
Exercise Training Induces a Shift in Extracellular Redox Status with Alterations in the Pulmonary and Systemic Redox Landscape in Asthma

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects
2.2. Exercise Intervention and Testing
2.3. Sample Collection and Processing
2.4. Outcome Measures
2.5. Data Analysis
3. Results
3.1. Demographics and Physiological Data
3.2. Asthma-Related Clinical Outcomes
3.3. The Integrated Adaptive Redox Response to Exercise in Patients with Asthma
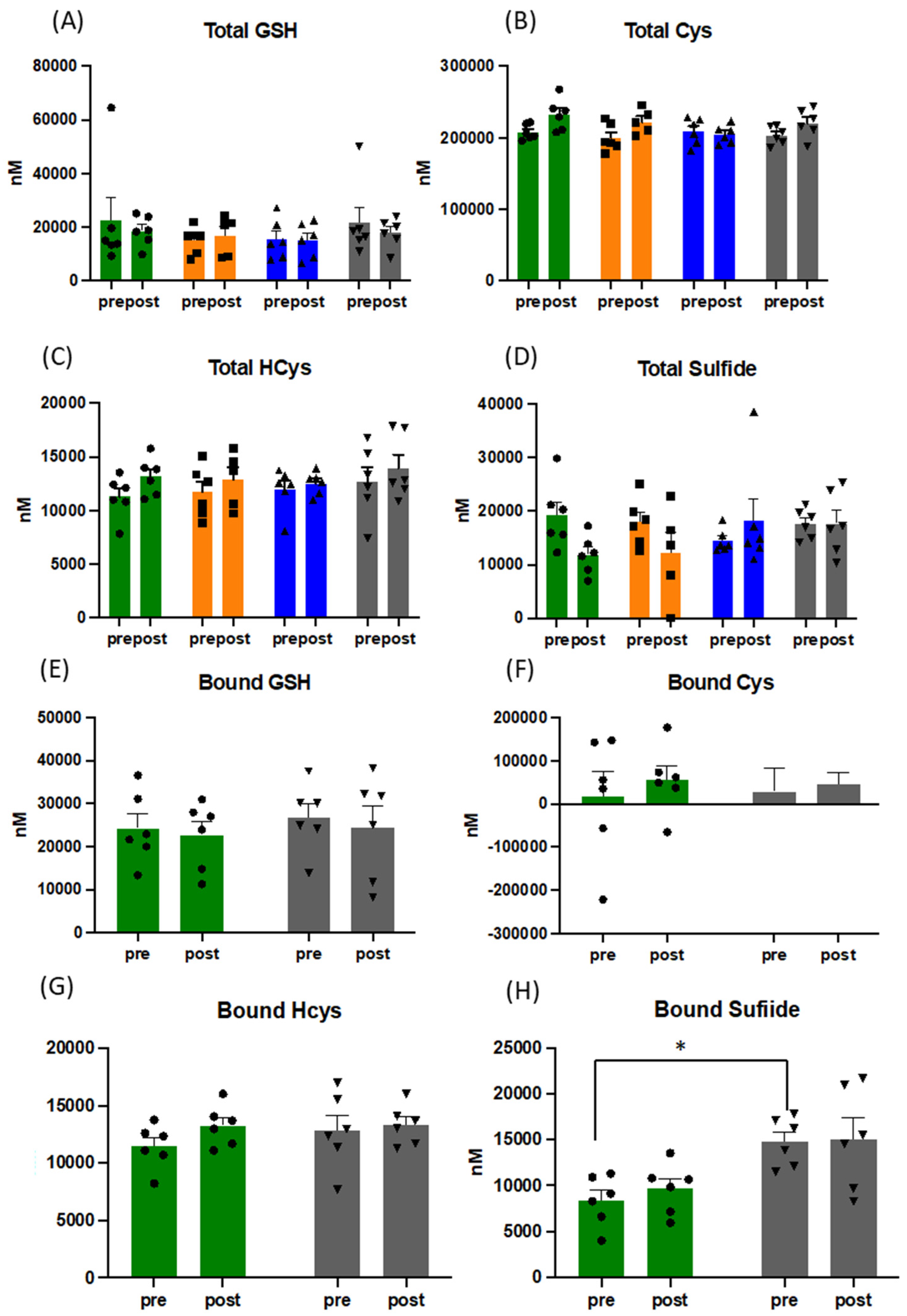
3.4. Acute and Long-Term Changes of the Thiol Metabolome in Response to Exercise
3.5. Effects of Exercise on Nitric Oxide Metabolites, Lipid Oxidation Products and Reducing Capacity
3.6. Redox Status Is Associated with Improved Physical Fitness and Reduced Inflammation
4. Discussion
Study Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Neufer, P.D.; Bamman, M.M.; Muoio, D.; Bouchard, C.; Cooper, D.M.; Goodpaster, B.H.; Booth, F.W.; Kohrt, W.M.; Gerszten, R.E.; Mattson, M.P.; et al. Understanding the Cellular and Molecular Mechanisms of Physical Activity-Induced Health Benefits. Cell Metab. 2015, 22, 4–11. [Google Scholar] [CrossRef] [Green Version]
- West, M.A.; Astin, R.; Moyses, H.E.; Cave, J.; White, D.; Levett, D.Z.H.; Bates, A.; Brown, G.; Grocott, M.P.W.; Jack, S. Exercise prehabilitation may lead to augmented tumor regression following neoadjuvant chemoradiotherapy in locally advanced rectal cancer. Acta Oncol. 2019, 58, 588–595. [Google Scholar] [CrossRef] [Green Version]
- Contrepois, K.; Wu, S.; Moneghetti, K.J.; Hornburg, D.; Ahadi, S.; Tsai, M.-S.; Metwally, A.A.; Wei, E.; Lee-McMullen, B.; Quijada, J.V.; et al. Molecular Choreography of Acute Exercise. Cell 2020, 181, 1112–1130.e16. [Google Scholar] [CrossRef]
- Powers, S.K.; Jackson, M.J. Exercise-Induced Oxidative Stress: Cellular Mechanisms and Impact on Muscle Force Production. Physiol. Rev. 2008, 88, 1243–1276. [Google Scholar] [CrossRef] [Green Version]
- Margaritelis, N.; Paschalis, V.; Theodorou, A.; Kyparos, A.; Nikolaidis, M. Redox basis of exercise physiology. Redox Biol. 2020, 35, 101499. [Google Scholar] [CrossRef] [PubMed]
- Ristow, M.; Zarse, K.; Oberbach, A.; Klöting, N.; Birringer, M.; Kiehntopf, M.; Stumvoll, M.; Kahn, C.R.; Blüher, M. Antioxidants prevent health-promoting effects of physical exercise in humans. Proc. Natl. Acad. Sci. USA 2009, 106, 8665–8670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, L.; Diaz, P.T.; Best, T.M.; Stimpfl, J.N.; He, F.; Zuo, L. Molecular characterization of redox mechanisms in allergic asthma. Ann. Allergy Asthma Immunol. 2014, 113, 137–142. [Google Scholar] [CrossRef] [PubMed]
- Page, L.K.; Staples, K.J.; Spalluto, C.M.; Watson, A.; Wilkinson, T. Influence of Hypoxia on the Epithelial-Pathogen Interactions in the Lung: Implications for Respiratory Disease. Front. Immunol. 2021, 12, 939. [Google Scholar] [CrossRef] [PubMed]
- Comhair, S.A.; Erzurum, S.C. Redox Control of Asthma: Molecular Mechanisms and Therapeutic Opportunities. Antioxid. Redox Signal. 2010, 12, 93–124. [Google Scholar] [CrossRef] [Green Version]
- Sanders, S.P.; Zweier, J.L.; Harrison, S.J.; Trush, A.M.; Rembish, S.J.; Liu, M.C. Spontaneous oxygen radical production at sites of antigen challenge in allergic subjects. Am. J. Respir. Crit. Care Med. 1995, 151, 1725–1733. [Google Scholar] [CrossRef]
