
Antioxidative Responses of Duckweed (Lemna minor L.) to Phenol and Rhizosphere-Associated Bacterial Strain Hafnia paralvei C32-106/3


 ,
,  and
and 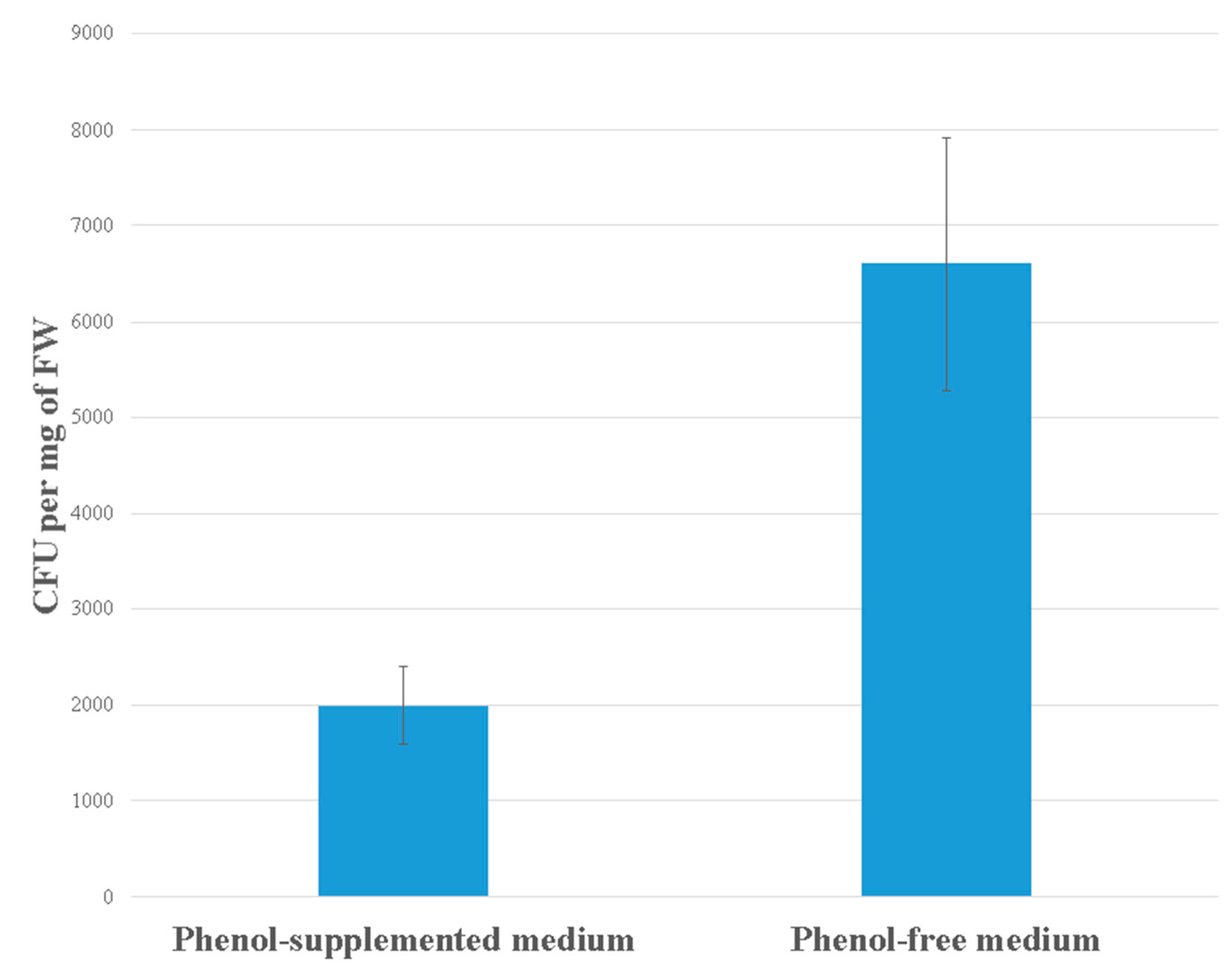{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Growth Conditions
2.2. Experiments with Phenol
2.3. Bacterial Strain Hafnia Paralvei Strain C32-106/3 (Accession Number MF526939)
2.4. Estimation of H. paralvei Density on the Surface of L. minor
2.5. Antioxidative Enzymes
2.5.1. Extraction of Total Proteins
2.5.2. Isoelectric Focusing and Zymogram Detection of Guaiacol Peroxidase (GPX, EC 1.11.1.7)
2.6. Determination of Hydrogen Peroxide
2.7. Lipid Peroxidation (MDA Content)
2.8. Determination of Ascorbic Acid
2.9. Analysis of IAA Production
2.10. Statistical Analysis
3. Results
3.1. Bacterial Density on the Duckweeds’ Surface
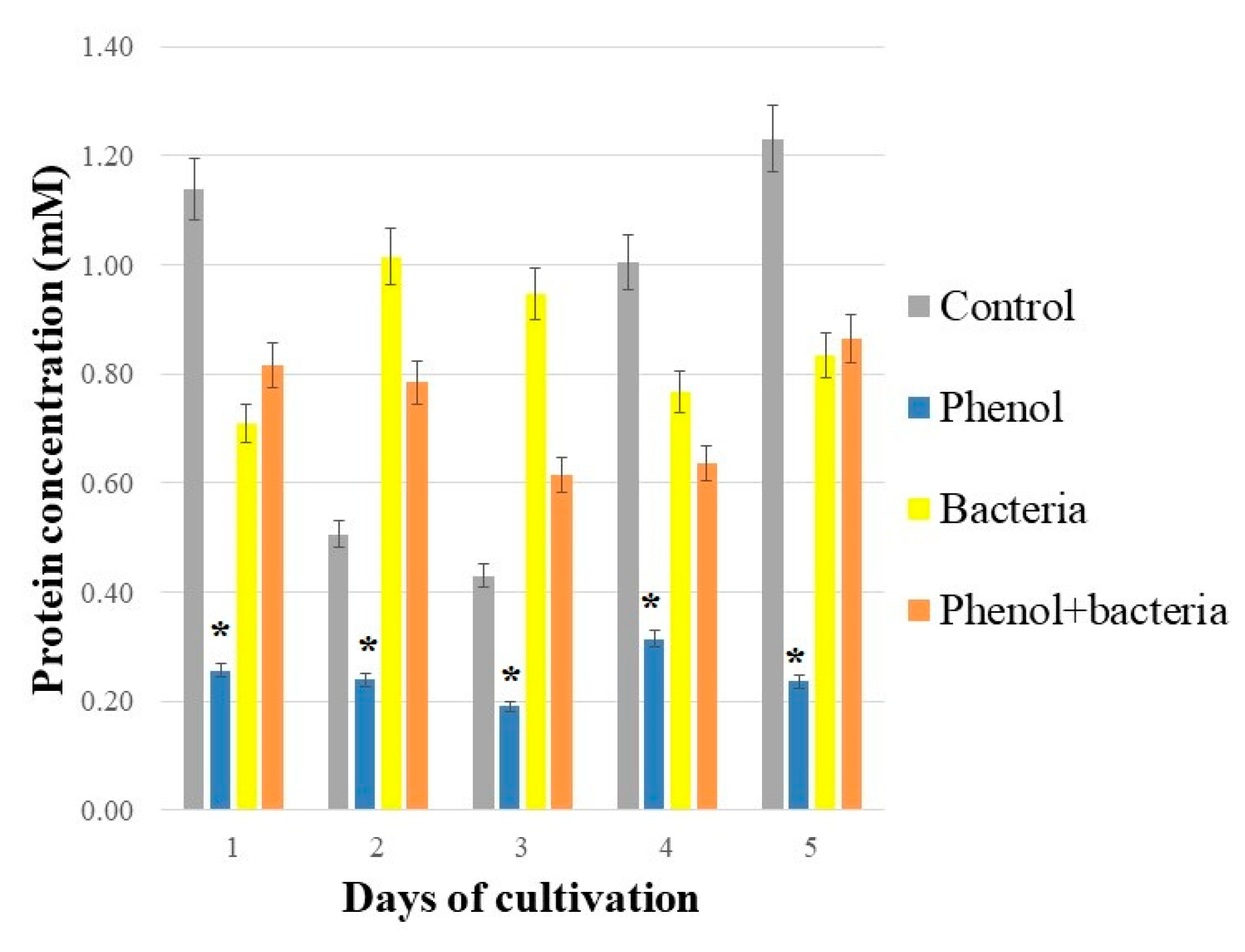
3.2. Total Soluble Proteins
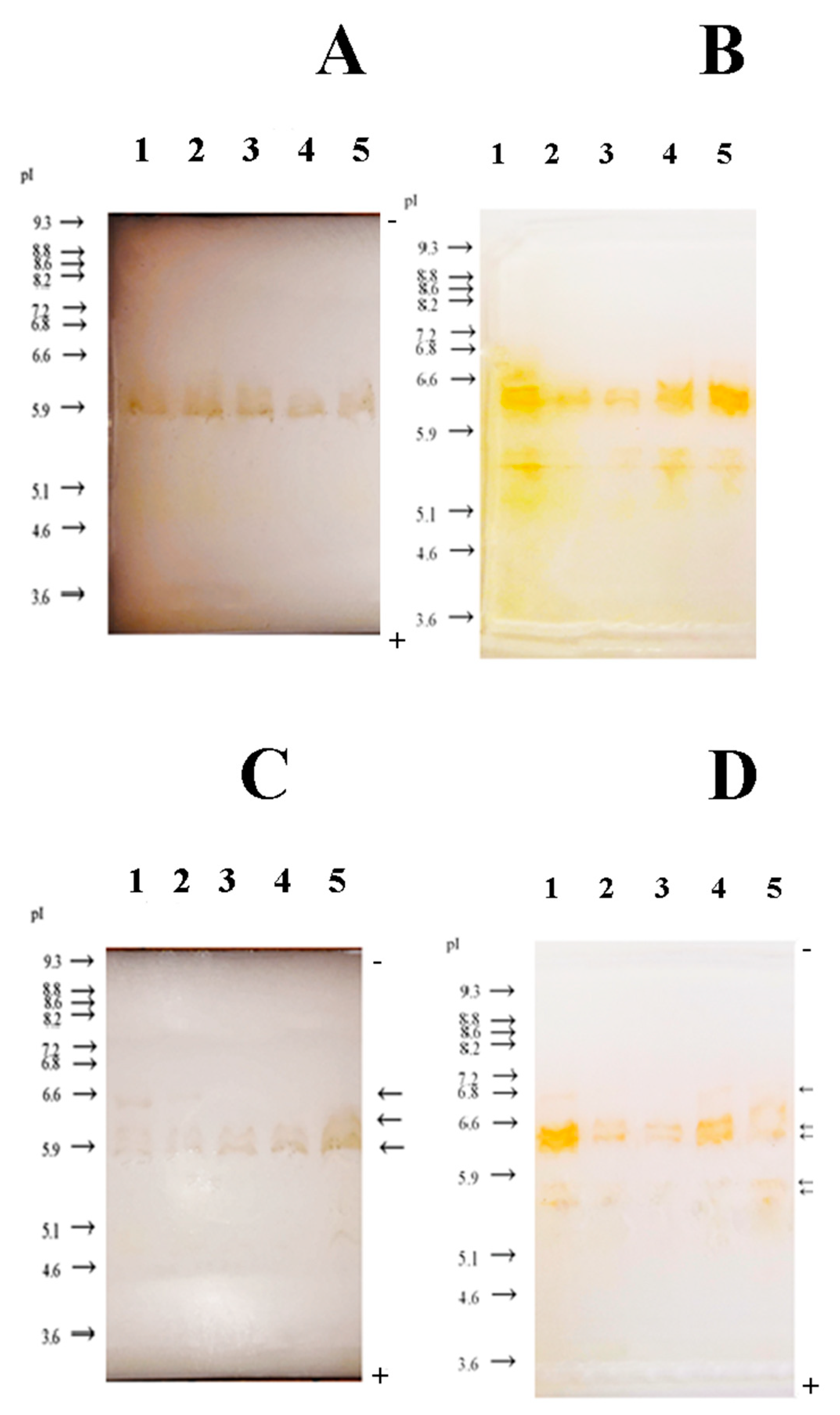
3.3. Zymogram Detection of GPX
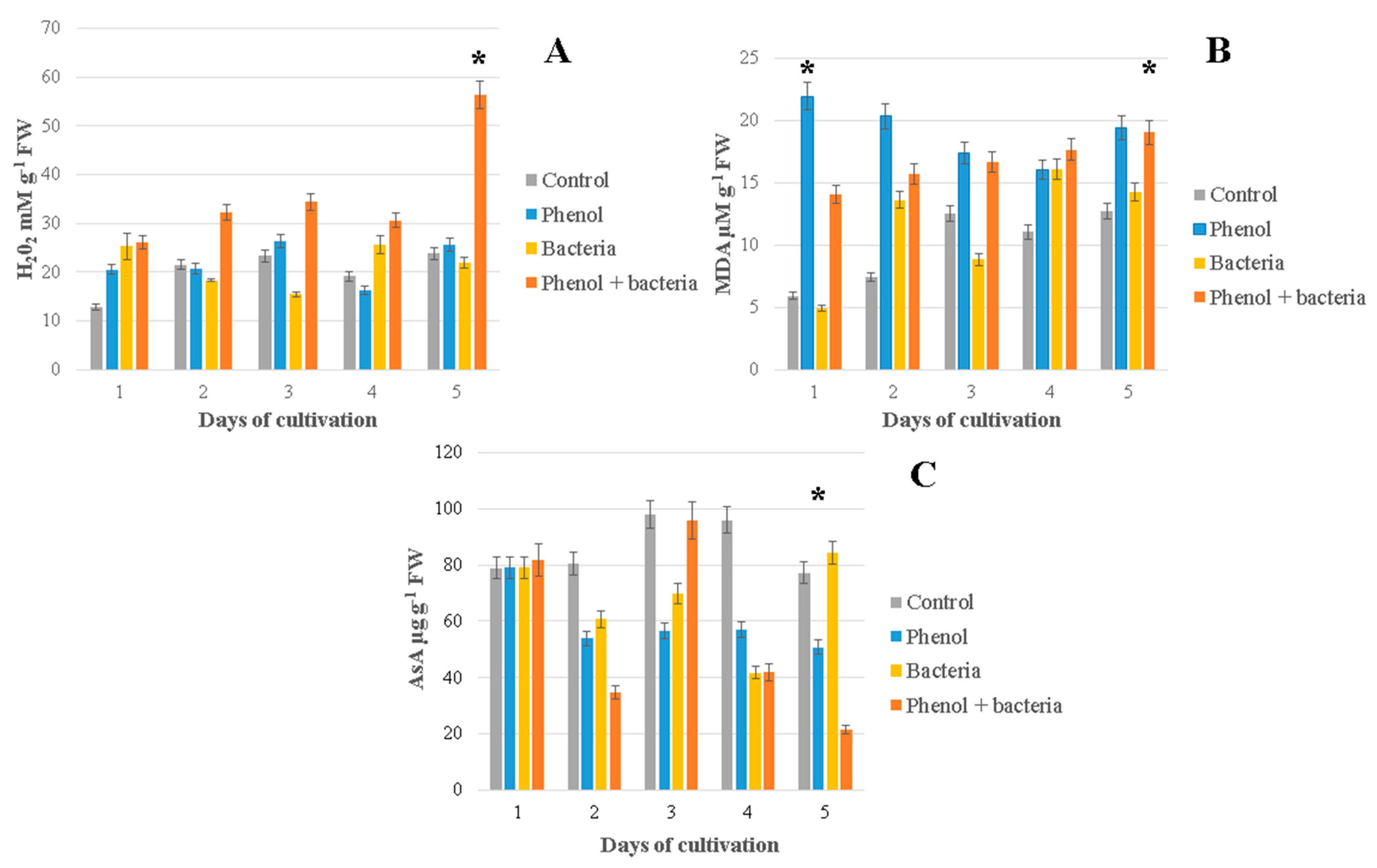
3.4. Hydrogen Peroxide (H2O2)
3.5. Lipid Peroxidation (MDA Content)
3.6. Ascorbic Acid
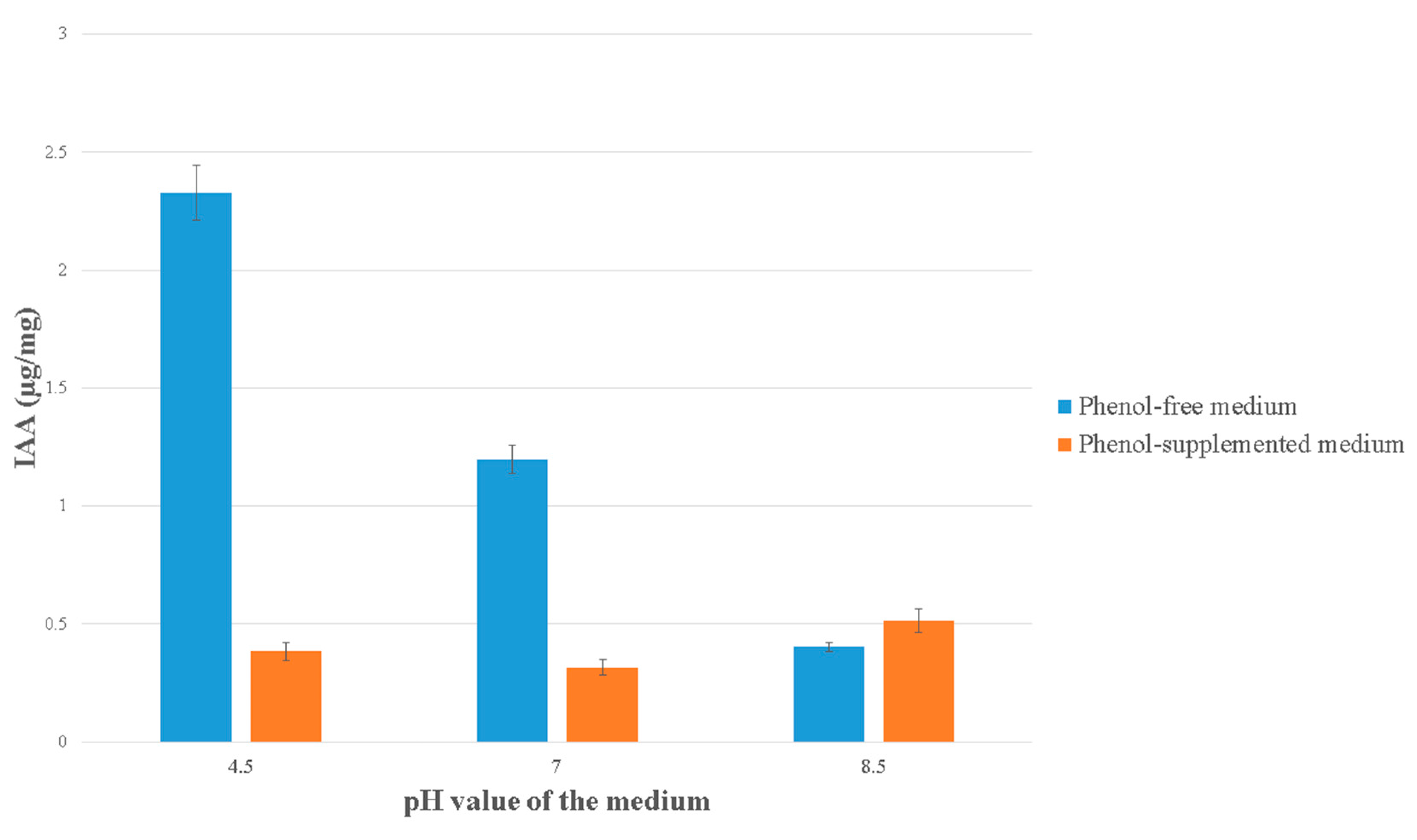
3.7. IAA Production
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gopalapillai, Y.; Vigneault, B.; Hale, B. Root Length of Aquatic Plant, Lemna minor L., as an Optimal Toxicity Endpoint for Biomonitoring of Mining Effluents. Integr. Environ. Assess. Manag. 2014, 10, 493–497. [Google Scholar] [CrossRef]
- Ziegler, P.; Sree, K.S.; Appenroth, K.J. Duckweeds for water remediation and toxicity testing. Toxicol. Environ. Chem. 2016, 98, 1127–1154. [Google Scholar] [CrossRef]
