
The Recovery of Cognitive and Affective Deficiencies Linked with Chronic Osteoarthritis Pain and Implicated Pathways by Slow-Releasing Hydrogen Sulfide Treatment

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Induction of Osteoarthritis Pain
2.3. Mechanical Allodynia
2.4. Measurement of Grip Strength
2.5. Cognitive Behavior
2.6. Depressive Behavioral Tests
2.7. Anxiety Behavioral Tests
2.8. Western Blot Analysis
2.9. Experimental Procedures
2.10. Drugs
2.11. Statistical Analyses
3. Results
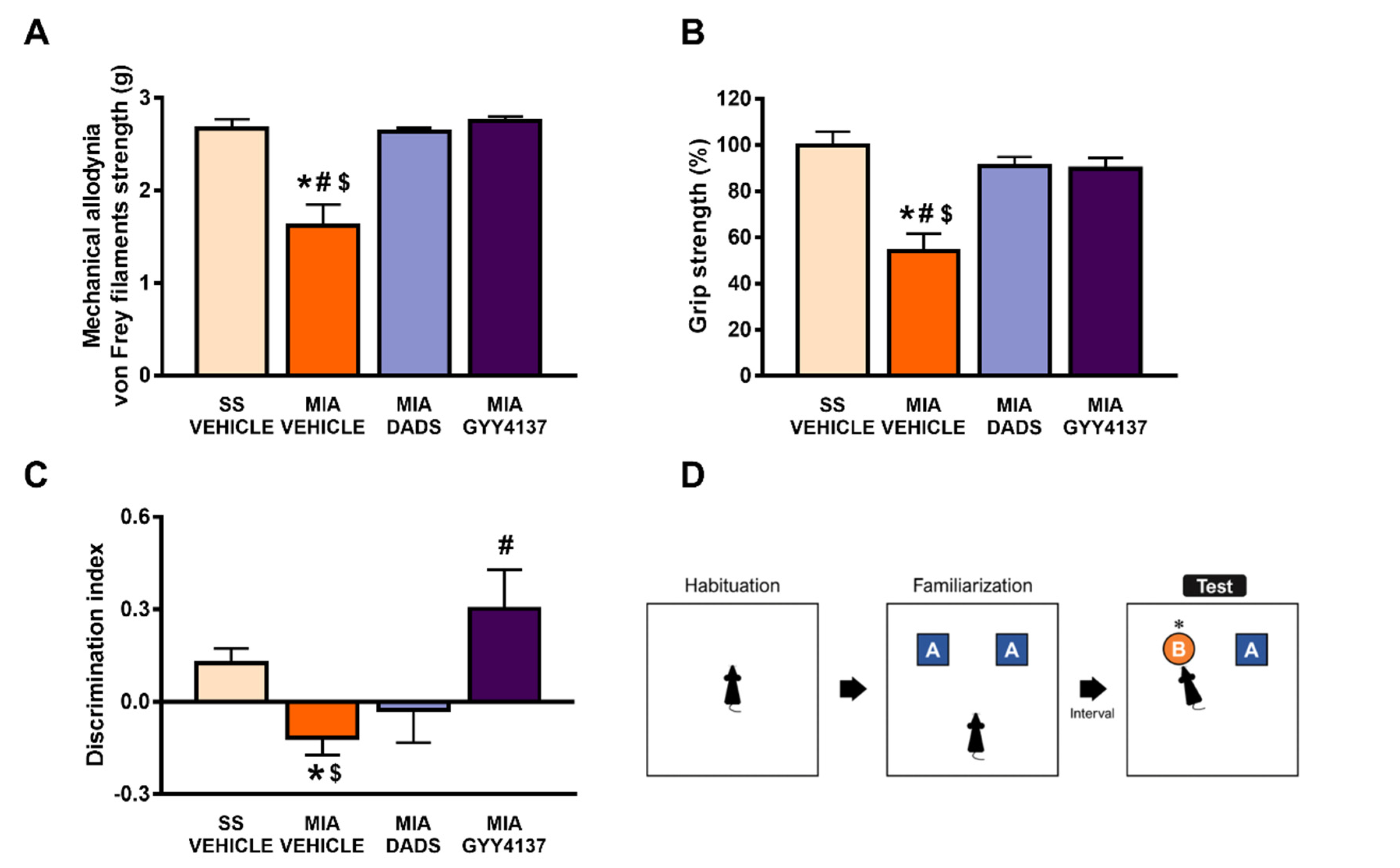
3.1. Treatment with DADS and/or GYY4137 Inhibits Osteoarthritis-Induced Cognitive Impairment, Hind Limb Grip Strength Deficits and Mechanical Allodynia
3.2. The Inhibition of the Anxiodepressive-Like Behaviors of DADS and GYY4137 in Animals with Chronic Osteoarthritis Pain
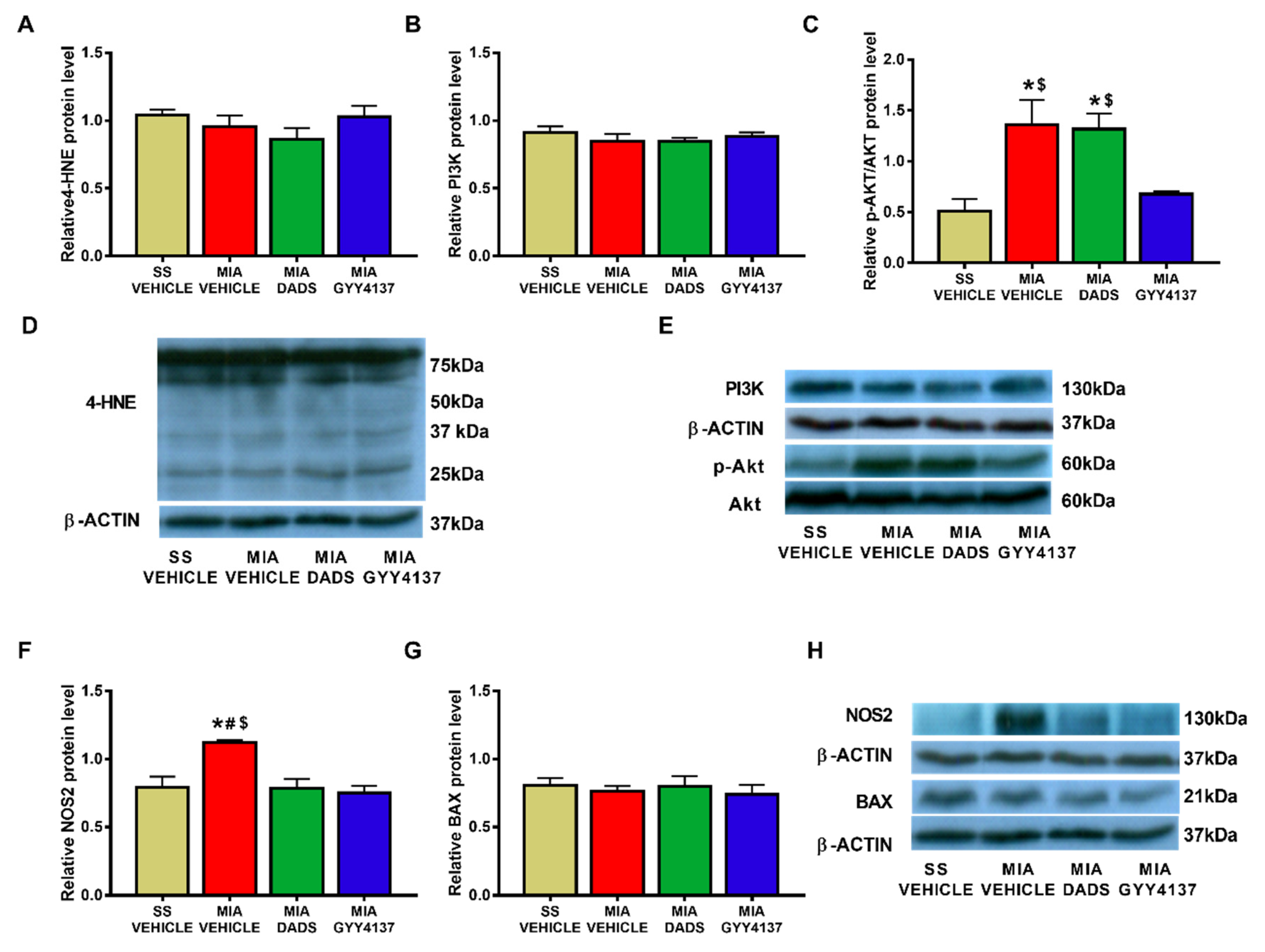
3.3. Effects of DADS and GYY4137 on the Protein Levels of 4-HNE, PI3K, p-Akt, NOS2 and BAX in the Amygdala of MIA-Injected Mice
3.4. Effects of DADS and GYY4137 on the Expression of 4-HNE, PI3K, p-Akt, NOS2 and BAX in the Periaqueductal Gray Matter of MIA-Injected Mice
3.5. Effects of DADS and GYY4137 on the Protein Levels of 4-HNE, PI3K, p-Akt, NOS2 and BAX in the Infralimbic Cortex of Mice with Osteoarthritic Pain
3.6. Effects of Treatment with DADS or GYY4137 on the Protein Levels of 4-HNE, PI3K, p-Akt, NOS2 and BAX in the Anterior Cingulate Cortex of MIA-Injected Mice
