
Reactive Oxygen Species as the Brainbox in Malaria Treatment

 , ,
, ,
Abstract
:1. Introduction
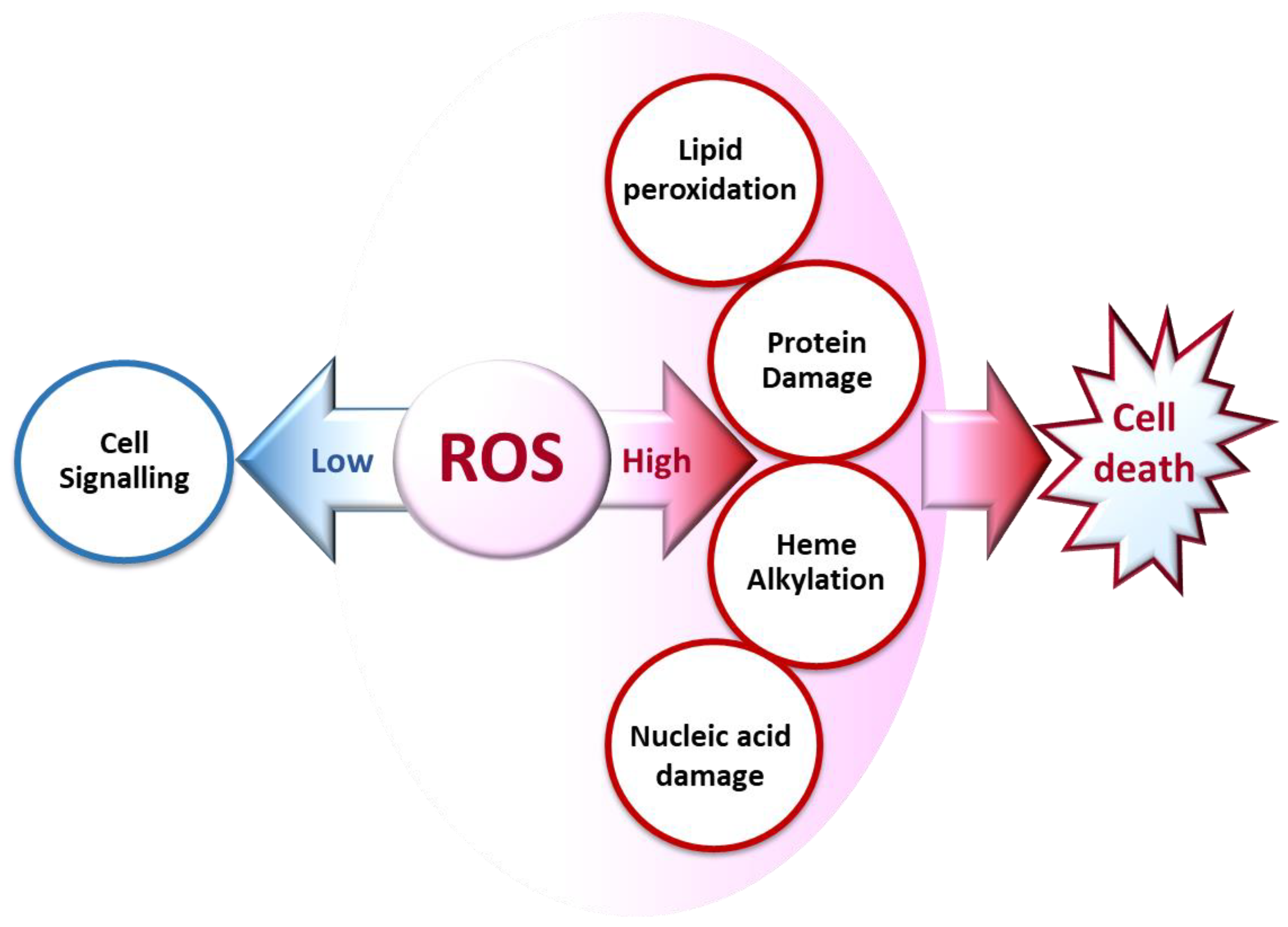
2. Defining ROS in Living Cells
3. The Biochemical Impacts of ROS in Living Cells: Is There Any Peculiarity in Malaria?
3.1. Cell Signaling
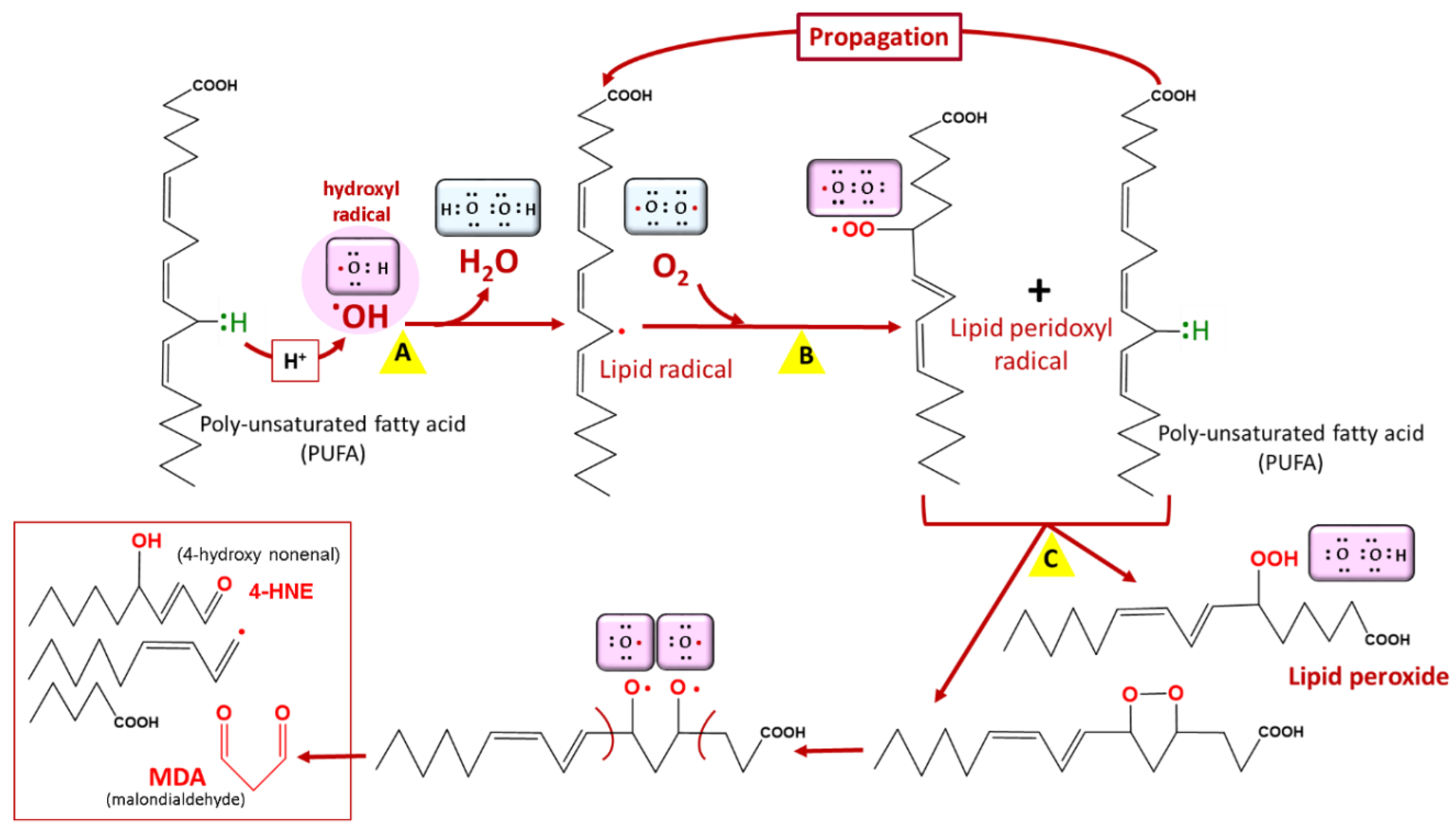
3.2. Lipid Peroxidation
3.3. Protein Damage
- (i)
- by a metal-catalyzed oxidative (MCO) attack targeting the amino acid moiety of arginine, lysine, threonine, and proline (Figure 4);
- (ii)
- by secondary reactions on lysine, cysteine, and histidine with reactive carbonyl derivatives, resulting, among other things, from lipid peroxidation.