- Nadeem, A.; Raj, H.G.; Chhabra, S.K. Increased Oxidative Stress in Acute Exacerbations of Asthma. J. Asthma 2005, 42, 45–50. [Google Scholar] [CrossRef]
- Fitzpatrick, A.M.; Stephenson, S.T.; Hadley, G.R.; Burwell, L.; Penugonda, M.; Simon, D.M.; Hansen, J.; Jones, D.P.; Brown, L.A.S. Thiol redox disturbances in children with severe asthma are associated with posttranslational modification of the transcription factor nuclear factor (erythroid-derived 2)–like 2. J. Allergy Clin. Immunol. 2011, 127, 1604–1611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stephenson, S.T.; Brown, L.A.S.; Helms, M.N.; Qu, H.; Brown, S.D.; Brown, M.R.; Fitzpatrick, A.M. Cysteine oxidation impairs systemic glucocorticoid responsiveness in children with difficult-to-treat asthma. J. Allergy Clin. Immunol. 2015, 136, 454–461.e9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janssen-Heininger, Y.; Reynaert, N.L.; van der Vliet, A.; Anathy, V. Endoplasmic reticulum stress and glutathione therapeutics in chronic lung diseases. Redox Biol. 2020, 33, 101516. [Google Scholar] [CrossRef]
- Chia, S.B.; Elko, E.A.; Aboushousha, R.; Manuel, A.M.; van de Wetering, C.; Druso, J.E.; van der Velden, J.; Seward, D.J.; Anathy, V.; Irvin, C.G.; et al. Dysregulation of the glutaredoxin/S-glutathionylation redox axis in lung diseases. Am. J. Physiol. 2020, 318, C304–C327. [Google Scholar] [CrossRef]
- Suzuki, Y.; Saito, J.; Munakata, M.; Shibata, Y. Hydrogen sulfide as a novel biomarker of asthma and chronic obstructive pulmonary disease. Allergol. Int. 2021, 70, 181–189. [Google Scholar] [CrossRef] [PubMed]
- Webb, R.; Hughes, M.G.; Thomas, A.W.; Morris, K. The Ability of Exercise-Associated Oxidative Stress to Trigger Redox-Sensitive Signalling Responses. Antioxidants 2017, 6, 63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cumpstey, A.F.; Minnion, M.; Fernandez, B.O.; Mikus-Lelinska, M.; Mitchell, K.; Martin, D.S.; Grocott, M.P.; Feelisch, M. Pushing arterial-venous plasma biomarkers to new heights: A model for personalised redox metabolomics? Redox Biol. 2019, 21, 101113. [Google Scholar] [CrossRef]
- Done, A.J.; Traustadóttir, T. Nrf2 mediates redox adaptations to exercise. Redox Biol. 2016, 10, 191–199. [Google Scholar] [CrossRef] [Green Version]
- Pakhale, S.; Luks, V.; Burkett, A.; Turner, L. Effect of physical training on airway inflammation in bronchial asthma: A systematic review. BMC Pulm. Med. 2013, 13, 38. [Google Scholar] [CrossRef] [Green Version]
- Hansen, E.S.H.; Pitzner-Fabricius, A.; Toennesen, L.L.; Rasmusen, H.K.; Hostrup, M.; Hellsten, Y.; Backer, V.; Henriksen, M. Effect of aerobic exercise training on asthma in adults: A systematic review and meta-analysis. Eur. Respir. J. 2020, 56, 2000146. [Google Scholar] [CrossRef]
- Cortese-Krott, M.M.; Koning, A.; Kuhnle, G.G.C.; Nagy, P.; Bianco, C.L.; Pasch, A.; Wink, D.A.; Fukuto, J.M.; Jackson, A.A.; Van Goor, H.; et al. The Reactive Species Interactome: Evolutionary Emergence, Biological Significance, and Opportunities for Redox Metabolomics and Personalized Medicine. Antioxid. Redox Signal. 2017, 27, 684–712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santolini, J.; Wootton, A.S.; Jackson, A.A.; Feelisch, M. The Redox architecture of physiological function. Curr. Opin. Physiol. 2019, 9, 34–47. [Google Scholar] [CrossRef]
