15686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okochi, Y.; Umemoto, E.; Okamura, Y. Hv1/VSOP regulates neutrophil directional migration and ERK activity by tuning ROS production. J. Leukoc. Biol. 2020, 107, 819–831. [Google Scholar] [CrossRef]
- Baruah, S.; Murthy, S.; Keck, K.; Galvan, I.; Prichard, A.; Allen, L.H.; Farrelly, M.; Klesney-Tait, J. TREM-1 regulates neutrophil chemotaxis by promoting NOX-dependent superoxide production. J. Leukoc. Biol. 2019, 105, 1195–1207. [Google Scholar] [CrossRef] [PubMed]
- Bastarache, J.A.; Sebag, S.C.; Clune, J.K.; Grove, B.S.; Lawson, W.E.; Janz, D.R.; Roberts, L.N.; Dworski, R.; Mackman, N.; Ware, L.B. Low levels of tissue factor lead to alveolar haemorrhage, potentiating murine acute lung injury and oxidative stress. Thorax 2012, 67, 1032–1039. [Google Scholar] [CrossRef] [Green Version]
- Ghio, A.J.; Richards, J.H.; Crissman, K.M.; Carter, J.D. Iron disequilibrium in the rat lung after instilled blood. Chest 2000, 118, 814–823. [Google Scholar] [CrossRef] [PubMed]
- Oh, J.Y.; Hamm, J.; Xu, X.; Genschmer, K.; Zhong, M.; Lebensburger, J.; Marques, M.B.; Kerby, J.D.; Pittet, J.F.; Gaggar, A.; et al. Absorbance and redox based approaches for measuring free heme and free hemoglobin in biological matrices. Redox Biol. 2016, 9, 167–177. [Google Scholar] [CrossRef] [Green Version]
- Shaver, C.M.; Upchurch, C.P.; Janz, D.R.; Grove, B.S.; Putz, N.D.; Wickersham, N.E.; Dikalov, S.I.; Ware, L.B.; Bastarache, J.A. Cell-free hemoglobin: A novel mediator of acute lung injury. Am. J. Physiol.-Lung Cell. Mol. Physiol. 2016, 310, L532–L541. [Google Scholar] [CrossRef] [Green Version]
- Gaggar, A.; Patel, R.P. There is blood in the water: Hemolysis, hemoglobin, and heme in acute lung injury. Am. J. Physiol.-Lung Cell. Mol. Physiol. 2016, 311, L714–L718. [Google Scholar] [CrossRef] [PubMed]
- Aggarwal, S.; Lazrak, A.; Ahmad, I.; Yu, Z.; Bryant, A.; Mobley, J.A.; Ford, D.A.; Matalon, S. Reactive species generated by heme impair alveolar epithelial sodium channel function in acute respiratory distress syndrome. Redox Biol. 2020, 36, 101592. [Google Scholar] [CrossRef]
- Byeon, J.C.; Lee, S.E.; Kim, T.H.; Ahn, J.B.; Kim, D.H.; Choi, J.S.; Park, J.S. Design of novel proliposome formulation for antioxidant peptide, glutathione with enhanced oral bioavailability and stability. Drug Deliv. 2019, 26, 216–225. [Google Scholar] [CrossRef] [Green Version]
- Tenorio, M.; Graciliano, N.G.; Moura, F.A.; Oliveira, A.; Goulart, M. N-Acetylcysteine (NAC): Impacts on Human Health. Antioxidants 2021, 10, 967. [Google Scholar] [CrossRef]
- Jugg, B.; Fairhall, S.; Smith, A.; Rutter, S.; Mann, T.; Perrott, R.; Jenner, J.; Salguero, J.; Shute, J.; Sciuto, A.M. N-acetyl-L-cysteine protects against inhaled sulfur mustard poisoning in the large swine. Clin. Toxicol. 2013, 51, 216–224. [Google Scholar] [CrossRef]