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bannuru, R.R.; Osani, M.C.; Vaysbrot, E.E.; Arden, N.K.; Bennell, K.; Bierma-Zeinstra, S.M.A.; Kraus, V.B.; Lohmander, L.S.; Abbott, J.H.; Bhandari, M.; et al. OARSI guidelines for the non-surgical management of knee, hip, and polyarticular osteoarthritis. Osteoarthr. Cartil. 2019, 27, 1578–1589. [Google Scholar] [CrossRef] [Green Version]
- Lucarini, E.; Micheli, L.; Martelli, A.; Testai, L.; Calderone, V.; Ghelardini, C.; Di Cesare Mannelli, L. Efficacy of isothiocyanate-based compounds on different forms of persistent pain. J. Pain. Res. 2018, 11, 2905–2913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alcaraz, M.J.; Guillén, M.I.; Ferrándiz, M.L. Emerging therapeutic agents in osteoarthritis. Biochem. Pharmacol. 2019, 165, 4–16. [Google Scholar] [CrossRef]
- Jacobs, C.A.; Mace, R.A.; Greenberg, J.; Popok, P.J.; Reichman, M.; Lattermann, C.; Burris, J.L.; Macklin, E.A.; Vranceanu, A.M. Development of a mind body program for obese knee osteoarthritis patients with comorbid depression. Contemp. Clin. Trials. Commun. 2021, 21, 100720. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, C.A.; Vranceanu, A.M.; Thompson, K.L.; Lattermann, C. Rapid Progression of Knee Pain and Osteoarthritis Biomarkers Greatest for Patients with Combined Obesity and Depression: Data from the Osteoarthritis Initiative. Cartilage 2020, 11, 38–46. [Google Scholar] [CrossRef] [PubMed]
- Hart, R.P.; Martelli, M.F.; Zasler, N.D. Chronic pain and neuropsychological functioning. Neuropsychol. Rev. 2000, 10, 131–149. [Google Scholar] [CrossRef] [PubMed]