- Codella, R.; Chirico, A.; Lucidi, F.; Ferrulli, A.; La Torre, A.; Luzi, L. The immune-modulatory effects of exercise should be favorably harnessed against COVID-19. J. Endocrinol. Investig. 2021, 44, 1119–1122. [Google Scholar] [CrossRef] [PubMed]
- Marrocco, I.; Altieri, F.; Peluso, I. Measurement and Clinical Significance of Biomarkers of Oxidative Stress in Humans. Oxidative Med. Cell. Longev. 2017, 2017, 6501046. [Google Scholar] [CrossRef] [PubMed]
- Cumpstey, A.F.; Clark, A.D.; Santolini, J.; Jackson, A.A.; Feelisch, M. COVID-19: A Redox Disease—What a Stress Pandemic Can Teach Us About Resilience and What We May Learn from the Reactive Species Interactome About Its Treatment. Antioxid. Redox Signal. 2021, 35, 1226–1268. [Google Scholar] [CrossRef]
- Juniper, E.F.; Bousquet, J.; Abetz, L.; Bateman, E.D. Identifying ‘well-controlled’ and ‘not well-controlled’ asthma using the Asthma Control Questionnaire. Respir. Med. 2006, 100, 616–621. [Google Scholar] [CrossRef] [Green Version]
- Levett, D.; Jack, S.; Swart, M.; Carlisle, J.; Wilson, J.; Snowden, C.; Riley, M.; Danjoux, G.; Ward, S.; Older, P.; et al. Perioperative cardiopulmonary exercise testing (CPET): Consensus clinical guidelines on indications, organization, conduct, and physiological interpretation. Br. J. Anaesth. 2018, 120, 484–500. [Google Scholar] [CrossRef] [Green Version]
- West, M.; Loughney, L.; Lythgoe, D.; Barben, C.; Sripadam, R.; Kemp, G.; Grocott, M.; Jack, S. Effect of prehabilitation on objectively measured physical fitness after neoadjuvant treatment in preoperative rectal cancer patients: A blinded interventional pilot study. Br. J. Anaesth. 2015, 114, 244–251. [Google Scholar] [CrossRef] [Green Version]
- Horváth, I.; Hunt, J.; Barnes, P.J. Exhaled breath condensate: Methodological recommendations and unresolved questions. Eur. Respir. J. 2005, 26, 523–548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rassaf, T.; Bryan, N.S.; Kelm, M.; Feelisch, M. Concomitant presence of N-nitroso and S-nitroso proteins in human plasma. Free Radic. Biol. Med. 2002, 33, 1590–1596. [Google Scholar] [CrossRef]
- Feelisch, M.; Rassaf, T.; Mnaimneh, S.; Singh, N.; Bryan, N.S.; Jourd’Heuil, D.; Kelm, M. Concomitant S-, N-, and heme-nitros(yl)ation in biological tissues and fluids: Implications for the fate of NO in vivo. FASEB J. 2002, 16, 1775–1785. [Google Scholar] [CrossRef]
- Sutton, T.; Minnion, M.; Barbarino, F.; Koster, G.; Fernandez, B.O.; Cumpstey, A.; Wischmann, P.; Madhani, M.; Frenneaux, M.; Postle, A.; et al. A robust and versatile mass spectrometry platform for comprehensive assessment of the thiol redox metabolome. Redox Biol. 2018, 16, 359–380. [Google Scholar] [CrossRef] [PubMed]
- Koning, A.M.; Meijers, W.C.; Pasch, A.; Leuvenink, H.; Frenay, A.-R.S.; Dekker, M.M.; Feelisch, M.; de Boer, R.A.; van Goor, H. Serum free thiols in chronic heart failure. Pharmacol. Res. 2016, 111, 452–458. [Google Scholar] [CrossRef] [Green Version]
- Tsikas, D. Assessment of lipid peroxidation by measuring malondialdehyde (MDA) and relatives in biological samples: Analytical and biological challenges. Anal. Biochem. 2017, 524, 13–30. [Google Scholar] [CrossRef] [PubMed]
- De Leon, J.A.D.; Borges, C.R. Evaluation of Oxidative Stress in Biological Samples Using the Thiobarbituric Acid Reactive Substances Assay. J. Vis. Exp. 2020, 12, 159. [Google Scholar] [CrossRef]