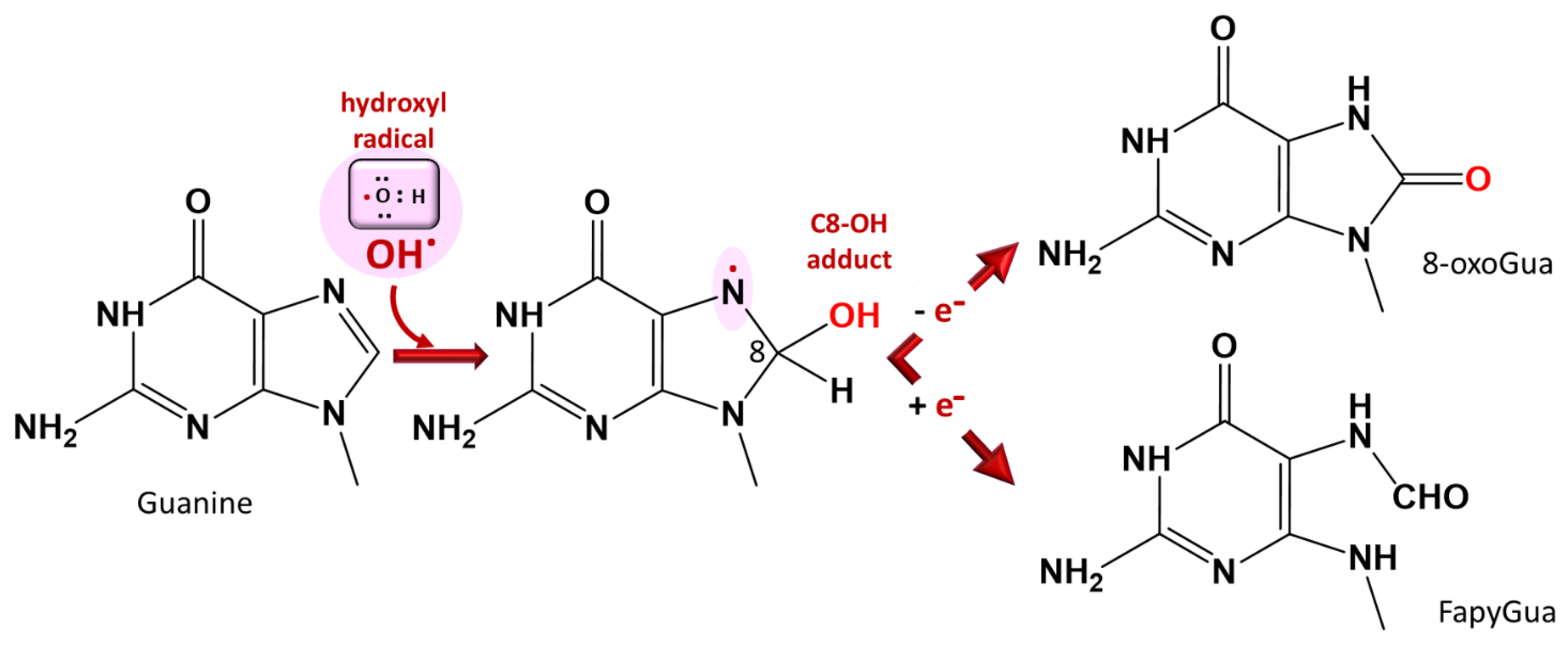
3.4. Nucleic Acid Damage
4. Sources and Management of ROS in Plasmodium-Infected Erythrocytes under Steady State
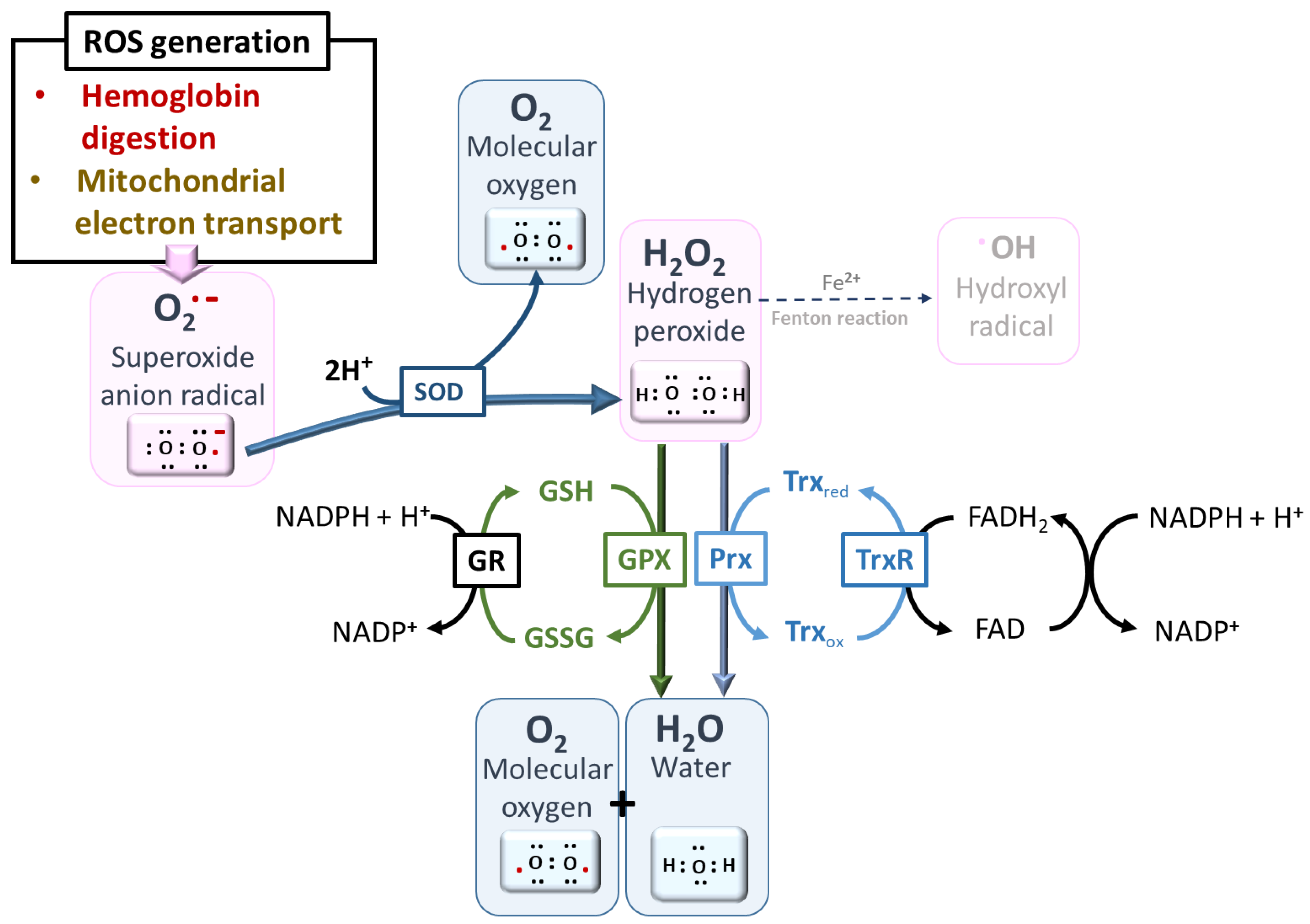
4.1. ROS Production from Mitochondrial Electron Transport Chain
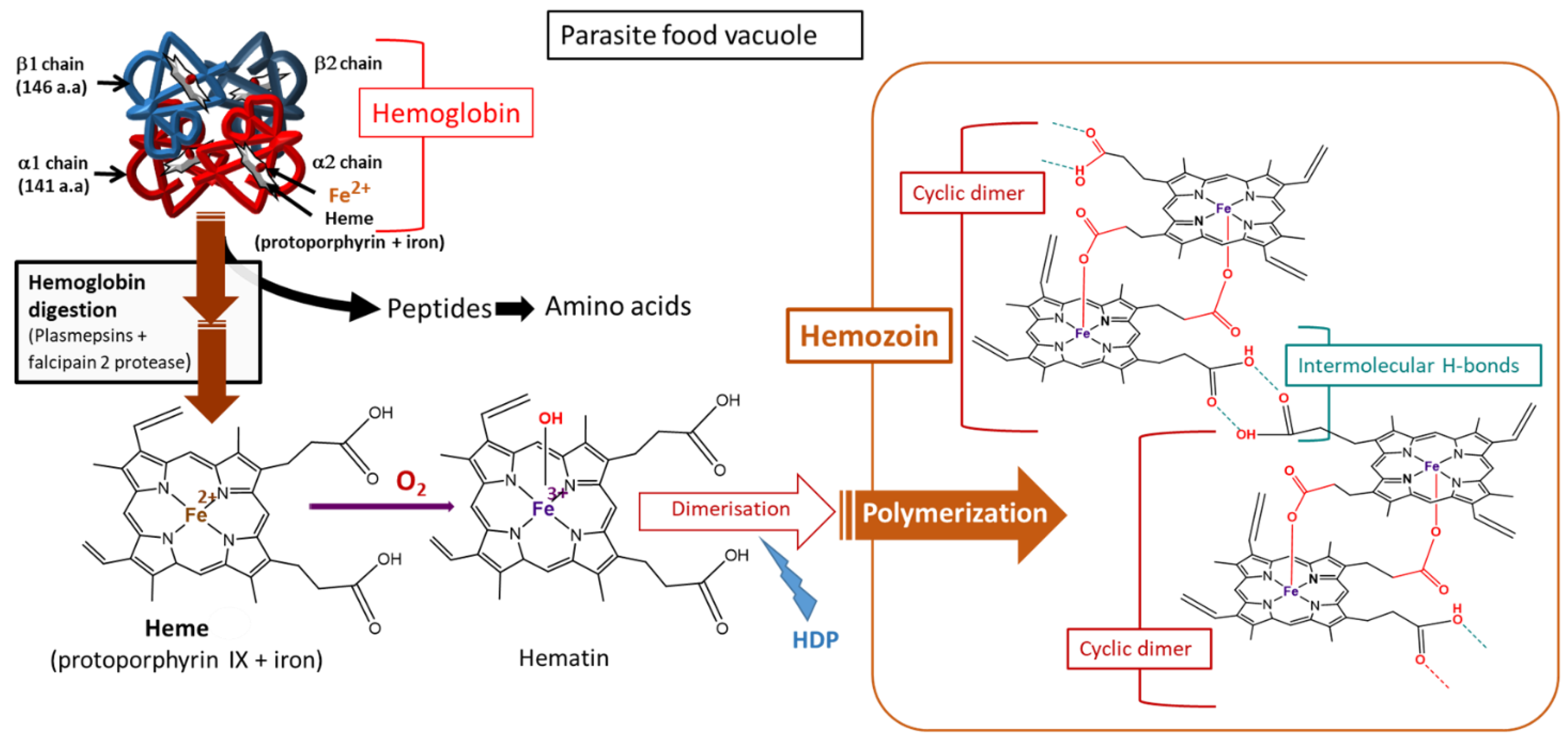
4.2. ROS Production from Hemoglobin Digestion


4.3. Management of ROS by Plasmodium under Steady State: The Antioxidant Machinery
- -
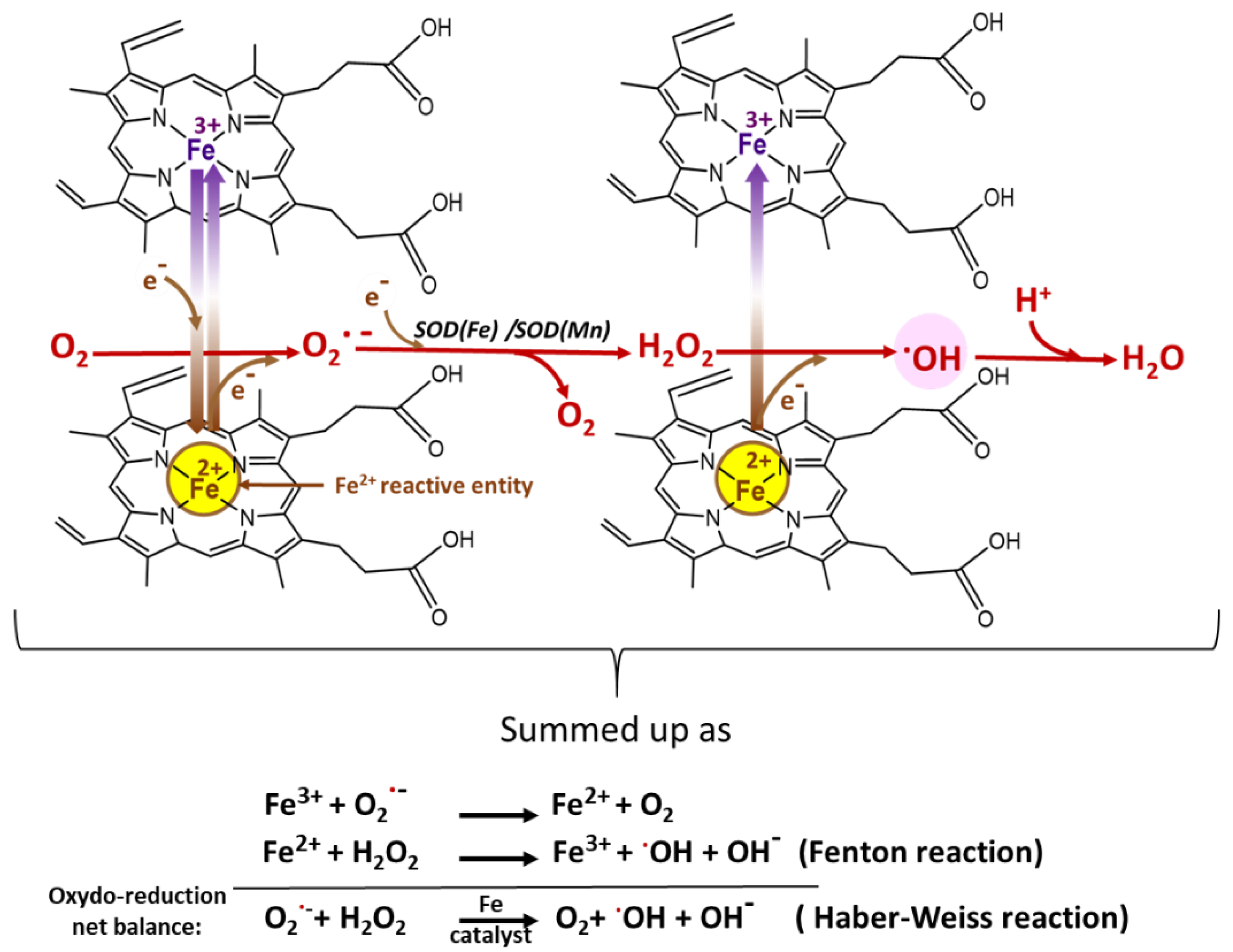
- The SODs act fundamentally to enable the spontaneous dismutation of O2•− to H2O2, which can then be reduced by other enzymes to water, preventing its reaction with iron (Fe2+) to form highly toxic •OH. Plasmodium expresses two forms of SOD: cytosolic PfSOD-1 (Fe-SOD) and mitochondrial PfSOD-2 (Mn-SOD). The uptake of SOD from the host may also be complementary [52,72].
- -
- Glutathione, a tripeptide composed of cysteine, glycine, and glutamate, plays a vital role in the defense system of malaria parasites. Although the level of heme that escapes polymerization is only 5%, it can still mount oxidative stress, which can be eliminated by GSH-dependent mechanisms [45,46,73]. GSH is generated by the reduction of GSSG by glutathione reductase, which can be amplified by other proteins with thiol groups, such as thioredoxin (Trx) [74] (Figure 8). This could be further boosted by the de novo synthesis of GSH, a predominant pathway under oxidative stress [75,76]. Suggestions of possible uptake from the host were rebuffed by the findings of Patzewitz et al., who demonstrated that GSH uptake from the host is not statistically significant [77]. In addition to the nonenzymatic degradation of heme by GSH to generate nonheme iron, Plasmodium also possesses a thiol enzyme, glutathione-S-transferase (GST), which binds and sequesters heme (Fe2+) [78]. To affect its detoxification roles, GSH also serves as a cofactor for GSH-dependent enzymes such as GST, which helps in the reduction of H2O2 and glutathione-peroxidase-like proteins. Overall, GSH acts as a redox buffer to provide redox homeostasis [79]. This is essential in preventing the escalation of ROS generation, which is often aided by the redox cycling of transition metals such as iron.
- -
