
Physiological Functions of Thiol Peroxidases (Gpx1 and Prdx2) during Xenopus laevis Embryonic Development


 , , and
, , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Xenopus Growth Conditions and In Vitro Fertilization
2.2. Plasmids and mRNA Synthesis
2.3. Morpholino Oligonucleotides Design and Xenopus Embryos Microinjection
2.4. Whole-Mount In Situ Hybridization (WISH)
2.5. RT-PCR (Reverse Transcription–Polymerase Chain Reaction)
2.6. Transcriptomic Analysis
2.7. Terminal Deoxynucleotidal Transferase dUTP Nick End Labeling (TUNEL) Staining
2.8. In Vivo Imaging of ROS
2.9. Statistical Analysis
3. Results
3.1. Spatiotemporal Expression Pattern of gpx1 and prdx2 during Xenopus Embryogenesis
3.2. gpx1 Knockdown Leads to Malformed Eyes, but prdx2 Morphant Embryos Exhibited No Specific Malformation during Xenopus Embryonic Development
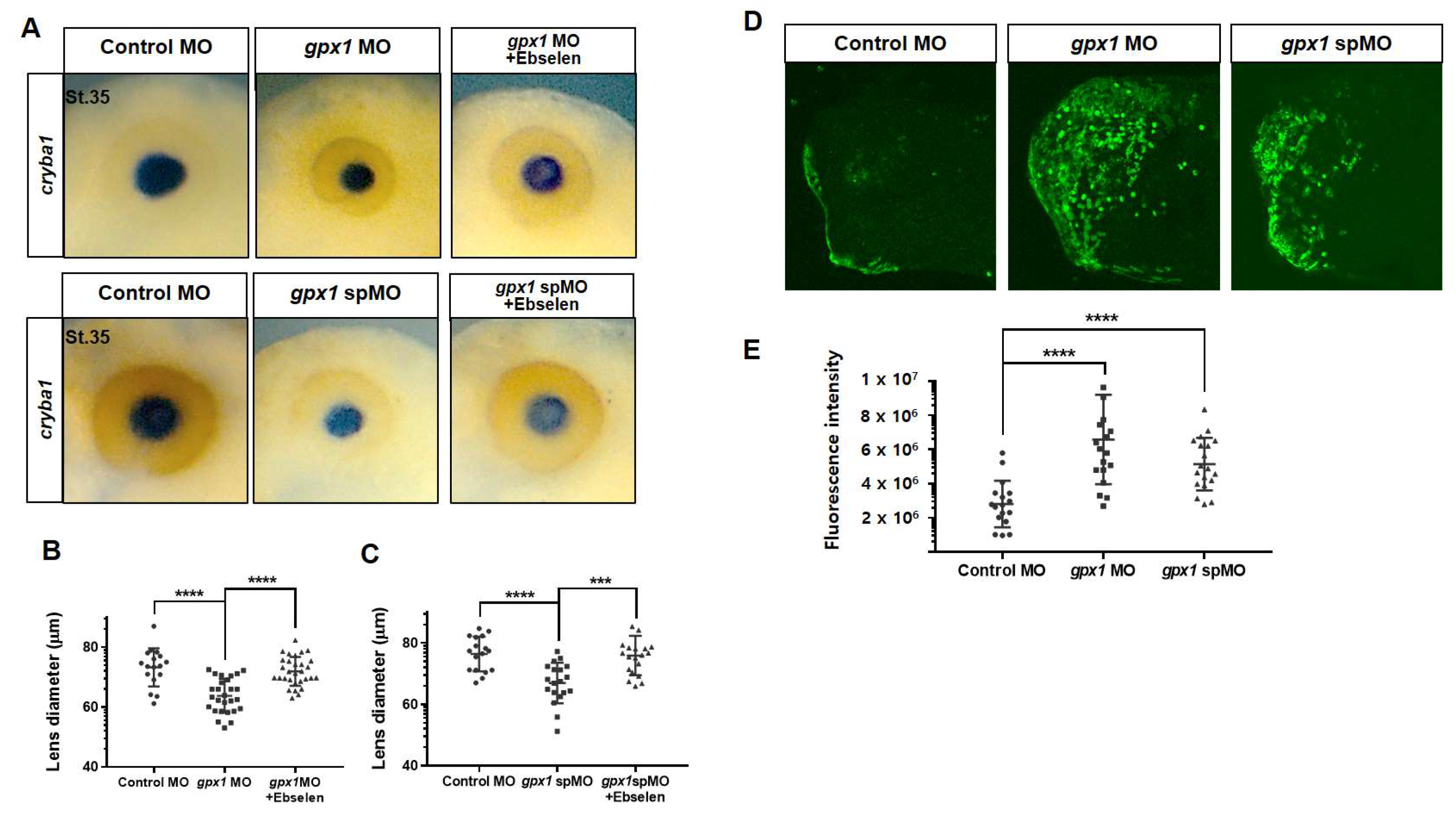
3.3. Gpx1 Interferes with Lens Development at the Molecular Level and Regulates ROS Levels in the Eye Regions during Embryogenesis
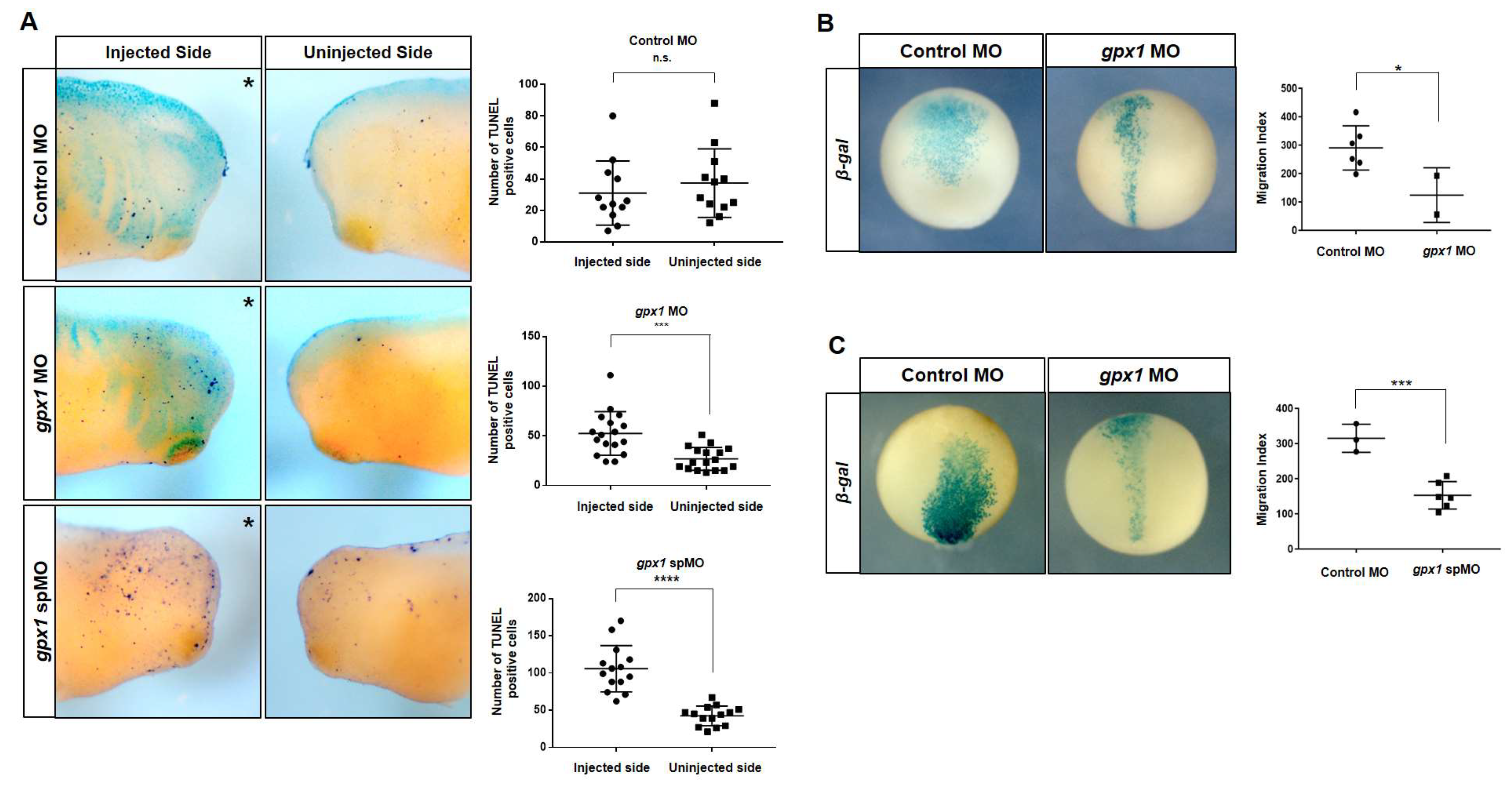
3.4. Gpx1 Regulates Cell Death and Interferes with Cell Migration during Xenopus Embryogenesis
3.5. Gpx1 Interferes with Wnt and Cadherin Signaling during Embryonic Eye Development in X. laevis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cadenas, E.; Davies, K.J. Mitochondrial free radical generation, oxidative stress, and aging. Free Radic. Biol. Med. 2000, 29, 222–230. [Google Scholar] [CrossRef]
- Dringen, R.; Pawlowski, P.G.; Hirrlinger, J. Peroxide detoxification by brain cells. J. Neurosci. Res. 2005, 79, 157–165. [Google Scholar] [CrossRef]
- Schieber, M.; Chandel, N.S. ROS function in redox signaling and oxidative stress. Curr. Biol. 2014, 24, R453–R462. [Google Scholar] [CrossRef] [Green Version]
- Timme-Laragy, A.R.; Hahn, M.E.; Hansen, J.M.; Rastogi, A.; Roy, M.A. Redox stress and signaling during vertebrate embryonic development: Regulation and responses. Semin. Cell Dev. Biol. 2018, 80, 17–28. [Google Scholar] [CrossRef]
- Imlay, J.A. Pathways of oxidative damage. Annu. Rev. Microbiol. 2003, 57, 395–418. [Google Scholar] [CrossRef] [PubMed]