- Mokra, D.; Drgova, A.; Petras, M.; Mokry, J.; Antosova, M.; Calkovska, A. N-acetylcysteine alleviates the meconium-induced acute lung injury. Adv. Exp. Med. Biol. 2015, 832, 59–67. [Google Scholar] [PubMed]
- Bernard, G.R.; Wheeler, A.P.; Arons, M.M.; Morris, P.E.; Paz, H.L.; Russell, J.A.; Wright, P.E. A trial of antioxidants N-acetylcysteine and procysteine in ARDS. The Antioxidant in ARDS Study Group. Chest 1997, 112, 164–172. [Google Scholar] [CrossRef]
- De Backer, W.A.; Amsel, B.; Jorens, P.G.; Bossaert, L.; Hiemstra, P.S.; van Noort, P.; van Overveld, F.J. N-acetylcysteine pretreatment of cardiac surgery patients influences plasma neutrophil elastase and neutrophil influx in bronchoalveolar lavage fluid. Intensive Care Med. 1996, 22, 900–908. [Google Scholar] [CrossRef] [PubMed]
- Suter, P.M.; Domenighetti, G.; Schaller, M.D.; Laverriere, M.C.; Ritz, R.; Perret, C. N-acetylcysteine enhances recovery from acute lung injury in man. A randomized, double-blind, placebo-controlled clinical study. Chest 1994, 105, 190–194. [Google Scholar] [CrossRef] [Green Version]
- Bernard, G.R. N-acetylcysteine in experimental and clinical acute lung injury. Am. J. Med. 1991, 91, S54–S59. [Google Scholar] [CrossRef]
- Zhang, Y.; Ding, S.; Li, C.; Wang, Y.; Chen, Z.; Wang, Z. Effects of N-acetylcysteine treatment in acute respiratory distress syndrome: A meta-analysis. Exp. Ther. Med. 2017, 14, 2863–2868. [Google Scholar] [CrossRef]
- Alamdari, D.H.; Moghaddam, A.B.; Amini, S.; Keramati, M.R.; Zarmehri, A.M.; Alamdari, A.H.; Damsaz, M.; Banpour, H.; Yarahmadi, A.; Koliakos, G. Application of methylene blue -vitamin C -N-acetyl cysteine for treatment of critically ill COVID-19 patients, report of a phase-I clinical trial. Eur. J. Pharmacol. 2020, 885, 173494. [Google Scholar] [CrossRef] [PubMed]
- Carothers, C.; Birrer, K.; Vo, M. Acetylcysteine for the Treatment of Suspected Remdesivir-Associated Acute Liver Failure in COVID-19: A Case Series. Pharmacotherapy 2020, 40, 1166–1171. [Google Scholar] [CrossRef]
- Ibrahim, H.; Perl, A.; Smith, D.; Lewis, T.; Kon, Z.; Goldenberg, R.; Yarta, K.; Staniloae, C.; Williams, M. Therapeutic blockade of inflammation in severe COVID-19 infection with intravenous N-acetylcysteine. Clin. Immunol. 2020, 219, 108544. [Google Scholar] [CrossRef]
- Ji, L.; Liu, R.; Zhang, X.D.; Chen, H.L.; Bai, H.; Wang, X.; Zhao, H.L.; Liang, X.; Hai, C.X. N-acetylcysteine attenuates phosgene-induced acute lung injury via up-regulation of Nrf2 expression. Inhal. Toxicol. 2010, 22, 535–542. [Google Scholar] [CrossRef]
- Pan, X.; Wu, X.; Yan, D.; Peng, C.; Rao, C.; Yan, H. Acrylamide-induced oxidative stress and inflammatory response are alleviated by N-acetylcysteine in PC12 cells: Involvement of the crosstalk between Nrf2 and NF-kappaB pathways regulated by MAPKs. Toxicol. Lett. 2018, 288, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Ren, J.; Su, D.; Li, L.; Cai, H.; Zhang, M.; Zhai, J.; Li, M.; Wu, X.; Hu, K. Anti-inflammatory effects of Aureusidin in LPS-stimulated RAW264.7 macrophages via suppressing NF-kappaB and activating ROS- and MAPKs-dependent Nrf2/HO-1 signaling pathways. Toxicol. Appl. Pharm. 2020, 387, 114846. [Google Scholar] [CrossRef]