- Basiglini, E.; Pintore, M.; Forni, C. Effects of treated industrial wastewaters and temperatures on growth and enzymatic activities of duckweed (Lemna minor L.). Ecotoxicol. Environ. Saf. 2018, 153, 54–59. [Google Scholar] [CrossRef] [PubMed]
- Iatrou, E.I.; Kora, E.; Stasinakis, A.S. Investigation of biomass production, crude protein and starch content in laboratory wastewater treatment systems planted with Lemna minor and Lemna gibba. Environ. Technol. 2019, 40, 2649–2656. [Google Scholar] [CrossRef] [PubMed]
- Duan, W.; Meng, F.; Cui, H.; Lin, Y.; Wang, G.; Wu, J. Ecotoxicity of phenol and cresols to aquatic organisms: A review. Ecotoxicol. Environ. Saf. 2018, 157, 441–456. [Google Scholar] [CrossRef]
- Czarnocka, W.; Karpiński, S. Friend or foe? Reactive oxygen species production, scavenging and signaling in plant response to environmental stresses. Free Radic. Biol. Med. 2018, 122, 4–20. [Google Scholar] [CrossRef]
- Chaki, M.; Begara-Morales, J.C.; Barroso, J.B. Oxidative Stress in Plants. Antioxidants 2020, 9, 481. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Bhuyan, M.H.M.B.; Zulfiqar, F.; Raza, A.; Mohsin, S.M.; Mahmud, J.A.; Fujita, M.; Fotopoulos, V. Reactive Oxygen Species and Antioxidant Defense in Plants under Abiotic Stress: Revisiting the Crucial Role of a Universal Defense Regulator. Antioxidants 2020, 9, 681. [Google Scholar] [CrossRef]
- Panda, S.K. Impact of copper on reactive oxygen species, lipid peroxidation and antioxidants in Lemna minor. Biol. Plant. 2008, 52, 561–564. [Google Scholar] [CrossRef]
- Obermeier, M.; Schröder, C.A.; Helmreich, B.; Schröder, P. The enzymatic and antioxidative stress response of Lemna minor to copper and a chloroacetamide herbicide. Environ. Sci. Pollut. Res. 2015, 22, 18495–18507. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Gu, W.; Dai, Z.; Li, J.; Jiang, H.; Zhang, Q. Boron accumulation by Lemna minor L. under salt stress. Sci. Rep. 2018, 8, 8954–8960. [Google Scholar] [CrossRef] [Green Version]