- Covarrubias, L.; Hernández-García, D.; Schnabel, D.; Salas-Vidal, E.; Castro-Obregón, S. Function of reactive oxygen species during animal development: Passive or active? Dev. Biol. 2008, 320, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valko, M.; Leibfritz, D.; Moncol, J.; Cronin, M.T.; Mazur, M.; Telser, J. Free radicals and antioxidants in normal physiological functions and human disease. Int. J. Biochem. Cell Biol. 2007, 39, 44–84. [Google Scholar] [CrossRef]
- Bindoli, A.; Fukuto, J.M.; Forman, H.J. Thiol chemistry in peroxidase catalysis and redox signaling. Antioxid. Redox Signal. 2008, 10, 1549–1564. [Google Scholar] [CrossRef] [PubMed]
- Flohé, L.; Toppo, S.; Cozza, G.; Ursini, F. A comparison of thiol peroxidase mechanisms. Antioxid. Redox Signal. 2011, 15, 763–780. [Google Scholar] [CrossRef]
- Flohé, L.; Brigelius-Flohé, R.; Hatfield, D. Selenium: Its Molecular Biology and Role in Human Health; Selenoproteins Glutathione System; Kluwer: Norwel, MA, USA, 2001; pp. 157–178. [Google Scholar]
- Low, F.M.; Hampton, M.B.; Winterbourn, C.C. Peroxiredoxin 2 and peroxide metabolism in the erythrocyte. Antioxid. Redox Signal. 2008, 10, 1621–1630. [Google Scholar] [CrossRef]
- Hall, A.; Nelson, K.; Poole, L.B.; Karplus, P.A. Structure-based insights into the catalytic power and conformational dexterity of peroxiredoxins. Antioxid. Redox Signal. 2011, 15, 795–815. [Google Scholar] [CrossRef] [Green Version]
- Rhee, S.G.; Woo, H.A.; Kil, I.S.; Bae, S.H. Peroxiredoxin functions as a peroxidase and a regulator and sensor of local peroxides. J. Biol. Chem. 2012, 287, 4403–4410. [Google Scholar] [CrossRef] [Green Version]
- Lee, T.H.; Kim, S.U.; Yu, S.L.; Kim, S.H.; Park, D.S.; Moon, H.B.; Dho, S.H.; Kwon, K.S.; Kwon, H.J.; Han, Y.H.; et al. Peroxiredoxin II is essential for sustaining life span of erythrocytes in mice. Blood 2003, 101, 5033–5038. [Google Scholar] [CrossRef] [Green Version]
- Rhee, S.G.; Kang, S.W.; Chang, T.S.; Jeong, W.; Kim, K. Peroxiredoxin, a novel family of peroxidases. IUBMB Life 2001, 52, 35–41. [Google Scholar] [CrossRef]