- Song, Q.; Lin, L.; Chen, L.; Cheng, L.; Zhong, W. Co-administration of N-acetylcysteine and dexmedetomidine plays a synergistic effect on protection of LPS-induced acute lung injury via correcting Th1/Th2/Th17 cytokines imbalance. Clin. Exp. Pharm. Physiol. 2020, 47, 294–301. [Google Scholar] [CrossRef] [PubMed]
- Gelfand, C.A.; Sakurai, R.; Wang, Y.; Liu, Y.; Segal, R.; Rehan, V.K. Inhaled vitamin A is more effective than intramuscular dosing in mitigating hyperoxia-induced lung injury in a neonatal rat model of bronchopulmonary dysplasia. Am. J. Physiol.-Lung Cell. Mol. Physiol. 2020, 319, L576–L584. [Google Scholar] [CrossRef]
- Al-Harbi, N.O.; Imam, F.; Nadeem, A.; Al-Harbi, M.M.; Korashy, H.M.; Sayed-Ahmed, M.M.; Hafez, M.M.; Al-Shabanah, O.A.; Nagi, M.N.; Bahashwan, S. Riboflavin attenuates lipopolysaccharide-induced lung injury in rats. Toxicol. Mech. Methods 2015, 25, 417–423. [Google Scholar] [CrossRef] [PubMed]
- Cerrada, A.; Haller, T.; Cruz, A.; Perez-Gil, J. Pneumocytes Assemble Lung Surfactant as Highly Packed/Dehydrated States with Optimal Surface Activity. Biophys. J. 2015, 109, 2295–2306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kullberg, A.; Ekholm, O.O.; Slotte, J.P. Miscibility of Sphingomyelins and Phosphatidylcholines in Unsaturated Phosphatidylcholine Bilayers. Biophys. J. 2015, 109, 1907–1916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wemhoner, A.; Jennings, P.; Haller, T.; Rudiger, M.; Simbruner, G. Effect of exogenous surfactants on viability and DNA synthesis in A549, immortalized mouse type II and isolated rat alveolar type II cells. BMC Pulm. Med. 2011, 11, 11. [Google Scholar] [CrossRef] [Green Version]
- Morita, N.; Shimoda, K.; Traber, M.G.; Westphal, M.; Enkhbaatar, P.; Murakami, K.; Leonard, S.W.; Traber, L.D.; Traber, D.L. Vitamin E attenuates acute lung injury in sheep with burn and smoke inhalation injury. Redox Rep. 2006, 11, 61–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xantus, G.; Anna, G.V.; Johnson, C.A.; Sanghera, P.; Zavori, L.; Kanizsai, P.L. The role of vitamin E acetate (VEA) and its derivatives in the vaping associated lung injury: Systematic review of evidence. Crit. Rev. Toxicol. 2021, 51, 15–23. [Google Scholar] [CrossRef]
- Yamaoka, S.; Kim, H.S.; Ogihara, T.; Oue, S.; Takitani, K.; Yoshida, Y.; Tamai, H. Severe Vitamin E deficiency exacerbates acute hyperoxic lung injury associated with increased oxidative stress and inflammation. Free Radic. Res 2008, 42, 602–612. [Google Scholar] [CrossRef] [PubMed]