- Singh, D.P.; Singh, V.; Gupta, V.K.; Shukla, R.; Prabha, R.; Sarma, B.K.; Patel, J.S. Microbial inoculation in rice regulates antioxidative reactions and defense related genes to mitigate drought stress. Sci. Rep. 2020, 10, 4818–4835. [Google Scholar] [CrossRef]
- Glick, B. Plant Growth-Promoting Bacteria: Mechanisms and Applications. Scientifica 2012, 2012, 963401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Backer, R.; Rokem, J.S.; Ilangumaran, G.; Lamont, J.; Praslickova, D.; Ricci, E.; Subramanian, S.; Smith, D.L. Plant Growth-Promoting Rhizobacteria: Context, Mechanisms of Action, and Roadmap to Commercialization of Biostimulants for Sustainable Agriculture. Front. Plant. Sci. 2018, 9, 1473–1490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishizawa, H.; Kuroda, M.; Morikawa, M.; Ike, M. Differential oxidative and antioxidative response of duckweed Lemna minor toward plant growth promoting/inhibiting bacteria. Plant Physiol. Biochem. 2017, 118, 667–673. [Google Scholar] [CrossRef]
- Ishizawa, H.; Tada, M.; Kuroda, M.; Inoue, D.; Ike, M. Performance of plant growth-promoting bacterium of duckweed under different kinds of abiotic stress factors. Biocatal. Agric. Biotechnol. 2019, 19, 101–146. [Google Scholar] [CrossRef]
- Radulovic, O.; Petrić, M.; Raspor, M.; Tadić, V.; Jovanović, P.; Zečević, V. Assessment of in vitro Multiplication of Lemna minor in the Presence of Phenol: Plant/Bacteria System for Potential Bioremediation—Part I. Pol. J. Environ. Stud. 2019, 28, 803–809. [Google Scholar] [CrossRef]