- Ho, Y.S.; Magnenat, J.L.; Bronson, R.T.; Cao, J.; Gargano, M.; Sugawara, M.; Funk, C.D. Mice deficient in cellular glutathione peroxidase develop normally and show no increased sensitivity to hyperoxia. J. Biol. Chem. 1997, 272, 16644–16651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, Y.; Xu, T.; Li, S.; Ruan, H. GPX1, a biomarker for the diagnosis and prognosis of kidney cancer, promotes the progression of kidney cancer. Aging 2019, 11, 12165–12176. [Google Scholar] [CrossRef] [PubMed]
- Wei, R.; Qiu, H.; Xu, J.; Mo, J.; Liu, Y.; Gui, Y.; Huang, G.; Zhang, S.; Yao, H.; Huang, X.; et al. Expression and prognostic potential of GPX1 in human cancers based on data mining. Ann. Transl. Med. 2020, 8, 124. [Google Scholar] [CrossRef] [PubMed]
- Min, S.Y.; Kim, H.S.; Jung, E.J.; Jee, C.D.; Kim, W.H. Prognostic significance of glutathione peroxidase 1 (GPX1) down-regulation and correlation with aberrant promoter methylation in human gastric cancer. Anticancer Res. 2012, 32, 3169–3175. [Google Scholar] [PubMed]
- Hu, J.; Zhou, G.W.; Wang, N.; Wang, Y.J. GPX1 Pro198Leu polymorphism and breast cancer risk: A meta-analysis. Breast Cancer Res. Treat 2010, 124, 425–431. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.H.; Kim, S.U.; Kwon, T.H.; Lee, D.S.; Ha, H.L.; Park, D.S.; Woo, E.J.; Lee, S.H.; Kim, J.M.; Chae, H.B.; et al. Peroxiredoxin II is essential for preventing hemolytic anemia from oxidative stress through maintaining hemoglobin stability. Biochem. Biophys. Res. Commun. 2012, 426, 427–432. [Google Scholar] [CrossRef]
- Wang, W.; Wei, J.; Zhang, H.; Zheng, X.; Zhou, H.; Luo, Y.; Yang, J.; Deng, Q.; Huang, S.; Fu, Z. PRDX2 promotes the proliferation of colorectal cancer cells by increasing the ubiquitinated degradation of p53. Cell Death Dis. 2021, 12, 605. [Google Scholar] [CrossRef]
- Li, J.; Wang, C.; Wang, W.; Liu, L.; Zhang, Q.; Zhang, J.; Wang, B.; Wang, S.; Hou, L.; Gao, C.; et al. PRDX2 Protects Against Atherosclerosis by Regulating the Phenotype and Function of the Vascular Smooth Muscle Cell. Front. Cardiovasc. Med. 2021, 8, 624796. [Google Scholar] [CrossRef]
- Chae, S.; Lee, H.-K.; Kim, Y.-K.; Jung Sim, H.; Ji, Y.; Kim, C.; Ismail, T.; Park, J.-W.; Kwon, O.-S.; Kang, B.-S.; et al. Peroxiredoxin1, a novel regulator of pronephros development, influences retinoic acid and Wnt signaling by controlling ROS levels. Sci. Rep. 2017, 7, 8874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ji, Y.; Chae, S.; Lee, H.K.; Park, I.; Kim, C.; Ismail, T.; Kim, Y.; Park, J.W.; Kwon, O.S.; Kang, B.S.; et al. Peroxiredoxin5 Controls Vertebrate Ciliogenesis by Modulating Mitochondrial Reactive Oxygen Species. Antioxid. Redox Signal. 