- Mudway, I.S.; Behndig, A.F.; Helleday, R.; Pourazar, J.; Frew, A.J.; Kelly, F.J.; Blomberg, A. Vitamin supplementation does not protect against symptoms in ozone-responsive subjects. Free Radic. Biol. Med. 2006, 40, 1702–1712. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.J.; Jiang, J.X.; Ren, Q.Q.; Jia, Y.L.; Shen, J.; Shen, H.J.; Lin, X.X.; Lu, H.; Xie, Q.M. Ambroxol inhalation ameliorates LPS-induced airway inflammation and mucus secretion through the extracellular signal-regulated kinase 1/2 signaling pathway. Eur. J. Pharmacol. 2016, 775, 138–148. [Google Scholar] [CrossRef] [PubMed]
- Piotrowski, W.J.; Pietras, T.; Kurmanowska, Z.; Nowak, D.; Marczak, J.; Marks-Konczalik, J.; Mazerant, P. Effect of paraquat intoxication and ambroxol treatment on hydrogen peroxide production and lipid peroxidation in selected organs of rat. J. Appl. Toxicol. 1996, 16, 501–507. [Google Scholar] [CrossRef]
- Ge, L.T.; Liu, Y.N.; Lin, X.X.; Shen, H.J.; Jia, Y.L.; Dong, X.W.; Sun, Y.; Xie, Q.M. Inhalation of ambroxol inhibits cigarette smoke-induced acute lung injury in a mouse model by inhibiting the Erk pathway. Int. Immunopharmacol. 2016, 33, 90–98. [Google Scholar] [CrossRef] [Green Version]
- Su, X.; Wang, L.; Song, Y.; Bai, C. Inhibition of inflammatory responses by ambroxol, a mucolytic agent, in a murine model of acute lung injury induced by lipopolysaccharide. Intensive Care Med. 2004, 30, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Boo, Y.C. Human Skin Lightening Efficacy of Resveratrol and Its Analogs: From in Vitro Studies to Cosmetic Applications. Antioxidants 2019, 8, 332. [Google Scholar] [CrossRef] [Green Version]
- Jiang, L.; Zhang, L.; Kang, K.; Fei, D.; Gong, R.; Cao, Y.; Pan, S.; Zhao, M.; Zhao, M. Resveratrol ameliorates LPS-induced acute lung injury via NLRP3 inflammasome modulation. Biomed. Pharmacother. 2016, 84, 130–138. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Zhang, Z.; Zhuo, Y.; Cui, L.; Li, C.; Li, D.; Zhang, S.; Cui, N.; Wang, X.; Gao, H. Resveratrol alleviates sepsis-induced acute lung injury by suppressing inflammation and apoptosis of alveolar macrophage cells. Am. J. Transl. Res. 2018, 10, 1961–1975. [Google Scholar]
- Cheng, K.; Yang, A.; Hu, X.; Zhu, D.; Liu, K. Curcumin Attenuates Pulmonary Inflammation in Lipopolysaccharide Induced Acute Lung Injury in Neonatal Rat Model by Activating Peroxisome Proliferator-Activated Receptor gamma (PPARgamma) Pathway. Med. Sci. Monit. 2018, 24, 1178–1184. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, Y.; Cai, N.; Xu, T.; He, F. Anti-inflammatory effects of curcumin in acute lung injury: In vivo and in vitro experimental model studies. Int. Immunopharmacol. 2021, 96, 107600. [Google Scholar] [CrossRef]
- Shaikh, S.B.; Najar, M.A.; Prasad, T.; Bhandary, Y.P. Comparative protein profiling reveals the inhibitory role of curcumin on IL-17A mediated minichromosome maintenance (MCM) proteins as novel putative markers for acute lung injury in vivo. Biomed. Pharmacother. 2021, 141, 111715. [Google Scholar] [CrossRef]
- Shaikh, S.B.; Bhat, S.G.; Bhandary, Y.P. Curcumin attenuates IL-17A mediated pulmonary SMAD dependent and non-dependent mechanism during acute lung injury in vivo. Mol. Biol. Rep. 2020, 47, 5643–5649. [Google Scholar] [CrossRef]