- Thioredoxins (Trxs) are disulfide oxidoreductases, expressed in all organisms, that interact with a myriad of proteins through their cysteine moiety to donate electrons [80,81]. Trxs keep biomolecules in their reduced and active conformations, consequently promoting parasite survival under oxidative stress. The regeneration of reduced thioredoxin is achieved through the action of thioredoxin reductase (a FAD-dependent enzyme) using NADPH [82]. Three isoforms of Trx have been characterized: a cytosolic isoform, Trx1, and two isoforms in the apicoplast, Trx2 and Trx3 [50].
- -
- Plasmodium lacks catalase and glutathione peroxidase and therefore will likely depend greatly on peroxiredoxins (Prx) to reduce H2O2 generated by SOD [52,53]. P. falciparum expresses five different isoforms of Prx that are strategically localized in the cytosol, mitochondria, and perhaps the apicoplast. These are 1-Cys Prx, two typical 2-Cys Prxs (Prx1 and Prx2), a 1-Cys antioxidant protein (AOP), and a GSH peroxidase-like thioredoxin peroxidase (TPxGl) [50]. It is known that 1-CysPrx and Prx1 are localized in the cytosol, while Prx2 is localized in the mitochondria. The localizations of AOP and TPxGl are not yet fully understood. Prx1, one of the most abundant peroxidases, is constitutively expressed across all blood parasitic stages and has a very high affinity for H2O2, hence being judged to serve a housekeeping role [53,83,84]. Prxs are kept in reduced form through reduction by thioredoxin-dependent systems, comprising three Trxs and Trx-like proteins (TLPs), Tlp1 and Tlp2, which have been identified genomically in the parasite [50]. The parasite, in addition to the expressed Trxs, has approximately 50% of its thioredoxin peroxidase activity depending on the imported human Prx-2 (hPrx-2) [85]. Moreover, it has been demonstrated that H2O2 can diffuse into the host and be handled by the host’s catalase enzyme [86]. The import of human antioxidant machinery may be applicable to other antioxidants, although it has not been fully verified.
- -
- NADPH is involved in maintaining vital antioxidant enzymes, such as glutathione and thioredoxin reductases, in their active conformations. However, the accumulation of heme shifts the equilibrium toward the oxidized form NADP+, increasing the pressure on the reduced form NADPH necessary to reduce the oxidized glutathione (GSSG) to glutathione (GSH). This can lead to the suppression of the main antioxidant mechanisms, such as glutathione reductase, thioredoxin reductase (TrxR), glyceraldehyde-3-phosphate dehydrogenase, and other proteins, leading to the elevation of oxidative stress [87].