2019, 30, 1731–1745. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Ismail, T.; Kim, Y.; Chae, S.; Ryu, H.Y.; Lee, D.S.; Kwon, T.K.; Park, T.J.; Kwon, T.; Lee, H.S. Xenopus gpx3 Mediates Posterior Development by Regulating Cell Death during Embryogenesis. Antioxidants 2020, 9, 1265. [Google Scholar] [CrossRef]
- Kim, Y.; Jeong, Y.; Kwon, K.; Ismail, T.; Lee, H.K.; Kim, C.; Park, J.W.; Kwon, O.S.; Kang, B.S.; Lee, D.S.; et al. Physiological effects of KDM5C on neural crest migration and eye formation during vertebrate development. Epigenet. Chromatin 2018, 11, 72. [Google Scholar] [CrossRef] [PubMed]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, Y.; Feng, Q.; Kumagai, T.; Torikai, K.; Ohigashi, H.; Osawa, T.; Noguchi, N.; Niki, E.; Uchida, K. Ebselen, a glutathione peroxidase mimetic seleno-organic compound, as a multifunctional antioxidant. Implication for inflammation-associated carcinogenesis. J. Biol. Chem. 2002, 277, 2687–2694. [Google Scholar] [CrossRef] [Green Version]
- Zigler, J.S., Jr.; Sinha, D. βA3/A1-crystallin: More than a lens protein. Prog. Retin. Eye Res. 2015, 44, 62–85. [Google Scholar] [CrossRef]
- Lubos, E.; Loscalzo, J.; Handy, D.E. Glutathione peroxidase-1 in health and disease: From molecular mechanisms to therapeutic opportunities. Antioxid. Redox Signal. 2011, 15, 1957–1997. [Google Scholar] [CrossRef] [Green Version]
- Flentjar, N.J.; Crack, P.J.; Boyd, R.; Malin, M.; de Haan, J.B.; Hertzog, P.; Kola, I.; Iannello, R. Mice lacking glutathione peroxidase-1 activity show increased TUNEL staining and an accelerated inflammatory response in brain following a cold-induced injury. Exp. Neurol. 2002, 177, 9–20. [Google Scholar] [CrossRef]
- Ufer, C.; Wang, C. The roles of glutathione peroxidases during embryo development. Front. Mol. Neurosci. 2011, 4, 12. [Google Scholar] [CrossRef] [Green Version]