- Juniper, E.F.; Buist, A.S.; Cox, F.M.; Ferrie, P.J.; King, D.R. Validation of a Standardized Version of the Asthma Quality of Life Questionnaire. Chest 1999, 115, 1265–1270. [Google Scholar] [CrossRef]
- Wanger, J.; Clausen, J.L.; Coates, A.; Pedersen, O.F.; Brusasco, V.; Burgos, F.; Casaburi, R.; Crapo, R.; Enright, P.; Van Der Grinten, C.P.M.; et al. Standardisation of the measurement of lung volumes. Eur. Respir. J. 2005, 26, 511–522. [Google Scholar] [CrossRef]
- Azim, A.; Mistry, H.; Freeman, A.; Barber, C.; Newell, C.; Gove, K.; Thirlwall, Y.; Harvey, M.; Bentley, K.; Knight, D.; et al. Protocol for the Wessex AsThma CoHort of difficult asthma (WATCH): A pragmatic real-life longitudinal study of difficult asthma in the clinic. BMC Pulm. Med. 2019, 19, 99. [Google Scholar] [CrossRef]
- Juniper, E.F.; Svensson, K.; Mörk, A.-C.; Ståhl, E. Measurement properties and interpretation of three shortened versions of the asthma control questionnaire. Respir. Med. 2005, 99, 553–558. [Google Scholar] [CrossRef] [Green Version]
- Kothmann, E.; Batterham, A.M.; Owen, S.J.; Turley, A.J.; Cheesman, M.; Parry, A.; Danjoux, G. Effect of short-term exercise training on aerobic fitness in patients with abdominal aortic aneurysms: A pilot study. Br. J. Anaesth. 2009, 103, 505–510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oberfeld, D.; Franke, T. Evaluating the robustness of repeated measures analyses: The case of small sample sizes and nonnormal data. Behav. Res. Methods 2012, 45, 792–812. [Google Scholar] [CrossRef] [PubMed]
- Azim, A.; Freeman, A.; Lavenu, A.; Mistry, H.; Haitchi, H.M.; Newell, C.; Cheng, Y.; Thirlwall, Y.; Harvey, M.; Barber, C.; et al. New Perspectives on Difficult Asthma; Sex and Age of Asthma-Onset Based Phenotypes. J. Allergy Clin. Immunol. Pract. 2020, 8, 3396–3406.e4. [Google Scholar] [CrossRef] [PubMed]
- Jones, D.P. Redefining Oxidative Stress. Antioxid. Redox Signal. 2006, 8, 1865–1879. [Google Scholar] [CrossRef]
- Sawa, T.; Ono, K.; Tsutsuki, H.; Zhang, T.; Ida, T.; Nishida, M.; Akaike, T. Reactive Cysteine Persulphides: Occurrence, Biosynthesis, Antioxidant Activity, Methodologies, and Bacterial Persulphide Signalling. Adv. Microb. Physiol. 2018, 72, 1–28. [Google Scholar] [CrossRef]
- Freeman, A.T.; Hill, D.; Newell, C.; Moyses, H.; Azim, A.; Knight, D.; Presland, L.; Harvey, M.; Haitchi, H.M.; Watson, A.; et al. Patient perceived barriers to exercise and their clinical associations in difficult asthma. Asthma Res. Pract. 2020, 6, 5. [Google Scholar] [CrossRef]
- de Sousa, C.V.; Sales, M.M.; Rosa, T.S.; Lewis, J.E.; de Andrade, R.V.; Simões, H.G. The Antioxidant Effect of Exercise: A Systematic Review and Meta-Analysis. Sports Med. 2017, 47, 277–293. [Google Scholar] [CrossRef] [PubMed]
- Fitzpatrick, A.M.; Teague, W.G.; Holguin, F.; Yeh, M.; Brown, L.A.S. Airway glutathione homeostasis is altered in children with severe asthma: Evidence for oxidant stress. J. Allergy Clin. Immunol. 2009, 123, 146–152.e8. [Google Scholar] [CrossRef] [Green Version]
- Fitzpatrick, A.M.; Teague, W.G.; Burwell, L.; Brown, M.S.; Brown, L.A.S. NIH/NHLBI Severe Asthma Research Program Glutathione Oxidation Is Associated with Airway Macrophage Functional Impairment in Children with Severe Asthma. Pediatr. Res. 2011, 69, 154–159. [Google Scholar] [CrossRef] [Green Version]