- Radulovic, O.; Petrić, M.; Raspor, M.; Stanojević, O.; Janakiev, T.; Tadić, V.; Stanković, S. Culture-Dependent Analysis of 16S rRNA Sequences Associated with the Rhizosphere of Lemna minor and Assessment of Bacterial Phenol-Resistance: Plant/Bacteria System for Potential Bioremediation—Part II. Pol. J. Environ. Stud. 2019, 28, 811–822. [Google Scholar] [CrossRef] [Green Version]
- Radulović, O.; Stanković, S.; Uzelac, B.; Tadić, V.; Trifunović-Momčilov, M.; Lozo, J.; Marković, M. Phenol Removal Capacity of the Common Duckweed (Lemna minor L.) and Six Phenol-Resistant Bacterial Strains from Its Rhizosphere: In Vitro Evaluation at High Phenol Concentrations. Plants 2020, 9, 599. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays with tobacco culture. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation pf microgram quantities of protein utilizing the principle of protein binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Vujčić, Z.; Janović, B.; Lončar, N.; Margetić, A.; Božić, N.; Dojnov, B.; Vujčić, M. Exploitation of neglected horseradish peroxidase izoenzymes for dye decolorization. Int. Biodeterior. Biodegrad. 2015, 97, 124–127. [Google Scholar] [CrossRef]
- Siegel, B.Z.; Galston, A.W. The isoperoxidases of Pisum sativum. Plant Physiol. 1976, 42, 221–226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sergiev, I.; Alexieva, V.; Karanov, E. Effect of spermine, atrazine and combination between them on some endogenous protective systems and stress markers in plants. Compt. Rend. Acad. Bulg. Sci. 1997, 51, 121–124. [Google Scholar]