- -
- Vitamin B6, in addition to playing several biochemical roles, has also been seen to act as a potent antioxidant [88,89]. The enzymes governing vitamin B6 synthesis are elevated in parasites exposed to oxidative stress, e.g., generated by methylene blue and hence seen as a potential pharmacological target [49]. Although its antioxidant mechanism is still subject to debate, it is believed to scavenge ROS and prevent lipid peroxidation [90,91,92].
5. ROS Production in Plasmodium-Infected Erythrocytes under Antimalarial Treatment
5.1. Mode of Action of Chloroquine and Other Quinolines
5.2. Mode of Action of Atovaquone and Hydroxynaphtoquinones
5.3. Mode of Action of Artemisinin: The Role of Endoperoxide
5.3.1. Activation of the Endoperoxide to Generate ROS
5.3.2. Depolarization of the Mitochondrial Membrane Potential
5.4. Artemisinin-Based Combination Therapies
5.5. ROS Evasive Mechanisms under Treatment
5.5.1. Preventive Mechanisms
5.5.2. Reductive Mechanisms
5.5.3. Reparative Mechanisms
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- WHO World Malaria Report 2020: 20 Years of Global Progress and Challenges. Available online: https://www.who.int/teams/global-malaria-programme/reports/world-malaria-report-2020 (accessed on 30 November 2020).
- Bilgin, R.; Yalcin, M.S.; Yucebilgic, G.; Koltas, I.S.; Yazar, S. Oxidative stress in vivax malaria. Korean J. Parasitol. 2012, 50, 375–377. [Google Scholar] [CrossRef] [PubMed]
- Becker, K.; Tilley, L.; Vennerstrom, J.L.; Roberts, D.; Rogerson, S.; Ginsburg, H. Oxidative stress in malaria parasite-infected erythrocytes: Host-parasite interactions. Int. J. Parasitol. 2004, 34, 163–189. [Google Scholar] [CrossRef] [PubMed]
- Erel, O.; Vural, H.; Aksoy, N.; Aslan, G.; Ulukanligil, M. Oxidative stress of platelets and thrombocytopenia in patients with vivax malaria. Clin. Biochem. 2001, 34, 341–344. [Google Scholar] [CrossRef]
- WHO. World Malaria Report 2019; WHO: Geneva, Switzerland, 2019; ISBN 9789241565721. [Google Scholar]
- Sahu, M.; Tediosi, F.; Noor, A.M.; Aponte, J.J.; Fink, G. Health systems and global progress towards malaria elimination, 2000–2016. Malar. J. 2020, 19, 141. [Google Scholar] [CrossRef]
- Dako-Gyeke, M.; Kofie, H.M. Factors influencing prevention and control of malaria among pregnant women resident in Urban slums, Southern Ghana. Afr. J. Reprod. Health 2015, 19, 44–53. [Google Scholar]
- Thomas, J.A.; Tan, M.S.Y.; Bisson, C.; Borg, A.; Umrekar, T.R.; Hackett, F.; Hale, V.L.; Vizcay-Barrena, G.; Fleck, R.A.; Snijders, A.P.; et al. A protease cascade regulates release of the human malaria parasite Plasmodium falciparum from host red blood cells. Nat. Microbiol. 2018, 3, 447–455. [Google Scholar] [CrossRef]
- Yewhalaw, D.; Wassie, F.; Steurbaut, W.; Spanoghe, P.; Van Bortel, W.; Denis, L.; Tessema, D.A.; Getachew, Y.; Coosemans, M.; Duchateau, L.; et al. Multiple Insecticide Resistance: An Impediment to Insecticide-Based Malaria Vector Control Program. PLoS ONE 2011, 6, e16066. [Google Scholar] [CrossRef] [Green Version]
- Shibeshi, M.A.; Kifle, Z.D.; Atnafie, S.A. Antimalarial Drug Resistance and Novel Targets for Antimalarial Drug Discovery. Infect. Drug Resist. 2020, 13, 4047–4060. [Google Scholar] [CrossRef]
- Payne, D. Spread of chloroquine resistance in Plasmodium falciparum. Parasitol. Today 1987, 3, 241–246. [Google Scholar] [CrossRef]
- Noedl, H.; Se, Y.; Schaecher, K.; Smith, B.L.; Socheat, D.; Fukuda, M.M. Evidence of artemisinin-resistant malaria in Western Cambodia. N. Engl. J. Med. 2008, 359, 2619–2620. [Google Scholar] [CrossRef]
- Dondorp, A.M.; Nosten, F.; Yi, P.; Das, D.; Phyo, A.P.; Tarning, J.; Lwin, K.M.; Ariey, F.; Hanpithakpong, W.; Lee, S.J.; et al. Artemisinin Resistance in Plasmodium falciparum Malaria. N. Engl. J. Med. 2009, 361, 455–467. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.R.; Trush, M. Defining ROS in Biology and Medicine. React. Oxyg. Species 2016, 1, 9–21. [Google Scholar] [CrossRef] [Green Version]
- Santo, A.; Zhu, H.; Li, Y.R. Free Radicals: From Health to Disease. React. Oxyg. Species 2016, 2, 245–263. [Google Scholar] [CrossRef]
- Egwu, C.O.; Tsamesidis, I.; Pério, P.; Augereau, J.-M.; Benoit-Vical, F.; Reybier, K. Superoxide: A major role in the mechanism of action of essential antimalarial drugs. Free Radic. Biol. Med. 2021, 167, 271–275. [Google Scholar] [CrossRef]
- Mohring, F.; Pretzel, J.; Jortzik, E.; Becker, K. The redox systems of Plasmodium falciparum and Plasmodium vivax: Comparison, in silico analyses and inhibitor studies. Curr. Med. Chem. 2014, 21, 1728–1756. [Google Scholar] [CrossRef]
- Hancock, J.T.; Desikan, R.; Neill, S.J. Role of reactive oxygen species in cell signalling pathways. Biochem. Soc. Trans. 2001, 2, 345–350. [Google Scholar] [CrossRef]
- Mittal, M.; Siddiqui, M.R.; Tran, K.; Reddy, S.P.; Malik, A.B. Reactive Oxygen Species in Inflammation and Tissue Injury. Antioxid. Redox Signal. 2014, 20, 1126. [Google Scholar] [CrossRef] [Green Version]
- Stefanatos, R.; Sanz, A. The role of mitochondrial ROS in the aging brain. FEBS Lett. 2018, 592, 743–758. [Google Scholar] [CrossRef] [Green Version]
- Sugamura, K.; Keaney, J.F. Reactive oxygen species in cardiovascular disease. Free Radic. Biol. Med. 2011, 51, 978–992. [Google Scholar] [CrossRef] [Green Version]
- Deavall, D.G.; Martin, E.A.; Horner, J.M.; Roberts, R. Drug-induced oxidative stress and toxicity. J. Toxicol. 2012, 2012, 645460. [Google Scholar] [CrossRef] [Green Version]
- Thannickal, V.J.; Fanburg, B.L. Reactive oxygen species in cell signaling. Am. J. Physiol. Lung Cell. Mol. Physiol. 2000, 279, L1005–L1028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahbari, M.; Rahlfs, S.; Jortzik, E.; Bogeski, I.; Becker, K. H2O2 dynamics in the malaria parasite Plasmodium falciparum. PLoS ONE 2017, 12, e0174837. [Google Scholar] [CrossRef]
- Sampson, C.; Keens, R.H.; Kattnig, D.R. On the magnetosensitivity of lipid peroxidation: Two- versus three-radical dynamics. Phys. Chem. Chem. Phys. 2019, 21, 13526–13538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hartwig, C.L.; Rosenthal, A.S.; D’Angelo, J.; Griffin, C.E.; Posner, G.H.; Cooper, R.A. Accumulation of artemisinin trioxane derivatives within neutral lipids of Plasmodium falciparum malaria parasites is endoperoxide-dependent. Biochem. Pharmacol. 2009, 77, 322–336. [Google Scholar] [CrossRef] [Green Version]
- Kumura, N.; Furukawa, H.; Onyango, A.N.; Izumi, M.; Nakajima, S.; Ito, H.; Hatano, T.; Kim, H.S.; Wataya, Y.; Baba, N. Different behavior of artemisinin and tetraoxane in the oxidative degradation of phospholipid. Chem. Phys. Lipids 2009, 160, 114–120. [Google Scholar] [CrossRef]
- Fernando, N.; Wickremesinghe, S.; Niloofa, R.; Rodrigo, C.; Karunanayake, L.; De Silva, H.J.; Wickremesinghe, A.R.; Premawansa, S.; Rajapakse, S.; Handunnetti, S.M. Protein carbonyl as a biomarker of oxidative stress in severe leptospirosis, and its usefulness in differentiating leptospirosis from dengue infections. PLoS ONE 2016, 11, e0156085. [Google Scholar] [CrossRef]