- McKenna, H.T.; O’Brien, K.A.; Fernandez, B.O.; Minnion, M.; Tod, A.; McNally, B.D.; West, J.A.; Griffin, J.L.; Grocott, M.P.; Mythen, M.G.; et al. Divergent trajectories of cellular bioenergetics, intermediary metabolism and systemic redox status in survivors and non-survivors of critical illness. Redox Biol. 2021, 41, 101907. [Google Scholar] [CrossRef]
- Seifi-Skishahr, F.; Damirchi, A.; Farjaminezhad, M.; Babaei, P. Physical Training Status Determines Oxidative Stress and Redox Changes in Response to an Acute Aerobic Exercise. Biochem. Res. Int. 2016, 2016, 3757623. [Google Scholar] [CrossRef] [Green Version]
- Kuo, H.-K.; Yen, C.-J.; Bean, J.F. Levels of homocysteine are inversely associated with cardiovascular fitness in women, but not in men: Data from the National Health and Nutrition Examination Survey 1999–2002. J. Intern. Med. 2005, 258, 328–335. [Google Scholar] [CrossRef] [PubMed]
- Randeva, H.S.; Lewandowski, K.; Drzewoski, J.; Brooke-Wavell, K.; O’Callaghan, C.; Czupryniak, L.; Hillhouse, E.W.; Prelevic, G.M. Exercise Decreases Plasma Total Homocysteine in Overweight Young Women with Polycystic Ovary Syndrome. J. Clin. Endocrinol. Metab. 2002, 87, 4496–4501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iglesias-Gutiérrez, E.; Egan, B.; Martinez, A.E.D.; Penalvo, J.L.; González-Medina, A.; Camblor, P.M.; O’Gorman, D.; Úbeda, N. Transient Increase in Homocysteine but Not Hyperhomocysteinemia during Acute Exercise at Different Intensities in Sedentary Individuals. PLoS ONE 2012, 7, e51185. [Google Scholar] [CrossRef] [Green Version]
- Avci, A.G.; Emre, A. Homocysteine: A risk factor for the development of cardiovascular events in chronic respiratory diseases. Biomed. Res. Health Sci. Bio. Converg. Technol. 2016, 27, S450–S453. [Google Scholar]
- Vallance, P. Homocysteine in Health and Disease; Carmel, R., Jacobsen, D.W., Eds.; Cambridge University Press: Cambridge, UK, 2001. [Google Scholar]
- Zou, C.-G.; Banerjee, R. Homocysteine and Redox Signaling. Antioxid. Redox Signal. 2005, 7, 547–559. [Google Scholar] [CrossRef]
- Vitvitsky, V.; Mosharov, E.; Tritt, M.; Ataullakhanov, F.; Banerjee, R. Redox regulation of homocysteine-dependent glutathione synthesis. Redox Rep. 2003, 8, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Morton, R.J.; Henry, R.L.; Thomas, P.S. Exhaled breath condensate nitrite/nitrate and pH in relation to pediatric asthma control and exhaled nitric oxide. Pediatr Pulmonol. 2006, 41, 929–936. [Google Scholar]
- Zetterquist, W.; Marteus, H.; Hedlin, G.; Alving, K. Increased exhaled nitrite in children with allergic asthma is not related to nitric oxide formation. Clin. Respir. J. 2008, 2, 166–174. [Google Scholar] [CrossRef] [PubMed]
- Malinovschi, A.; Pizzimenti, S.; Sciascia, S.; Heffler, E.; Badiu, I.; Rolla, G. Exhaled breath condensate nitrates, but not nitrites or FENO, relate to asthma control. Respir. Med. 2011, 105, 1007–1013. [Google Scholar] [CrossRef] [Green Version]
- Rassaf, T.; Lauer, T.; Heiss, C.; Balzer, J.; Mangold, S.; Leyendecker, T.; Rottler, J.; Drexhage, C.; Meyer, C.; Kelm, M. Nitric oxide synthase-derived plasma nitrite predicts exercise capacity. Br. J. Sports Med. 2007, 41, 669–673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deveci, F.; Ilhan, N.; Turgut, T.; Akpolat, N.; Kirkil, G.; Muz, M. Glutathione and nitrite in induced sputum from patients with stable and acute asthma compared with controls. Ann. Allergy Asthma Immunol. 2004, 93, 91–97. [Google Scholar] [CrossRef]