- Velikova, V.; Yordanov, I.; Edreva, A. Oxidative stress and some antioxidant systems in acid rain-treated bean plants: Protective role of exogenous polyamines. Plant Sci. 2000, 151, 59–66. [Google Scholar] [CrossRef]
- Mukherjee, S.P.; Choudhuri, M.A. Implications of water stress-induced changes in the levels of endogenous ascorbic acid and hydrogen peroxide in Vigna seedlings. Physiol. Plant. 1983, 58, 166–170. [Google Scholar] [CrossRef]
- Gordon, S.A.; Weber, R.P. Colorimetric Estimation of Indoleacetic Acid. Plant Physiol. 1951, 26, 192–195. [Google Scholar] [CrossRef] [Green Version]
- Toyama, T.; Sei, K.; Yu, N.; Kumada, H.; Inoue, D.; Hoang, H.; Soda, S.; Chang, Y.C.; Kikuchi, S.; Fujita, M.; et al. Enrichment of bacteria possessing catechol dioxygenase genes in the rhizosphere of Spirodela polyrrhiza: A mechanism of accelerated biodegradation of phenol. Water Res. 2009, 43, 3765–3776. [Google Scholar] [CrossRef]
- Yamaga, F.; Washio, K.; Morikawa, M. Sustainable biodegradation of phenol by Acinetobacter calcoaceticus P23 isolated from the rhizosphere of duckweed Lemna aoukikusa. Environ. Sci. Technol. 2010, 44, 6470–6474. [Google Scholar] [CrossRef]
- Rucká, L.; Nešvera, J.; Pátek, M. Biodegradation of phenol and its derivatives by engineered bacteria: Current knowledge and perspectives. World J. Microbiol. Biotechnol. 2017, 33, 174–226. [Google Scholar] [CrossRef]
- Shigeto, J.; Tsutsumi, Y. Diverse functions and reactions of class III peroxidases. New Phytol. 2016, 209, 1395–1402. [Google Scholar] [CrossRef] [Green Version]
- Passardi, F.; Cosio, C.; Penel, C.; Dunand, C. Peroxidases have more functions than a Swiss army knife. Plant Cell Rep. 2005, 24, 255–265. [Google Scholar] [CrossRef] [PubMed]