- Gouda, M.M.; Prabhu, A.; Bhandary, Y.P. Curcumin alleviates IL-17A-mediated p53-PAI-1 expression in bleomycin-induced alveolar basal epithelial cells. J. Cell. Biochem. 2018, 119, 2222–2230. [Google Scholar] [CrossRef] [PubMed]
- Kim, G.; Piao, C.; Oh, J.; Lee, M. Combined delivery of curcumin and the heme oxygenase-1 gene using cholesterol-conjugated polyamidoamine for anti-inflammatory therapy in acute lung injury. Phytomedicine 2019, 56, 165–174. [Google Scholar] [CrossRef] [PubMed]
- Kim, G.; Lee, Y.; Ha, J.; Han, S.; Lee, M. Engineering exosomes for pulmonary delivery of peptides and drugs to inflammatory lung cells by inhalation. J. Control. Release 2021, 330, 684–695. [Google Scholar] [CrossRef]
- Ma, C.; Zhu, L.; Wang, J.; He, H.; Chang, X.; Gao, J.; Shumin, W.; Yan, T. Anti-inflammatory effects of water extract of Taraxacum mongolicum hand.-Mazz on lipopolysaccharide-induced inflammation in acute lung injury by suppressing PI3K/Akt/mTOR signaling pathway. J. Ethnopharmacol. 2015, 168, 349–355. [Google Scholar] [CrossRef]
- Lee, J.W.; Seo, K.H.; Ryu, H.W.; Yuk, H.J.; Park, H.A.; Lim, Y.; Ahn, K.S.; Oh, S.R. Anti-inflammatory effect of stem bark of Paulownia tomentosa Steud. in lipopolysaccharide (LPS)-stimulated RAW264.7 macrophages and LPS-induced murine model of acute lung injury. J. Ethnopharmacol. 2018, 210, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Li, C.; Shen, F.; Wang, M.; Jia, N.; Wang, C. Naringenin ameliorates LPS-induced acute lung injury through its anti-oxidative and anti-inflammatory activity and by inhibition of the PI3K/AKT pathway. Exp. Ther. Med. 2017, 14, 2228–2234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mei, H.; Tao, Y.; Zhang, T.; Qi, F. Emodin alleviates LPS-induced inflammatory response in lung injury rat by affecting the function of granulocytes. J. Inflamm. 2020, 17, 26. [Google Scholar] [CrossRef]
- Lv, H.; Liu, Q.; Wen, Z.; Feng, H.; Deng, X.; Ci, X. Xanthohumol ameliorates lipopolysaccharide (LPS)-induced acute lung injury via induction of AMPK/GSK3beta-Nrf2 signal axis. Redox Biol. 2017, 12, 311–324. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Ji, M.; Chen, L.; Wu, X.; Wang, L. Breviscapine reduces acute lung injury induced by left heart ischemic reperfusion in rats by inhibiting the expression of ICAM-1 and IL-18. Exp. Ther. Med. 2013, 6, 1322–1326. [Google Scholar] [CrossRef]
- Song, Y.; Leonard, S.W.; Traber, M.G.; Ho, E. Zinc deficiency affects DNA damage, oxidative stress, antioxidant defenses, and DNA repair in rats. J. Nutr. 2009, 139, 1626–1631. [Google Scholar] [CrossRef] [Green Version]
- Gunther, V.; Lindert, U.; Schaffner, W. The taste of heavy metals: Gene regulation by MTF-1. Biochim. Biophys. Acta 2012, 1823, 1416–1425. [Google Scholar] [CrossRef] [Green Version]
- Efeovbokhan, N.; Bhattacharya, S.K.; Ahokas, R.A.; Sun, Y.; Guntaka, R.V.; Gerling, I.C.; Weber, K.T. Zinc and the prooxidant heart failure phenotype. J. Cardiovasc. Pharm. 2014, 64, 393–400. [Google Scholar] [CrossRef] [Green Version]