- Luerding, R.; Weigand, T.; Bogdahn, U.; Schmidt-Wilcke, T. Working memory performance is correlated with local brain morphology in the medial frontal and anterior cingulate cortex in fibromyalgia patients: Structural correlates of pain-cognition interaction. Brain 2008, 131, 3222–3231. [Google Scholar] [CrossRef] [Green Version]
- Sallinen, M.; Marit Mengshoel, A. Memory gaps, lost words and crucial mistakes—Men’s experiences of cognitive difficulties in fibromyalgia. Chronic Illn. 2021, 17, 41–52. [Google Scholar] [CrossRef] [Green Version]
- Sharma, A.; Kudesia, P.; Shi, Q.; Gandhi, R. Anxiety and depression in patients with osteoarthritis: Impact and management challenges. Open Access Rheumatol. 2016, 8, 103–113. [Google Scholar] [CrossRef] [Green Version]
- Sun, K.; Luo, J.; Guo, J.; Yao, X.; Jing, X.; Guo, F. The PI3K/AKT/mTOR signaling pathway in osteoarthritis: A narrative review. Osteoarthr. Cartil. 2020, 28, 400–409. [Google Scholar] [CrossRef]
- Patki, G.; Solanki, N.; Atrooz, F.; Allam, F.; Salim, S. Depression, anxiety-like behavior and memory impairment are associated with increased oxidative stress and inflammation in a rat model of social stress. Brain Res. 2013, 1539, 73–86. [Google Scholar] [CrossRef] [Green Version]
- Pol, O. The role of carbon monoxide, heme oxygenase 1, and the Nrf2 transcription factor in the modulation of chronic pain and their interactions with opioids and cannabinoids. Med. Res. Rev. 2021, 41, 136–155. [Google Scholar] [CrossRef] [PubMed]
- Cravero, J.D.; Carlson, C.S.; Im, H.J.; Yammani, R.R.; Long, D.; Loeser, R.F. Increased expression of the Akt/PKB inhibitor TRB3 in osteoarthritic chondrocytes inhibits insulin-like growth factor 1-mediated cell survival and proteoglycan synthesis. Arthritis Rheum. 