- Chang, I.H.; Cheng, K.T.; Huang, P.C.; Lin, Y.Y.; Cheng, L.J.; Cheng, T.S. Oxidative stress in greater duckweed (Spirodela polyrhiza) caused by long-term NaCl exposure. Acta Physiol. Plant. 2012, 34, 1165–1176. [Google Scholar] [CrossRef]
- Ibáñez, S.G.; Sosa Alderete, L.G.; Medina, M.I.; Agostini, E. Phytoremediation of phenol using Vicia sativa L. plants and its antioxidative response. Environ. Sci. Pollut. Res. 2012, 19, 1555–1562. [Google Scholar] [CrossRef] [PubMed]
- Razinger, J.; Dermastia, M.; Drinovec, L.; Drobne, D.; Zrimec, A.; Koce, J.D. Antioxidative Responses of Duckweed (Lemna minor L.) to Short-Term Copper Exposure. Environ. Sci. Pollut. Res. 2007, 14, 194–201. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.; Zhang, Y.; Cui, Y.; Ma, J. Effects of a rhizobacterium on the growth of and chromium remediation by Lemna minor. Environ. Sci. Pollut. Res. 2015, 22, 9686–9693. [Google Scholar] [CrossRef]
- Habib, S.H.; Kausar, H.; Saud, H.M. Plant growth-promoting 456 rhizobacteria enhance salinity stress tolerance in okra through ROS-scavenging enzyme. BioMed Res. Int. 2016, 2016, 6284547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kahoush, M.; Nemeshwaree, B.; Aurélie, C.; Vincent, N. Bio-Fenton and Bio-electro-Fenton as sustainable methods for degrading organic pollutants in wastewater. Process. Biochem. 2017, 64, 237–247. [Google Scholar] [CrossRef]
- White, J.; Torres, M.; Somu, M.; Johnson, H.; Irizarry, I.; Chen, Q.; Zhang, N.; Walsh, E.; Tadych, M. Hydrogen Peroxide Staining to Visualize Intracellular Bacterial Infections of Seedling Root Cells. Microsc. Res. Tech. 2014, 77, 566–573. [Google Scholar] [CrossRef]