- Pal, D.T.; Gowda, N.K.; Sampath, K.T. Effect of copper- and zinc-methionine supplementation on bioavailability, mineral status and tissue concentrations of copper and zinc in ewes. J. Trace Elem. Med. Biol. 2010, 24, 89–94. [Google Scholar] [CrossRef] [PubMed]
- Wan, C.; Zhang, M.; Fang, Q.; Xiong, L.; Zhao, X.; Hasunuma, T.; Bai, F.; Kondo, A. The impact of zinc sulfate addition on the dynamic metabolic profiling of Saccharomyces cerevisiae subjected to long term acetic acid stress treatment and identification of key metabolites involved in the antioxidant effect of zinc. Metallomics 2015, 7, 322–332. [Google Scholar] [CrossRef] [PubMed]
- St Croix, C.M.; Leelavaninchkul, K.; Watkins, S.C.; Kagan, V.E.; Pitt, B.R. Nitric oxide and zinc homeostasis in acute lung injury. Proc. Am. Thorac. Soc. 2005, 2, 236–242. [Google Scholar] [CrossRef]
- Taylor, C.G.; Towner, R.A.; Janzen, E.G.; Bray, T.M. MRI detection of hyperoxia-induced lung edema in Zn-deficient rats. Free Radic. Biol. Med. 1990, 9, 229–233. [Google Scholar] [CrossRef]
- Taylor, C.G.; McCutchon, T.L.; Boermans, H.J.; DiSilvestro, R.A.; Bray, T.M. Comparison of Zn and vitamin E for protection against hyperoxia-induced lung damage. Free Radic. Biol. Med. 1997, 22, 543–550. [Google Scholar] [CrossRef]
- Anttinen, H.; Oikarinen, A.; Puistola, U.; Paakko, P.; Ryhanen, L. Prevention by zinc of rat lung collagen accumulation in carbon tetrachloride injury. Am. Rev. Respir. Dis. 1985, 132, 536–540. [Google Scholar] [PubMed]
- Wessels, I.; Pupke, J.T.; von Trotha, K.T.; Gombert, A.; Himmelsbach, A.; Fischer, H.J.; Jacobs, M.J.; Rink, L.; Grommes, J. Zinc supplementation ameliorates lung injury by reducing neutrophil recruitment and activity. Thorax 2020, 75, 253–261. [Google Scholar] [CrossRef]
- Liu, J.; Yang, Y.; Zeng, X.; Bo, L.; Jiang, S.; Du, X.; Xie, Y.; Jiang, R.; Zhao, J.; Song, W. Investigation of selenium pretreatment in the attenuation of lung injury in rats induced by fine particulate matters. Env. Sci. Pollut. Res. Int. 2017, 24, 4008–4017. [Google Scholar] [CrossRef]
- Jia, W.; Ding, W.; Chen, X.; Xu, Z.; Tang, Y.; Wang, M.; Zheng, B.; Zhang, Y.; Wei, T.; Zhu, Z. Selenium-Containing Compound Ameliorates Lipopolysaccharide-Induced Acute Lung Injury via Regulating the MAPK/AP-1 Pathway. Inflammation 2021, 44, 2518–2530. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.S.; Suh, G.J.; Kwon, W.Y.; Kwak, Y.H.; Lee, K.; Lee, H.J.; Jeong, K.Y.; Lee, M.W. Antioxidant effects of selenium on lung injury in paraquat intoxicated rats. Clin. Toxicol. 2012, 50, 749–753. [Google Scholar] [CrossRef]
- Kwon, W.Y.; Suh, G.J.; Kim, K.S.; Jung, Y.S.; Kim, S.H.; Kim, J.S.; You, K.M. Niacin and Selenium Attenuate Sepsis-Induced Lung Injury by Up-Regulating Nuclear Factor Erythroid 2-Related Factor 2 Signaling. Crit. Care Med. 2016, 44, e370–e382. [Google Scholar] [CrossRef]