- Fujimura, N. WNT/β-Catenin Signaling in Vertebrate Eye Development. Front. Cell Dev. Biol. 2016, 4, 138. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Chung, J.-Y.; Rai, U.; Esumi, N. Cadherins in the retinal pigment epithelium (RPE) revisited: P-cadherin is the highly dominant cadherin expressed in human and mouse RPE in vivo. PLoS ONE 2018, 13, e0191279. [Google Scholar] [CrossRef] [Green Version]
- Walker, J.; Menko, A.S. Integrins in lens development and disease. Exp. Eye Res. 2009, 88, 216–225. [Google Scholar] [CrossRef] [Green Version]
- Fokina, V.M.; Frolova, E.I. Expression patterns of Wnt genes during development of an anterior part of the chicken eye. Dev. Dyn. Off. Publ. Am. Assoc. Anat. 2006, 235, 496–505. [Google Scholar] [CrossRef] [Green Version]
- Etzrodt, J.; Krishna-K, K.; Redies, C. Expression of classic cadherins and delta-protocadherins in the developing ferret retina. BMC Neurosci. 2009, 10, 153. [Google Scholar] [CrossRef] [Green Version]
- Flach, H.; Basten, T.; Schreiner, C.; Dietmann, P.; Greco, S.; Nies, L.; Roßmanith, N.; Walter, S.; Kühl, M.; Kühl, S.J. Retinol binding protein 1 affects Xenopus anterior neural development via all-trans retinoic acid signaling. Dev. Dyn. 2021, 250, 1096–1112. [Google Scholar] [CrossRef] [PubMed]
- Abouzeid, H.; Favez, T.; Schmid, A.; Agosti, C.; Youssef, M.; Marzouk, I.; El Shakankiry, N.; Bayoumi, N.; Munier, F.L.; Schorderet, D.F. Mutations in ALDH1A3 represent a frequent cause of microphthalmia/anophthalmia in consanguineous families. Hum. Mutat. 2014, 35, 949–953. [Google Scholar] [CrossRef] [PubMed]
- Alfano, G.; Conte, I.; Caramico, T.; Avellino, R.; Arnò, B.; Pizzo, M.T.; Tanimoto, N.; Beck, S.C.; Huber, G.; Dollé, P.; et al. Vax2 regulates retinoic acid distribution and cone opsin expression in the vertebrate eye. Development 2011, 138, 261–271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Halbleib, J.M.; Nelson, W.J. Cadherins in development: Cell adhesion, sorting, and tissue morphogenesis. Genes Dev. 2006, 20, 3199–3214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, D.; Liu, L.; Hu, Z.; Song, Z.; Wang, Y.; Chen, Z. Evaluation of the oxidative stress-related genes ALOX5, ALOX5AP, GPX1, GPX3 and MPO for contribution to the risk of type 2 diabetes mellitus in the Han Chinese population. Diabetes Vasc. Dis. Res. 2018, 15, 336–339. [Google Scholar] [CrossRef] [Green Version]