- Nanda, A.K.; Andrio, E.; Marino, D.; Pauly, N.; Dunand, C. Reactive Oxygen Species during Plant-microorganism Early Interactions. J. Integr. Plant Biol. 2010, 52, 195–204. [Google Scholar] [CrossRef]
- Hyodo, K.; Suzuki, N.; Mise, K.; Okuno, T. Roles of superoxide anion and hydrogen peroxide during replication of two unrelated plant RNA viruses in Nicotiana benthamiana. Plant Signal. Behav. 2017, 12, e1338223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barber, J.T.; Sharma, H.A.; Ensley, H.E.; Polito, M.A.; Thomas, D.A. Detoxification of phenol by the aquatic angiosperm, Lemna gibba. Chemosphere 1995, 31, 3567–3574. [Google Scholar] [CrossRef]
- Appenroth, K.J.; Sree, K.S.; Fakhoorian, T.; Lam, E. Resurgence of duckweed research and applications: Report from the 3rd International Duckweed Conference. Plant Mol. Biol. 2015, 89, 647–654. [Google Scholar] [CrossRef] [PubMed]
- Almagro, L.; Gómez Ros, L.V.; Belchi-Navarro, S.; Bru, R.; Ros Barceló, A.; Pedreño, M.A. Class III peroxidases in plant defense reactions. J. Exp. Bot. 2009, 60, 377–390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Li, X.; Zhou, L.; Wang, L.; Zhou, Q.; Huang, X. Analysis of effects of a new environmental pollutant, bisphenol A, on antioxidant systems in soybean roots at different growth stages. Sci. Rep. 2016, 31, 23782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smirnoff, N.; Wheeler, G.L. Ascorbic acid in plants: Biosynthesis and function. Crit. Rev. Biochem. Mol. Biol. 2000, 35, 291–314. [Google Scholar] [CrossRef]
- Trebst, A. Inhibition of photosynthetic electron flow by phenol and diphenylether herbicides in control and trypsin-treated chloroplasts. Z. Fur Nat. C 1979, 34, 986–991. [Google Scholar] [CrossRef]