- Dalle-Donne, I.; Rossi, R.; Giustarini, D.; Milzani, A.; Colombo, R. Protein carbonyl groups as biomarkers of oxidative stress. Clin. Chim. Acta 2003, 329, 23–38. [Google Scholar] [CrossRef]
- Nyström, T. Role of oxidative carbonylation in protein quality control and senescence. EMBO J. 2005, 24, 1311–1317. [Google Scholar] [CrossRef]
- Fedorova, M.; Bollineni, R.C.; Hoffmann, R. Protein carbonylation as a major hallmark of oxidative damage. Mass Spectrom Rev 2014, 33, 79–97. [Google Scholar] [CrossRef]
- Rosenthal, P.J. Cysteine proteases of malaria parasites. Int. J. Parasitol. 2004, 34, 1489–1499. [Google Scholar] [CrossRef]
- Wu, W.M.; Chen, Y.L.; Zhai, Z.; Xiao, S.H.; Wu, Y.L. Study on the mechanism of action of artemether against schistosomes: The identification of cysteine adducts of both carbon-centred free radicals derived from artemether. Bioorg. Med. Chem. Lett. 2003, 13, 1645–1647. [Google Scholar] [CrossRef]
- Eckstein-Ludwig, U.; Webb, R.J.; Van Goethem, I.D.A.; East, J.M.; Lee, A.G.; Kimura, M.; O’Neill, P.M.; Bray, P.G.; Ward, S.A.; Krishna, S. Artemisinins target the SERCA of Plasmodium falciparum. Nature 2003, 424, 957–961. [Google Scholar] [CrossRef]
- Wang, J.; Xu, C.; Lun, Z.R.; Meshnick, S.R. Unpacking ‘Artemisinin Resistance’. Trends Pharmacol. Sci. 2017, 38, 506–511. [Google Scholar] [CrossRef]
- Jena, N.R. DNA damage by reactive species: Mechanisms, mutation and repair. J. Biosci. 2012, 37, 503–517. [Google Scholar] [CrossRef]
- Cadet, J.; Wagner, J.R. DNA base damage by reactive oxygen species, oxidizing agents, and UV radiation. Cold Spring Harb. Perspect. Biol. 2013, 5, a012559. [Google Scholar] [CrossRef]
- Gopalakrishnan, A.M.; Kumar, N. Antimalarial action of artesunate involves DNA damage mediated by reactive oxygen species. Antimicrob. Agents Chemother. 2015, 59, 317–325. [Google Scholar] [CrossRef] [Green Version]
- Freese, R. Markers of oxidative DNA damage in human interventions with fruit and berries. Nutr. Cancer 2006, 54, 143–147. [Google Scholar] [CrossRef]
- Uhlemann, A.C.; Wittlin, S.; Matile, H.; Bustamante, L.Y.; Krishna, S. Mechanism of antimalarial action of the synthetic trioxolane RBX11160 (OZ277). Antimicrob. Agents Chemother. 2007, 51, 667–672. [Google Scholar] [CrossRef] [Green Version]
- Postma, N.S.; Mommers, E.C.; Eling, W.M.C.; Zuidema, J. Oxidative stress in malaria; implications for prevention and therapy. Pharm. World Sci. 1996, 18, 121–129. [Google Scholar] [CrossRef]
- Percário, S.; Moreira, D.R.; Gomes, B.A.Q.; Ferreira, M.E.S.; Gonçalves, A.C.M.; Laurindo, P.S.O.C.; Vilhena, T.C.; Dolabela, M.F.; Green, M.D. Oxidative stress in Malaria. Int. J. Mol. Sci. 2012, 13, 16346–16372. [Google Scholar] [CrossRef] [Green Version]
- Tiwari, S.; Sharma, N.; Sharma, G.P.; Mishra, N. Redox interactome in malaria parasite Plasmodium falciparum. Parasitol. Res. 2021, 120, 423–434. [Google Scholar] [CrossRef] [PubMed]
- Loria, P.; Miller, S.; Foley, M.; Tilley, L. Inhibition of the peroxidative degradation of haem as the basis of action of chloroquine and other quinoline antimalarials. Biochem. J. 1999, 339, 363–370. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Krugliak, M.; Ginsburg, H. The fate of ferriprotorphyrin IX in malaria infected erythrocytes in conjunction with the mode of action of antimalarial drugs. Mol. Biochem. Parasitol. 1999, 99, 129–141. [Google Scholar] [CrossRef]
- Egan, T.J.; Combrinck, J.M.; Egan, J.; Hearne, G.R.; Marques, H.M.; Ntenteni, S.; Sewell, B.T.; Smith, P.J.; Taylor, D.; Van Schalkwyk, D.A.; et al. Fate of haem iron in the malaria parasite Plasmodium falciparum. Biochem. J. 2002, 365, 343–347. [Google Scholar] [CrossRef]
- Rahlfs, S.; Schirmer, R.H.; Becker, K. The thioredoxin system of Plasmodium falciparum and other parasites. Cell. Mol. Life Sci. 2002, 59, 1024–1041. [Google Scholar] [CrossRef]
- Desakorn, V.; Dondorp, A.M.; Silamut, K.; Pongtavornpinyo, W.; Sahassananda, D.; Chotivanich, K.; Pitisuttithum, P.; Smithyman, A.M.; Day, N.P.J.; White, N.J. Stage-dependent production and release of histidine-rich protein 2 by Plasmodium falciparum. Trans. R. Soc. Trop. Med. Hyg. 2005, 99, 517–524. [Google Scholar] [CrossRef]
- Wrenger, C.; Eschbach, M.L.; Müller, I.B.; Warnecke, D.; Walter, R.D. Analysis of the vitamin B6 biosynthesis pathway in the human malaria parasite Plasmodium falciparum. J. Biol. Chem. 2005, 280, 5242–5248. [Google Scholar] [CrossRef] [Green Version]
- Nickel, C.; Rahlfs, S.; Deponte, M.; Koncarevic, S.; Becker, K. Thioredoxin networks in the malarial parasite Plasmodium falciparum. Antioxid. Redox Signal. 2006, 8, 1227–1239. [Google Scholar] [CrossRef]
- Goldberg, D.E.; Sigala, P.A. Plasmodium heme biosynthesis: To be or not to be essential? PLOS Pathog. 2017, 13, e1006511. [Google Scholar] [CrossRef] [Green Version]
- Müller, S. Redox and antioxidant systems of the malaria parasite Plasmodium falciparum. Mol. Microbiol. 2004, 53, 1291–1305. [Google Scholar] [CrossRef]
- Kawazu, S.; Komaki-Yasuda, K.; Oku, H.; Kano, S. Peroxiredoxins in malaria parasites: Parasitologic aspects. Parasitol. Int. 2008, 57, 1–7. [Google Scholar] [CrossRef]
- Mustacich, D.; Powis, G. Thioredoxin reductase. Biochem. J. 2000, 346, 1–8. [Google Scholar] [CrossRef]
- Slater, A.F.G.; Cerami, A. Inhibition by chloroquine of a novel haem polymerase enzyme activity in malaria trophozoites. Nature 1992, 355, 167–169. [Google Scholar] [CrossRef]
- Gupta, P.; Mehrotra, S.; Sharma, A.; Chugh, M.; Pandey, R.; Kaushik, A.; Khurana, S.; Srivastava, N.; Srivastava, T.; Deshmukh, A.; et al. Exploring Heme and Hemoglobin Binding Regions of Plasmodium Heme Detoxification Protein for New Antimalarial Discovery. J. Med. Chem. 2017, 60, 8298–8308. [Google Scholar] [CrossRef]
- Choi, C.Y.H.; Cerda, J.F.; Hsiu-An, C.; Babcock, G.T.; Marletta, M.A. Spectroscopic characterization of the heme-binding sites in Plasmodium falciparum histidine-rich protein 2. Biochemistry 1999, 38, 16916–16924. [Google Scholar] [CrossRef]
- Sullivan, D.J.; Gluzman, I.Y.; Goldberg, D.E. Plasmodium Hemozoin Formation Mediated by Histidine-Rich Proteins. Science 1996, 271, 219–222. [Google Scholar] [CrossRef]
- Kapishnikov, S.; Grolimund, D.; Schneider, G.; Pereiro, E.; McNally, J.G.; Als-Nielsen, J.; Leiserowitz, L. Unraveling heme detoxification in the malaria parasite by in situ correlative X-ray fluorescence microscopy and soft X-ray tomography. Sci. Rep. 2017, 7, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Sigala, P.A.; Goldberg, D.E. The peculiarities and paradoxes of Plasmodium heme metabolism. Annu. Rev. Microbiol. 2014, 68, 259–278. [Google Scholar] [CrossRef] [Green Version]
- Wallace, W.J.; Houtchens, R.A.; Maxwell, J.C.; Caughey, W.S. Mechanism of autooxidation for hemoglobins and myoglobins. Promotion of superoxide production by protons and anions. J. Biol. Chem. 1982, 257, 4966–4977. [Google Scholar] [CrossRef]