- Ganas, K.; Loukides, S.; Papatheodorou, G.; Panagou, P.; Kalogeropoulos, N. Total nitrite/nitrate in expired breath condensate of patients with asthma. Respir. Med. 2001, 95, 649–654. [Google Scholar] [CrossRef] [Green Version]
- Maeda, S.; Miyauchi, T.; Kakiyama, T.; Sugawara, J.; Iemitsu, M.; Irukayama-Tomobe, Y.; Murakami, H.; Kumagai, Y.; Kuno, S.; Matsuda, M. Effects of exercise training of 8 weeks and detraining on plasma levels of endothelium-derived factors, endothelin-1 and nitric oxide, in healthy young humans. Life Sci. 2001, 69, 1005–1016. [Google Scholar] [CrossRef]
- Maeda, S.; Tanabe, T.; Otsuki, T.; Sugawara, J.; Iemitsu, M.; Miyauchi, T.; Kuno, S.; Ajisaka, R.; Matsuda, M. Moderate Regular Exercise Increases Basal Production of Nitric Oxide in Elderly Women. Hypertens. Res. 2004, 27, 947–953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, W.; Chen, Y.; Hazen, S.L. Eosinophil peroxidase nitrates protein tyrosyl residues. Implications for oxidative damage by nitrating intermediates in eosinophilic inflammatory disorders. J. Biol. Chem. 1999, 274, 25933–25944. [Google Scholar] [CrossRef] [Green Version]
- Gaston, B.; Singel, D.; Doctor, A.; Stamler, J.S. S-Nitrosothiol Signaling in Respiratory Biology. Am. J. Respir. Crit. Care Med. 2006, 173, 1186–1193. [Google Scholar] [CrossRef] [Green Version]
- Jones, A.M.; Vanhatalo, A.; Seals, D.R.; Rossman, M.J.; Piknova, B.; Jonvik, K.L. Dietary Nitrate and Nitric Oxide Metabolism: Mouth, Circulation, Skeletal Muscle, and Exercise Performance. Med. Sci. Sports Exerc. 2021, 53, 280–294. [Google Scholar] [CrossRef]
- Piknova, B.; Park, J.W.; Lam, K.K.; Schechter, A.N. Nitrate as a source of nitrite and nitric oxide during exercise hyperemia in rat skeletal muscle. Nitric Oxide 2016, 55–56, 54–61. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Samouilov, A.; Liu, X.; Zweier, J.L. Characterization of the Magnitude and Kinetics of Xanthine Oxidase-Catalyzed Nitrate Reduction: Evaluation of Its Role in Nitrite and Nitric Oxide Generation in Anoxic Tissues. Biochemistry 2003, 42, 1150–1159. [Google Scholar] [CrossRef]
- Feelisch, M.; Fernandez, B.O.; Bryan, N.S.; Garcia-Saura, M.F.; Bauer, S.; Whitlock, D.R.; Ford, P.C.; Janero, D.R.; Rodriguez, J.; Ashrafian, H. Tissue Processing of Nitrite in Hypoxia: An Intricate Interplay of Nitric Oxide-Generating and -Scavenging Systems. J. Biol. Chem. 2008, 283, 33927–33934. [Google Scholar] [CrossRef] [Green Version]
- Totzeck, M.; Hendgen-Cotta, U.B.; Rammos, C.; Frommke, L.-M.; Knackstedt, C.; Predel, H.-G.; Kelm, M.; Rassaf, T. Higher endogenous nitrite levels are associated with superior exercise capacity in highly trained athletes. Nitric Oxide 2012, 27, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Sriboonyong, T.; Kawamatawong, T.; Sriwantana, T.; Srihirun, S.; Titapiwatanakun, V.; Vivithanaporn, P.; Pornsuriyasak, P.; Sibmooh, N.; Kamalaporn, H. Efficacy and safety of inhaled nebulized sodium nitrite in asthmatic patients. Pulm. Pharmacol. Ther. 2021, 66, 101984. [Google Scholar] [CrossRef] [PubMed]
- Lundberg, J.O.; Weitzberg, E.; Gladwin, M.T. The nitrate-nitrite-nitric oxide pathway in physiology and therapeutics. Nat. Rev. Drug Discov. 2008, 7, 156–167. [Google Scholar] [CrossRef]