- Mohammedi, K.; Patente, T.A.; Bellili-Muñoz, N.; Driss, F.; Le Nagard, H.; Fumeron, F.; Roussel, R.; Hadjadj, S.; Corrêa-Giannella, M.L.; Marre, M.; et al. Glutathione peroxidase-1 gene (GPX1) variants, oxidative stress and risk of kidney complications in people with type 1 diabetes. Metabolism 2016, 65, 12–19. [Google Scholar] [CrossRef]
- Blum, M.; Beyer, T.; Weber, T.; Vick, P.; Andre, P.; Bitzer, E.; Schweickert, A. Xenopus, an ideal model system to study vertebrate left-right asymmetry. Dev. Dyn. 2009, 238, 1215–1225. [Google Scholar] [CrossRef] [PubMed]
- Shaping embryonic development. Nat. Chem. Biol. 2017, 13, 559. [CrossRef] [Green Version]
- Donner, A.L.; Lachke, S.A.; Maas, R.L. Lens induction in vertebrates: Variations on a conserved theme of signaling events. Semin. Cell Dev. Biol. 2006, 17, 676–685. [Google Scholar] [CrossRef]
- Seigfried, F.A.; Cizelsky, W.; Pfister, A.S.; Dietmann, P.; Walther, P.; Kühl, M.; Kühl, S.J. Frizzled 3 acts upstream of Alcam during embryonic eye development. Dev. Biol. 2017, 426, 69–83. [Google Scholar] [CrossRef] [PubMed]
- Van Raay, T.J.; Vetter, M.L. Wnt/frizzled signaling during vertebrate retinal development. Dev. Neurosci. 2004, 26, 352–358. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, H.; Lee, N.Y.; Kim, Y.; Choi, H.-S.; Ismail, T.; Ryu, H.-Y.; Cho, D.-H.; Ryoo, Z.Y.; Lee, D.-S.; Kwon, T.K.; et al. Physiological Functions of Thiol Peroxidases (Gpx1 and Prdx2) during Xenopus laevis Embryonic Development. Antioxidants 2021, 10, 1636. https://doi.org/10.3390/antiox10101636
Lee H, Lee NY, Kim Y, Choi H-S, Ismail T, Ryu H-Y, Cho D-H, Ryoo ZY, Lee D-S, Kwon TK, et al. Physiological Functions of Thiol Peroxidases (Gpx1 and Prdx2) during Xenopus laevis Embryonic Development. Antioxidants. 2021; 10(10):1636. https://doi.org/10.3390/antiox10101636
Chicago/Turabian StyleLee, Hongchan, Na Young Lee, Youni Kim, Hong-Seok Choi, Tayaba Ismail, Hong-Yeoul Ryu, Dong-Hyung Cho, Zae Young Ryoo, Dong-Seok Lee, Taeg Kyu Kwon, and et al. 2021. "Physiological Functions of Thiol Peroxidases (Gpx1 and Prdx2) during Xenopus laevis Embryonic Development" Antioxidants 10, no. 10: 1636. https://doi.org/10.3390/antiox10101636