- Carrell, R.W.; Winterbourn, C.C.; Rachmilewitz, E.A. Annotation: Activated oxygen and haemolysis. Br. J. Haematol. 1975, 30, 259–264. [Google Scholar] [CrossRef]
- Thomas, C.; Mackey, M.; Diaz, A.; Cox, D. Hydroxyl radical is produced via the Fenton reaction in submitochondrial particles under oxidative stress: Implications for diseases associated with iron accumulation. Redox Rep. 2009, 14, 102–108. [Google Scholar] [CrossRef] [PubMed]
- Jani, D.; Nagarkatti, R.; Beatty, W.; Angel, R.; Slebodnick, C.; Andersen, J.; Kumar, S.; Rathore, D. HDP—A novel heme detoxification protein from the malaria parasite. PLoS Pathog. 2008, 4, e1000053. [Google Scholar] [CrossRef] [PubMed]
- Lew, V.L.; Tiffert, T.; Ginsburg, H. Excess hemoglobin digestion and the osmotic stability of Plasmodium falciparum-Infected red blood cells. Blood 2003, 101, 4189–4194. [Google Scholar] [CrossRef] [PubMed]
- Mairbäurl, H.; Weber, R.E. Oxygen transport by hemoglobin. Compr. Physiol. 2012, 2, 1463–1489. [Google Scholar] [CrossRef] [PubMed]
- Clark, I.A.; Rockett, K.A. Nitric oxide and parasitic disease. Adv. Parasitol. 1996, 37, 1–56. [Google Scholar] [CrossRef]
- Ghigo, D.; Todde, R.; Ginsburg, H.; Costamagna, C.; Gautret, P.; Bussolino, F.; Ulliers, D.; Giribaldi, G.; Deharo, E.; Gabrielli, G.; et al. Erythrocyte stages of Plasmodium falciparum exhibit a high nitric oxide synthase (NOS) activity and release an NOS-inducing soluble factor. J. Exp. Med. 1995, 182, 677–688. [Google Scholar] [CrossRef] [Green Version]
- Egwu, C.O.; Pério, P.; Augereau, J.-M.; Tsamesidis, I.; Benoit-Vical, F.; Reybier, K. Resistance to artemisinin in falciparum malaria parasites: A redox-mediated phenomenon. Free Radic. Biol. Med. 2021, S0891-5849, 00476–00477. [Google Scholar] [CrossRef]
- Nogueira, F.; Diez, A.; Radfar, A.; Pérez-Benavente, S.; Rosario, V.E.; Puyet, A.; Bautista, J.M. Early transcriptional response to chloroquine of the Plasmodium falciparum antioxidant defence in sensitive and resistant clones. Acta Trop. 2010, 114, 109–115. [Google Scholar] [CrossRef]
- Wang, W.; Huang, P.; Jiang, N.; Lu, H.; Zhang, D.; Wang, D.; Zhang, K.; Wahlgren, M.; Chen, Q. A thioredoxin homologous protein of Plasmodium falciparum participates in erythrocyte invasion. Infect. Immun. 2018, 86, e00289-18. [Google Scholar] [CrossRef] [Green Version]
- Fairfield, A.S.; Meshnick, S.R.; Eaton, J.W. Malaria parasites adopt host cell superoxide dismutase. Science 1983, 221, 764–766. [Google Scholar] [CrossRef]
- Ginsburg, H.; Famin, O.; Zhang, J.; Krugliak, M. Inhibition of glutathione-dependent degradation of heme by chloroquine and amodiaquine as a possible basis for their antimalarial mode of action. Biochem. Pharmacol. 1998, 56, 1305–1313. [Google Scholar] [CrossRef]
- Porras, P.; Pedrajas, J.R.; Martínez-Galisteo, E.; Alicia Padilla, C.; Johansson, C.; Holmgren, A.; Antonio Bárcena, J. Glutaredoxins catalyze the reduction of glutathione by dihydrolipoamide with high efficiency. Biochem. Biophys. Res. Commun. 2002, 295, 1046–1051. [Google Scholar] [CrossRef]
- Patzewitz, E.-M.; Müller, S. Glutathione biosynthesis and metabolism in Plasmodium falciparum. Malar. J. 2010, 9, 37. [Google Scholar] [CrossRef] [Green Version]
- Lüersen, K.; Walter, R.D.; Müller, S. Plasmodium falciparum-infected red blood cells depend on a functional glutathione de novo synthesis attributable to an enhanced loss of glutathione. Biochem. J. 2000, 346, 545–552. [Google Scholar] [CrossRef]
- Patzewitz, E.M.; Wong, E.H.; Müller, S. Dissecting the role of glutathione biosynthesis in Plasmodium falciparum. Mol. Microbiol. 2012, 83, 304–318. [Google Scholar] [CrossRef] [Green Version]
- Harvey, J.W.; Beutler, E. Binding of heme by glutathione S-transferase: A possible role of the erythrocyte enzyme. Blood 1982, 60, 1227–1230. [Google Scholar] [CrossRef]
- Müller, S. Role and regulation of glutathione metabolism in Plasmodium falciparum. Molecules 2015, 20, 10511–10534. [Google Scholar] [CrossRef] [Green Version]
- Hirt, R.P.; Müller, S.; Martin Embley, T.; Coombs, G.H. The diversity and evolution of thioredoxin reductase: New perspectives. Trends Parasitol. 2002, 18, 302–308. [Google Scholar] [CrossRef]
- Bozdech, Z.; Ginsburg, H. Antioxidant defense in Plasmodium falciparum - Data mining of the transcriptome. Malar. J. 2004, 3, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Lillig, C.H.; Holmgren, A. Thioredoxin and related molecules - From biology to health and disease. Antioxid. Redox Signal. 2007, 9, 25–47. [Google Scholar] [CrossRef]
- Yano, K.; Komaki-Yasuda, K.; Kobayashi, T.; Takemae, H.; Kita, K.; Kano, S.; Kawazu, S.I. Expression of mRNAs and proteins for peroxiredoxins in Plasmodium falciparum erythrocytic stage. Parasitol. Int. 2005, 54, 35–41. [Google Scholar] [CrossRef] [PubMed]
- Akerman, S.E.; Müller, S. 2-Cys peroxiredoxin PfTrx-Px1 is involved in the antioxidant defence of Plasmodium falciparum. Mol. Biochem. Parasitol. 2003, 130, 75–81. [Google Scholar] [CrossRef]
- Koncarevic, S.; Rohrbach, P.; Deponte, M.; Krohne, G.; Prieto, J.H.; Yates, J.; Rahlfs, S.; Becker, K. The malarial parasite Plasmodium falciparum imports the human protein peroxiredoxin 2 for peroxide detoxification. Proc. Natl. Acad. Sci. USA 2009, 106, 13323–13328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atamna, H.; Pascarmona, G.; Ginsburg, H. Hexose-monophosphate shunt activity in intact Plasmodium falciparum-infected erythrocytes and in free parasites. Mol. Biochem. Parasitol. 1994, 67, 79–89. [Google Scholar] [CrossRef]
- Campanale, N.; Nickel, C.; Daubenberger, C.A.; Wehlan, D.A.; Gorman, J.J.; Klonis, N.; Becker, K.; Tilley, L. Identification and characterization of heme-interacting proteins in the malaria parasite, Plasmodium falciparum. J. Biol. Chem. 2003, 278, 27354–27361. [Google Scholar] [CrossRef] [Green Version]
- Ehrenshaft, M.; Bilski, P.; Li, M.; Chignell, C.F.; Daub, M.E. A highly conserved sequence is a novel gene involved in de novo vitamin B6 biosynthesis. Proc. Natl. Acad. Sci. USA 1999, 96, 9374–9378. [Google Scholar] [CrossRef] [Green Version]
- Bilski, P.; Li, M.Y.; Ehrenshaft, M.; Daub, M.E.; Chignell, C.F. Vitamin B6 (Pyridoxine) and Its Derivatives Are Efficient Singlet Oxygen Quenchers and Potential Fungal Antioxidants. Photochem. Photobiol. 2000, 71, 129–134. [Google Scholar] [CrossRef]
- Matxain, J.M.; Padro, D.; Ristilä, M.; Strid, Å.; Eriksson, L.A. Evidence of high ·OH radical quenching efficiency by vitamin B 6. J. Phys. Chem. B 2009, 113, 9629–9632. [Google Scholar] [CrossRef]
- Natera, J.; Massad, W.; García, N.A. The role of vitamin B6 as an antioxidant in the presence of vitamin B2-photogenerated reactive oxygen species. A kinetic and mechanistic study. Photochem. Photobiol. Sci. 2012, 11, 938. [Google Scholar] [CrossRef]