- HARMAN, D. Aging: A theory based on free radical and radiation chemistry. J. Gerontol. 1956, 11, 298–300. [Google Scholar] [CrossRef] [Green Version]
- McCord, J.M.; Fridovich, I. The utility of superoxide dismutase in studying free radical reactions. I. Radicals generated by the interaction of sulfite, dimethyl sulfoxide, and oxygen. J. Biol. Chem. 1969, 244, 6056–6063. [Google Scholar] [CrossRef]
- Lei, J.; Wei, Y.; Song, P.; Li, Y.; Zhang, T.; Feng, Q.; Xu, G. Cordycepin inhibits LPS-induced acute lung injury by inhibiting inflammation and oxidative stress. Eur. J. Pharmacol. 2018, 818, 110–114. [Google Scholar] [CrossRef] [PubMed]
- Cipak, G.A. Free Radical Research in Cancer. Antioxidants 2020, 9, 157. [Google Scholar] [CrossRef] [Green Version]
- Xu, T.; Ding, W.; Ji, X.; Ao, X.; Liu, Y.; Yu, W.; Wang, J. Oxidative Stress in Cell Death and Cardiovascular Diseases. Oxid. Med. Cell. Longev. 2019, 2019, 9030563. [Google Scholar] [CrossRef] [Green Version]
- Scicchitano, P.; Cortese, F.; Gesualdo, M.; De Palo, M.; Massari, F.; Giordano, P.; Ciccone, M.M. The role of endothelial dysfunction and oxidative stress in cerebrovascular diseases. Free Radic. Res. 2019, 53, 579–595. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhou, Y.; Gao, Q.; Ping, D.; Wang, Y.; Wu, W.; Lin, X.; Fang, Y.; Zhang, J.; Shao, A. The Role of Exosomal microRNAs and Oxidative Stress in Neurodegenerative Diseases. Oxid. Med. Cell. Longev. 2020, 2020, 3232869. [Google Scholar] [CrossRef]
- Da, F.L.; Nunes-Souza, V.; Goulart, M.; Rabelo, L.A. Oxidative Stress in Rheumatoid Arthritis: What the Future Might Hold regarding Novel Biomarkers and Add-On Therapies. Oxid. Med. Cell Longev. 2019, 2019, 7536805. [Google Scholar] [CrossRef] [Green Version]
- Luc, K.; Schramm-Luc, A.; Guzik, T.J.; Mikolajczyk, T.P. Oxidative stress and inflammatory markers in prediabetes and diabetes. J. Physiol. Pharmacol. 2019, 70, 809–824. [Google Scholar] [CrossRef]
- Rice, T.W.; Wheeler, A.P.; Thompson, B.T.; DeBoisblanc, B.P.; Steingrub, J.; Rock, P. Enteral omega-3 fatty acid, gamma-linolenic acid, and antioxidant supplementation in acute lung injury. JAMA 2011, 306, 1574–1581. [Google Scholar] [CrossRef] [Green Version]
- Heart Protection Study Collaborative Group. MRC/BHF Heart Protection Study of antioxidant vitamin supplementation in 20,536 high-risk individuals: A randomised placebo-controlled trial. Lancet 2002, 360, 23–33. [Google Scholar] [CrossRef]
- Bjelakovic, G.; Nikolova, D.; Gluud, L.L.; Simonetti, R.G.; Gluud, C. Antioxidant supplements for prevention of mortality in healthy participants and patients with various diseases. Cochrane Database Syst. Rev. 2012, 2012, D7176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, E.R.; Pastor-Barriuso, R.; Dalal, D.; Riemersma, R.A.; Appel, L.J.; Guallar, E. Meta-analysis: High-dosage vitamin E supplementation may increase all-cause mortality. Ann. Intern. Med. 2005, 142, 37–46. [Google Scholar] [CrossRef] [Green Version]