- Bryan, N.S.; Fernandez, B.O.; Bauer, S.M.; Garcia-Saura, M.F.; Milsom, A.B.; Rassaf, T.; Maloney, E.R.; Bharti, A.; Rodriguez, J.; Feelisch, M. Nitrite is a signaling molecule and regulator of gene expression in mammalian tissues. Nat. Chem. Biol. 2005, 1, 290–297. [Google Scholar] [CrossRef]
- Feelisch, M.; Akaike, T.; Griffiths, K.; Ida, T.; Prysyazhna, O.; Goodwin, J.J.; Gollop, N.D.; Fernandez, B.O.; Minnion, M.; Cortese-Krott, M.M.; et al. Long-lasting blood pressure lowering effects of nitrite are NO-independent and mediated by hydrogen peroxide, persulfides, and oxidation of protein kinase G1α redox signalling. Cardiovasc. Res. 2020, 116, 51–62. [Google Scholar] [CrossRef] [Green Version]
- Stucki, D.; Steinhausen, J.; Westhoff, P.; Krahl, H.; Brilhaus, D.; Massenberg, A.; Weber, A.P.M.; Reichert, A.S.; Brenneisen, P.; Stahl, W. Endogenous Carbon Monoxide Signaling Modulates Mitochondrial Function and Intracellular Glucose Utilization: Impact of the Heme Oxygenase Substrate Hemin. Antioxidants 2020, 9, 652. [Google Scholar] [CrossRef] [PubMed]
- de Souza, A.R.W.; Gallo, D.; Lee, G.R.; Katsuyama, E.; Schaufler, A.; Weber, J.; Csizmadia, E.; Tsokos, G.C.; Koch, L.G.; Britton, S.L.; et al. Skeletal muscle heme oxygenase-1 activity regulates aerobic capacity. Cell Rep. 2021, 35, 109018. [Google Scholar] [CrossRef]
- Adenan, D.M.; Jaafar, Z.; Jayapalan, J.; Aziz, A.A. Plasma antioxidants and oxidative stress status in obese women: Correlation with cardiopulmonary response. Peer J. 2020, 8, e9230. [Google Scholar] [CrossRef]
- Sakelliou, A.; Fatouros, I.G.; Athanailiidis, I.; Tsoukas, D.; Chatzinikolaou, A.; Draganidis, D.; Jamurtas, A.Z.; Liacos, C.; Papassotiriou, I.; Mandalidis, D.; et al. Evidence of a Redox-Dependent Regulation of Immune Responses to Exercise-Induced Inflammation. Oxidative Med. Cell. Longev. 2016, 2016, 2840643. [Google Scholar] [CrossRef] [Green Version]
- Watson, A.; Wilkinson, T.M.A.; Freeman, A. Evidence Around the Impact of Pulmonary Rehabilitation and Exercise on Redox Status in COPD: A Systematic Review. Front. Sports Act. Living 2021, 3, 335. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristic | Number (%) or Median [IQR] |
|---|---|
| Female sex | 6 (100) |
| Age (years) | 31.3 ± 10 |
| Never smoker | 5 (83) |
| BMI (kg/m2) | 27.6 [22.26, 30.66] |
| Peripheral blood eosinophil count | 0.25 [0.2, 0.73] |
| FeNO (ppb) | 50.75 [27.25, 93] |
| Co-morbidities | Number (%) |
| Atopy | 5 (83) |
| Anxiety and depression | 2 (33) |
| Urticaria and angioedema | 1 (16%) |
| Anaphylaxis | 1 (16%) |
| Dysfunctional breathing | 1 (16%) |
| Pulmonary Function | Median [IQR] |
| FEV1 % predicted | 89 [78.75, 94.5] |
| FVC % predicted | 100.5 [91.75, 1.3] |
| FEV1/FVC | 77 [73.75, 82.25] |
| FEF 25–75 % predicted | 68.5 [31.30, 85.25] |
| % change FEV1 post BD | 5 [3, 13.25] |
| Asthma Medication | Number (%) or mean ± SD |
| on ICS | 4 (66%) |
| ICS dose (BDP equivalent µg/day) | 483 ± 371 |
| on LABA | 2 |
| LABA dose (formoterol equivalent µg/day) | 7.2 ± 10.73 |
| on Montelukast | 1 (16%) |
| Variables | Baseline Median (IQR) | Post Intervention Median (IQR) | p Value |
|---|---|---|---|
| Anthropometric Values | |||
| BMI Height (cm) Weight (kg) | 27.7 (22.3, 30.7) 171.5 (164.3, 172.1) 78.2 (58.8, 91.8) | 27.6 (22, 29.6) n.d. n.d. | 0.136 - - |