- Kannan, K.; Jain, S.K. Effect of vitamin B 6 on oxygen radicals, mitochondrial membrane potential, and lipid peroxidation in H 2 O 2 -treated U937 monocytes. Free Radic. Biol. Med. 2004, 36, 423–428. [Google Scholar] [CrossRef]
- Tsamesidis, I.; Egwu, C.O.; Pério, P.; Augereau, J.M.; Benoit-Vical, F.; Reybier, K. An LC–MS assay to measure superoxide radicals and hydrogen peroxide in the blood system. Metabolites 2020, 10, 175. [Google Scholar] [CrossRef]
- Sullivan, D.J.; Matile, H.; Ridley, R.G.; Goldberg, D.E. A common mechanism for blockade of heme polymerization by antimalarial quinolines. J. Biol. Chem. 1998, 273, 31103–31107. [Google Scholar] [CrossRef] [Green Version]
- Sugioka, Y.; Suzuki, M.; Sugioka, K.; Nakano, M. A ferriprotoporphyrin IX-chloroquine complex promotes membrane phospholipid peroxidation A possible mechanism for antimalarial action. FEBS Lett. 1987, 223, 251–254. [Google Scholar] [CrossRef] [Green Version]
- Haynes, R.K.; Cheu, K.W.; Chan, H.W.; Wong, H.N.; Li, K.Y.; Tang, M.M.K.; Chen, M.J.; Guo, Z.F.; Guo, Z.H.; Sinniah, K.; et al. Interactions between Artemisinins and other Antimalarial Drugs in Relation to the Cofactor Model-A Unifying Proposal for Drug Action. ChemMedChem 2012, 7, 2204–2226. [Google Scholar] [CrossRef]
- De Almeida Ribeiro, M.C.; Augusto, O.; Da Costa Ferreira, A.M. Inhibitory effect of chloroquine on the peroxidase activity of ferriprotoporphyrin IX. J. Chem. Soc. Dalt. Trans. 1995, 3759–3766. [Google Scholar] [CrossRef]
- Birth, D.; Kao, W.C.; Hunte, C. Structural analysis of atovaquone-inhibited cytochrome bc 1 complex reveals the molecular basis of antimalarial drug action. Nat. Commun. 2014, 5, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Fry, M.; Pudney, M. Site of action of the antimalarial hydroxynaphthoquinone, 2-[trans-4-(4’-chlorophenyl) cyclohexyl]-3- hydroxy-1,4-naphthoquinone (566C80). Biochem. Pharmacol. 1992, 43, 1545–1553. [Google Scholar] [CrossRef]
- Srivastava, I.K.; Rottenberg, H.; Vaidya, A.B. Atovaquone, a broad spectrum antiparasitic drug, collapses mitochondrial membrane potential in a malarial parasite. J. Biol. Chem. 1997, 272, 3961–3966. [Google Scholar] [CrossRef] [Green Version]
- Barton, V.; Fisher, N.; Biagini, G.A.; Ward, S.A.; O’Neill, P.M. Inhibiting Plasmodium cytochrome bc1: A complex issue. Curr. Opin. Chem. Biol. 2010, 14, 440–446. [Google Scholar] [CrossRef]
- Fiorillo, M.; Lamb, R.; Tanowitz, H.B.; Mutti, L.; Krstic-Demonacos, M.; Cappello, A.R.; Martinez-Outschoorn, U.E.; Sotgia, F.; Lisanti, M.P. Repurposing atovaquone: Targeting mitochondrial complex III and OXPHOS to eradicate cancer stem cells. Oncotarget 2016, 7, 34084–34099. [Google Scholar] [CrossRef] [Green Version]
- Kevin Park, B.; O’Neill, P.M.; Maggs, J.L.; Pirmohamed, M. Safety assessment of peroxide antimalarials: Clinical and chemical perspectives. Br. J. Clin. Pharmacol. 1998, 46, 521–529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nosten, F.; White, N.J. Artemisinin-based combination treatment of falciparum malaria. Am. J. Trop. Med. Hyg. 2007, 77, 181–192. [Google Scholar] [CrossRef] [PubMed]
- Rudrapal, M.; Chetia, D. Endoperoxide antimalarials: Development, structural diversity and pharmacodynamic aspects with reference to 1,2,4-trioxane-based structural scaffold. Drug Des. Devel. Ther. 2016, 10, 3575–3590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Posner, G.H.; Oh, C.H. A Regiospecifically Oxygen-18 Labeled 1,2,4-Trioxane: A Simple Chemical Model System To Probe the Mechanism(s) for the Antimalarial Activity of Artemisinin (Qinghaosu). J. Am. Chem. Soc. 1992, 114, 8328–8329. [Google Scholar] [CrossRef]
- Mercer, A.E.; Copple, I.M.; Maggs, J.L.; O’Neill, P.M.; Park, B.K. The role of heme and the mitochondrion in the chemical and molecular mechanisms of mammalian cell death induced by the artemisinin antimalarials. J. Biol. Chem. 2011, 286, 987–996. [Google Scholar] [CrossRef] [Green Version]
- Posner, G.H.; Oh, C.H.; Wang, D.; Gerena, L.; Milhous, W.K.; Meshnick, S.R.; Asawamahasadka, W. Mechanism-Based Design, Synthesis, and in Vitro Antimalarial Testing of New 4-Methylated Trioxanes Structurally Related to Artemisinin: The Importance of a Carbon-Centered Radical for Antimalarial Activity. J. Med. Chem. 1994, 37, 1256–1258. [Google Scholar] [CrossRef]
- Robert, A.; Benoit-Vical, F.; Claparols, C.; Meunier, B. The antimalarial drug artemisinin alkylates heme in infected mice. Proc. Natl. Acad. Sci. USA 2005, 102, 13676–13680. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Gerhard, G.S. Heme activates artemisinin more efficiently than hemin, inorganic iron, or hemoglobin. Bioorg. Med. Chem. 2008, 16, 7853–7861. [Google Scholar] [CrossRef]
- Xie, S.C.; Dogovski, C.; Hanssen, E.; Chiu, F.; Yang, T.; Crespo, M.P.; Stafford, C.; Batinovic, S.; Teguh, S.; Charman, S.; et al. Haemoglobin degradation underpins the sensitivity of early ring stage Plasmodium falciparum to artemisinins. J. Cell Sci. 2016, 129, 406–416. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Huang, L.; Li, J.; Fan, Q.; Long, Y.; Li, Y.; Zhou, B. Artemisinin directly targets malarial mitochondria through its specific mitochondrial activation. PLoS ONE 2010, 5, e9582. [Google Scholar] [CrossRef] [Green Version]
- Meunier, B.; de Visser, S.P.; Shaik, S. Mechanism of oxidation reactions catalyzed by cytochrome P450 enzymes. Chem. Rev. 2004, 104, 3947–3980. [Google Scholar] [CrossRef]
- Dawson, J.H. Probing structure-function relations in heme-containing oxygenases and peroxidases. Science 1988, 240, 433–439. [Google Scholar] [CrossRef]
- Fita, I.; Rossmann, M.G. The active center of catalase. J. Mol. Biol. 1985, 185, 21–37. [Google Scholar] [CrossRef]
- Edwards, S.L.; Kraut, J.; Poulos, T.L. Crystal Structure of Nitric Oxide Inhibited Cytochrome c Peroxidase. Biochemistry 1988, 27, 8074–8081. [Google Scholar] [CrossRef]
- Allen, R.J.W.; Kirk, K. The Membrane Potential of the Intraerythrocytic Malaria Parasite Plasmodium falciparum. J. Biol. Chem. 2004, 279, 11264–11272. [Google Scholar] [CrossRef] [Green Version]
- Biagini, G.A.; Viriyavejakul, P.; O’Neill, P.M.; Bray, P.G.; Ward, S.A. Functional characterization and target validation of alternative complex I of Plasmodium falciparum mitochondria. Antimicrob. Agents Chemother. 2006, 50, 1841–1851. [Google Scholar] [CrossRef] [Green Version]
- Antoine, T.; Fisher, N.; Amewu, R.; O’Neill, P.M.; Ward, S.A.; Biagini, G.A. Rapid kill of malaria parasites by artemisinin and semi-synthetic endoperoxides involves ROS-dependent depolarization of the membrane potential. J. Antimicrob. Chemother. 2014, 69, 1005–1016. [Google Scholar] [CrossRef]
- WHO. Artemisinin Resistance and Artemisinin-Based Combination Therapy Efficacy (Status Report—August 2018). Available online: https://apps.who.int/iris/bitstream/handle/10665/274362/WHO-CDS-GMP-2018.18-eng.pdf?ua=1 (accessed on 26 December 2020).