- Chandra, S.; Askari, K.; Kumari, M. Optimization of indole acetic acid production by isolated bacteria from Stevia rebaudiana rhizosphere and its effects on plant growth. J. Genet. Eng. Biotechnol. 2018, 16, 581–586. [Google Scholar] [CrossRef] [PubMed]
- Idris, E.E.; Iglesias, D.J.; Talon, M.; Borriss, R. Tryptophan-dependent production of indole-3-acetic acid (IAA) affects level of plant growth promotion by Bacillus amyloliquefaciens FZB42. Mol. Plant-Microbe Interactions 2007, 20, 619–626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Utami, D.; Kawahata, A.; Sugawara, M.; Jog, R.N.; Miwa, K.; Morikawa, M. Effect of Exogenous General Plant Growth Regulators on the Growth of the Duckweed Lemna minor. Front. Chem. 2018, 6, 251. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Radulović, O.; Stanković, S.; Stanojević, O.; Vujčić, Z.; Dojnov, B.; Trifunović-Momčilov, M.; Marković, M. Antioxidative Responses of Duckweed (Lemna minor L.) to Phenol and Rhizosphere-Associated Bacterial Strain Hafnia paralvei C32-106/3. Antioxidants 2021, 10, 1719. https://doi.org/10.3390/antiox10111719
Radulović O, Stanković S, Stanojević O, Vujčić Z, Dojnov B, Trifunović-Momčilov M, Marković M. Antioxidative Responses of Duckweed (Lemna minor L.) to Phenol and Rhizosphere-Associated Bacterial Strain Hafnia paralvei C32-106/3. Antioxidants. 2021; 10(11):1719. https://doi.org/10.3390/antiox10111719
Chicago/Turabian StyleRadulović, Olga, Slaviša Stanković, Olja Stanojević, Zoran Vujčić, Biljana Dojnov, Milana Trifunović-Momčilov, and Marija Marković. 2021. "Antioxidative Responses of Duckweed (Lemna minor L.) to Phenol and Rhizosphere-Associated Bacterial Strain Hafnia paralvei C32-106/3" Antioxidants 10, no. 11: 1719. https://doi.org/10.3390/antiox10111719