- Bendich, A.; Machlin, L.J. Safety of oral intake of vitamin E. Am. J. Clin. Nutr. 1988, 48, 612–619. [Google Scholar] [CrossRef]
- Roberts, H.J. Perspective on vitamin E as therapy. JAMA 1981, 246, 129–131. [Google Scholar] [CrossRef] [PubMed]
- Abudu, N.; Miller, J.J.; Attaelmannan, M.; Levinson, S.S. Vitamins in human arteriosclerosis with emphasis on vitamin C and vitamin E. Clin. Chim. Acta 2004, 339, 11–25. [Google Scholar] [CrossRef]
- Bowry, V.W.; Mohr, D.; Cleary, J.; Stocker, R. Prevention of tocopherol-mediated peroxidation in ubiquinol-10-free human low density lipoprotein. J. Biol. Chem. 1995, 270, 5756–5763. [Google Scholar] [CrossRef] [Green Version]
- Huang, H.Y.; Appel, L.J. Supplementation of diets with alpha-tocopherol reduces serum concentrations of gamma- and delta-tocopherol in humans. J. Nutr. 2003, 133, 3137–3140. [Google Scholar] [CrossRef] [Green Version]
- Kim, T.J.; Byun, J.S.; Kwon, H.S.; Kim, D.Y. Cellular toxicity driven by high-dose vitamin C on normal and cancer stem cells. Biochem. Biophys. Res. Commun. 2018, 497, 347–353. [Google Scholar] [CrossRef]
- Dahlgren, C.; Karlsson, A. Respiratory burst in human neutrophils. J. Immunol. Methods 1999, 232, 3–14. [Google Scholar] [CrossRef]
- Wang, T.; Zhang, H.; Han, Y.; Liu, H.; Ren, F.; Zeng, J.; Sun, Q.; Li, Z.; Gao, M. Light-Enhanced O2-Evolving Nanoparticles Boost Photodynamic Therapy To Elicit Antitumor Immunity. ACS Appl. Mater. Interfaces 2019, 11, 16367–16379. [Google Scholar] [CrossRef] [PubMed]
- Calabrese, E.J.; Baldwin, L.A. Inorganics and hormesis. Crit. Rev. Toxicol. 2003, 33, 215–304. [Google Scholar] [CrossRef] [PubMed]
- Calabrese, E.J. Hormesis: Path and Progression to Significance. Int. J. Mol. Sci. 2018, 19, 2871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Z.; Chi, J.; Feng, Z. Observation of curative effect of nasal continuous positive airway pressure combined with high-dose ambroxol for neonatal respiratory failure. Pak. J. Pharm. Sci. 2018, 31, 1663–1666. [Google Scholar] [PubMed]
- Xiang, J.; Wang, P. Efficacy of pulmonary surfactant combined with high-dose ambroxol hydrochloride in the treatment of neonatal respiratory distress syndrome. Exp. Ther. Med. 2019, 18, 654–658. [Google Scholar] [CrossRef] [PubMed] [Green Version]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Y.; Zhou, S.; Xiang, D.; Ju, L.; Shen, D.; Wang, X.; Wang, Y. Friend or Foe? The Roles of Antioxidants in Acute Lung Injury. Antioxidants 2021, 10, 1956. https://doi.org/10.3390/antiox10121956
Liu Y, Zhou S, Xiang D, Ju L, Shen D, Wang X, Wang Y. Friend or Foe? The Roles of Antioxidants in Acute Lung Injury. Antioxidants. 2021; 10(12):1956. https://doi.org/10.3390/antiox10121956
Chicago/Turabian StyleLiu, Yang, Shujun Zhou, Du Xiang, Lingao Ju, Dexin Shen, Xinghuan Wang, and Yanfeng Wang. 2021. "Friend or Foe? The Roles of Antioxidants in Acute Lung Injury" Antioxidants 10, no. 12: 1956. https://doi.org/10.3390/antiox10121956