- Van Vugt, M.; Brockman, A.; Gemperli, B.; Luxemburger, C.; Gathmann, I.; Royce, C.; Slight, T.; Looareesuwan, S.; White, N.J.; Nosten, F. Randomized comparison of artemether-benflumetol and artesunate-mefloquine in treatment of multidrug-resistant falciparum malaria. Antimicrob. Agents Chemother. 1998, 42, 135–139. [Google Scholar] [CrossRef] [Green Version]
- Benoit-Vical, F.; Lelièvre, J.; Berry, A.; Deymier, C.; Dechy-Cabaret, O.; Cazelles, J.; Loup, C.; Robert, A.; Magnaval, J.F.; Meunier, B. Trioxaquines are new antimalarial agents active on all erythrocytic forms, including gametocytes. Antimicrob. Agents Chemother. 2007, 51, 1463–1472. [Google Scholar] [CrossRef] [Green Version]
- Leang, R.; Barrette, A.; Bouth, D.M.; Menard, D.; Abdur, R.; Duong, S.; Ringwald, P. Efficacy of dihydroartemisinin-piperaquine for treatment of uncomplicated Plasmodium falciparum and Plasmodium vivax in Cambodia, 2008 to 2010. Antimicrob. Agents Chemother. 2013, 57, 818–826. [Google Scholar] [CrossRef] [Green Version]
- Bopp, S.; Magistrado, P.; Wong, W.; Schaffner, S.F.; Mukherjee, A.; Lim, P.; Dhorda, M.; Amaratunga, C.; Woodrow, C.J.; Ashley, E.A.; et al. Plasmepsin II-III copy number accounts for bimodal piperaquine resistance among Cambodian Plasmodium falciparum. Nat. Commun. 2018, 9, 1769. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toler, S. The plasmodial apicoplast was retained under evolutionary selective pressure to assuage blood stage oxidative stress. Med. Hypotheses 2005, 65, 683–690. [Google Scholar] [CrossRef] [PubMed]
- Storm, J. Lipoic Acid Metabolism of Plasmodium—A Suitable Drug Target. Curr. Pharm. Des. 2012, 18, 3480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Birnbaum, J.; Scharf, S.; Schmidt, S.; Jonscher, E.; Maria Hoeijmakers, W.A.; Flemming, S.; Toenhake, C.G.; Schmitt, M.; Sabitzki, R.; Bergmann, B.; et al. A Kelch13-defined endocytosis pathway mediates artemisinin resistance in malaria parasites. Science 2020, 367, 51–59. [Google Scholar] [CrossRef]
- Klonis, N.; Crespo-Ortiz, M.P.; Bottova, I.; Abu-Bakar, N.; Kenny, S.; Rosenthal, P.J.; Tilley, L. Artemisinin activity against Plasmodium falciparum requires hemoglobin uptake and digestion. Proc. Natl. Acad. Sci. USA 2011, 108, 11405–11410. [Google Scholar] [CrossRef] [Green Version]
- Ariey, F.; Witkowski, B.; Amaratunga, C.; Beghain, J.; Langlois, A.C.; Khim, N.; Kim, S.; Duru, V.; Bouchier, C.; Ma, L.; et al. A molecular marker of artemisinin-resistant Plasmodium falciparum malaria. Nature 2014, 505, 50–55. [Google Scholar] [CrossRef]
- Witkowski, B.; Lelièvre, J.; Barragán, M.J.L.; Laurent, V.; Su, X.Z.; Berry, A.; Benoit-Vical, F. Increased tolerance to artemisinin in Plasmodium falciparum is mediated by a quiescence mechanism. Antimicrob. Agents Chemother. 2010, 54, 1872–1877. [Google Scholar] [CrossRef] [Green Version]
- Witkowski, B.; Khim, N.; Chim, P.; Kim, S.; Ke, S.; Kloeung, N.; Chy, S.; Duong, S.; Leang, R.; Ringwald, P.; et al. Reduced artemisinin susceptibility of Plasmodium falciparum ring stages in western cambodia. Antimicrob. Agents Chemother. 2013, 57, 914–923. [Google Scholar] [CrossRef] [Green Version]
- Peatey, C.; Chen, N.; Gresty, K.; Anderson, K.; Pickering, P.; Watts, R.; Gatton, M.L.; McCarthy, J.; Cheng, Q. Dormant Plasmodium falciparum Parasites in Human Infections Following Artesunate Therapy. J. Infect. Dis. 2021, 223, 1631–1638. [Google Scholar] [CrossRef]
- Clarebout, G.; Slomianny, C.; Delcourt, P.; Leu, B.; Masset, A.; Camus, D.; Dive, D. Status of Plasmodium falciparum towards catalase. Br. J. Haematol. 1998, 103, 52–59. [Google Scholar] [CrossRef]
- Varadharajan, S.; Sagar, B.K.C.; Rangarajan, P.N.; Padmanaban, G. Localization of ferrochelatase in Plasmodium falciparum. Biochem. J. 2004, 384, 429–436. [Google Scholar] [CrossRef] [Green Version]
- Bonday, Z.Q.; Dhanasekaran, S.; Rangarajan, P.N.; Padmanaban, G. Import of host δ-aminolevulinate dehydratase into the malarial parasite: Identification of a new drug target. Nat. Med. 2000, 6, 898–903. [Google Scholar] [CrossRef]
- Rocamora, F.; Zhu, L.; Liong, K.Y.; Dondorp, A.; Miotto, O.; Mok, S.; Bozdech, Z. Oxidative stress and protein damage responses mediate artemisinin resistance in malaria parasites. PLoS Pathog. 2018, 14, e1006930. [Google Scholar] [CrossRef]
- Mok, S.; Ashley, E.A.; Ferreira, P.E.; Zhu, L.; Lin, Z.; Yeo, T.; Chotivanich, K.; Imwong, M.; Pukrittayakamee, S.; Dhorda, M.; et al. Population transcriptomics of human malaria parasites reveals the mechanism of artemisinin resistance. Science 2015, 347, 431–435. [Google Scholar] [CrossRef] [Green Version]
- Mbengue, A.; Bhattacharjee, S.; Pandharkar, T.; Liu, H.; Estiu, G.; Stahelin, R.V.; Rizk, S.S.; Njimoh, D.L.; Ryan, Y.; Chotivanich, K.; et al. A molecular mechanism of artemisinin resistance in Plasmodium falciparum malaria. Nature 2015, 520, 683–687. [Google Scholar] [CrossRef] [Green Version]
- Tawk, L.; Chicanne, G.; Dubremetz, J.F.; Richard, V.; Payrastre, B.; Vial, H.J.; Roy, C.; Wengelnik, K. Phosphatidylinositol 3-phosphate, an essential lipid in Plasmodium, localizes to the food vacuole membrane and the apicoplast. Eukaryot. Cell 2010, 9, 1519–1530. [Google Scholar] [CrossRef] [Green Version]
- Isozumi, R.; Uemura, H.; Kimata, I.; Ichinose, Y.; Logedi, J.; Omar, A.H.; Kaneko, A. Novel mutations in k13 propeller gene of artemisinin-resistant Plasmodium falciparum. Emerg. Infect. Dis. 2015, 21, 490–492. [Google Scholar] [CrossRef] [Green Version]
- Balikagala, B.; Mita, T.; Ikeda, M.; Sakurai, M.; Yatsushiro, S.; Takahashi, N.; Tachibana, S.I.; Auma, M.; Ntege, E.H.; Ito, D.; et al. Absence of in vivo selection for K13 mutations after artemether-lumefantrine treatment in Uganda. Malar. J. 2017, 16, 23. [Google Scholar] [CrossRef] [Green Version]
- Lee, A.H.; Fidock, D.A. Evidence of a mild mutator phenotype in cambodian Plasmodium falciparum malaria parasites. PLoS ONE 2016, 11, e0154166. [Google Scholar] [CrossRef] [Green Version]
- Paloque, L.; Witkowski, B.; Lelièvre, J.; Ouji, M.; Ben Haddou, T.; Ariey, F.; Robert, A.; Augereau, J.M.; Ménard, D.; Meunier, B.; et al. Endoperoxide-based compounds: Cross-resistance with artemisinins and selection of a Plasmodium falciparum lineage with a K13 non-synonymous polymorphism. J. Antimicrob. Chemother. 2018, 73, 395–403. [Google Scholar] [CrossRef] [Green Version]
- Schirmer, R.H.; Coulibaly, B.; Stich, A.; Scheiwein, M.; Merkle, H.; Eubel, J.; Becker, K.; Becher, H.; Müller, O.; Zich, T.; et al. Methylene blue as an antimalarial agent. Redox Rep. 2003, 8, 272–275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McNamara, C.W.; Lee, M.C.S.; Lim, C.S.; Lim, S.H.; Roland, J.; Nagle, A.; Simon, O.; Yeung, B.K.S.; Chatterjee, A.K.; McCormack, S.L.; et al. Targeting Plasmodium PI(4)K to eliminate malaria. Nature 2013, 504, 248–253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tiwari, N.K.; Reynolds, P.J.; Calderón, A.I. Preliminary LC-MS Based Screening for Inhibitors of Plasmodium falciparum Thioredoxin Reductase (PfTrxR) among a Set of Antimalarials from the Malaria Box. Molecules 2016, 21, 424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vallières, C.; Avery, S.V. The candidate antimalarial drug mmv665909 causes oxygen-dependent mRNA mistranslation and synergizes with quinoline-derived antimalarials. Antimicrob. Agents Chemother. 2017, 61, e00459-17. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Agent | Site of Production | Role in Oxidative Homeostasis |
|---|---|---|
| HRP | FV | Binding with heme for polymerization |
| HDP | FV | Heme polymerization to hemozoin |
| H2O2 | Cytosol and FV | Degrades heme |
| SOD | Cytosol, Mitochondria | Dismutation of O2•− |
| Prx | Cytosol, Mitochondria, Apicoplast | Reduction of H2O2 to H2O |
| Trx | Cytosol, Mitochondria | Reduction of Prx |
| GSH | Cytosol | Degradation of heme, reduction of proteins and ROS |
| Vit B6 | Cytosol | Role unclear |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Egwu, C.O.; Augereau, J.-M.; Reybier, K.; Benoit-Vical, F. Reactive Oxygen Species as the Brainbox in Malaria Treatment. Antioxidants 2021, 10, 1872. https://doi.org/10.3390/antiox10121872
Egwu CO, Augereau J-M, Reybier K, Benoit-Vical F. Reactive Oxygen Species as the Brainbox in Malaria Treatment. Antioxidants. 2021; 10(12):1872. https://doi.org/10.3390/antiox10121872
Chicago/Turabian StyleEgwu, Chinedu Ogbonnia, Jean-Michel Augereau, Karine Reybier, and Françoise Benoit-Vical. 2021. "Reactive Oxygen Species as the Brainbox in Malaria Treatment" Antioxidants 10, no. 12: 1872. https://doi.org/10.3390/antiox10121872