| Fat-free mass (%) | 62.9 (58.3, 67.5) | 60.7 (56.2, 67.5) | 1 |
| Fat mass (%) | 36.6 (32.5, 41.7) | 36.8 (32.5, 42.1) | 0.528 |
| Visceral Adipose Tissue (litres) | 0.83 (0.45, 1.23) | 0.74 (0.16, 1) | 0.465 |
| Skeletal Muscle Mass (kg) | 23 (18.8, 27.2) | 23 (18.3, 27.2) | 0.463 |
| Aerobic Capacity | |||
| VO2 peak (mL/kg/min) | 21.6 (19, 28) | 25 (20, 31.5) | 0.058 * |
| Anaerobic Threshold (mL/kg/min) Maximum Workload (watts) | 10.4 (9.4, 14) 161 (135, 205) | 12.6 (10, 15.6) 186 (172, 235) | 0.046 * 0.028 * |
| Variable | Baseline Median (IQR) | Post 12-Week Intervention Median (IQR) | p Value |
|---|---|---|---|
| ACQ 6 Score | 2 (1.8, 2.6) | 1.2 (0.8, 1.5) | 0.028 * |
| AQLQ Total Score | 4.8 (4.1, 5.3) | 5.8 (5.5, 6.3) | 0.046 * |
| AQLQ Symptoms Domain | 4.5 (3.8, 5) | 5.8, 5.1, 6.3) | 0.046 * |
| AQLQ Activity Domain | 5.3 (4.4, 6) | 6.1 5.7, 6.8) | 0.116 |
| AQLQ Emotions Domain | 4.8 (2.7, 5.3) | 6.1 (5.3, 6.5) | 0.027 * |
| AQLQ Environmental Domain | 4.5 (3.6, 5.4) | 5.6 (4.9, 6.6) | 0.043 * |
| Variables | Baseline Median (IQR) | Post Intervention Median (IQR) | p Value |
|---|---|---|---|
| Airway Inflammation | |||
| FeNO (ppb) | 50 (27, 93) | 35 (19, 94) | 0.753 |
| Clinical inflammatory markers | |||
| WCC × 109/L | 7.2 (5.6, 9.1) | 6.7 (4.0, 7.5) | 0.046 * |
| Neutrophils × 109/L | 6.7 (2.7, 5.8) | 3.9 (1.9, 4.6) | 0.054 |
| Eosinophils × 109/L | 0.25 (0.2, 0.73) | 0.2 (0.18, 0.43) | 0.007 * |
| Lymphocytes × 109/L | 2.2 (1.6, 2.7) | 1.8 (1.7, 2.2) | 0.049 * |
| Monocytes × 109/L | 0.5 (0.38, 0.55) | 0.45 (0.45, 0.53) | 0.414 |
| CRP mg/L | 3 (1.5, 6.3) | 1 (0.75, 6.8) | 0.340 |
| Cytokines (pg/mL) | |||
| CCL11/eotaxin | 173.0 (138.5, 214.0) | 153.97 (134.0, 180.1) | 0.046 * |
| IL-5 | 7.24 (6.39, 10.33) | 5.34 (4.04, 7.37) | 0.046 * |
| TNFα | 15.63 (9.75, 15.63) | 9.96 (5.19, 12.53) | 0.046 * |
| IFNγ | 17.89 (10.87, 20.32) | 11.9 (7.48, 12.91) | 0.046 * |
| IL1-ra | 644.3 (328.0, 1077.8) | 469.9 (307.1, 900.8) | 0.116 |
| IL-6 | 8.32 (6.35, 12.48) | 6.60 (5.40, 8.74) | 0.115 |
| IL-10 | 79.20 (65.89, 96.72) | 72.92 (58.80, 78.7) | 0.173 |
| IL-13 | 66. 81 (58.95, 76.23) | 55.72 (48.70, 64.85) | 0.141 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Freeman, A.; Cellura, D.; Minnion, M.; Fernandez, B.O.; Spalluto, C.M.; Levett, D.; Bates, A.; Wallis, T.; Watson, A.; Jack, S.; et al. Exercise Training Induces a Shift in Extracellular Redox Status with Alterations in the Pulmonary and Systemic Redox Landscape in Asthma. Antioxidants 2021, 10, 1926. https://doi.org/10.3390/antiox10121926
Freeman A, Cellura D, Minnion M, Fernandez BO, Spalluto CM, Levett D, Bates A, Wallis T, Watson A, Jack S, et al. Exercise Training Induces a Shift in Extracellular Redox Status with Alterations in the Pulmonary and Systemic Redox Landscape in Asthma. Antioxidants. 2021; 10(12):1926. https://doi.org/10.3390/antiox10121926
Chicago/Turabian StyleFreeman, Anna, Doriana Cellura, Magdalena Minnion, Bernadette O. Fernandez, Cosma Mirella Spalluto, Denny Levett, Andrew Bates, Timothy Wallis, Alastair Watson, Sandy Jack, and et al. 2021. "Exercise Training Induces a Shift in Extracellular Redox Status with Alterations in the Pulmonary and Systemic Redox Landscape in Asthma" Antioxidants 10, no. 12: 1926. https://doi.org/10.3390/antiox10121926