2009, 60, 492–500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, F.; Wang, Y.; Hemmings, B.A.; Rüegg, C.; Xue, G. PKB/Akt-dependent regulation of inflammation in cancer. Semin. Cancer Biol. 2018, 48, 62–69. [Google Scholar] [CrossRef] [Green Version]
- Qiao, X.; Gai, H.; Su, R.; Deji, C.; Cui, J.; Lai, J.; Zhu, Y. PI3K-AKT-GSK3β-CREB signaling pathway regulates anxiety-like behavior in rats following alcohol withdrawal. J. Affect. Disord. 2018, 235, 96–104. [Google Scholar] [CrossRef] [PubMed]
- Kitagishi, Y.; Kobayashi, M.; Kikuta, K.; Matsuda, S. Roles of PI3K/AKT/GSK3/mTOR Pathway in Cell Signaling of Mental Illnesses. Depress. Res. Treat. 2012, 2012, 752563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, L.; Liu, C.; Wan, Y.; Yan, H.; Li, T. Effect of HDAC2/Inpp5f on neuropathic pain and cognitive function through regulating PI3K/Akt/GSK-3β signal pathway in rats with neuropathic pain. Exp. Ther. Med. 2019, 18, 678–684. [Google Scholar] [CrossRef]
- Lepetsos, P.; Papavassiliou, A.G. ROS/oxidative stress signaling in osteoarthritis. Biochim. Biophys. Acta 2016, 1862, 576–591. [Google Scholar] [CrossRef]
- Zahan, O.M.; Serban, O.; Gherman, C.; Fodor, D. The evaluation of oxidative stress in osteoarthritis. Med. Pharm. Rep. 2020, 93, 12–22. [Google Scholar] [CrossRef]
- Salim, S.; Asghar, M.; Taneja, M.; Hovatta, I.; Chugh, G.; Vollert, C.; Vu, A. Potential contribution of oxidative stress and inflammation to anxiety and hypertension. Brain Res. 2011, 1404, 63–71. [Google Scholar] [CrossRef] [Green Version]
- Brocardo, P.S.; Boehme, F.; Patten, A.; Cox, A.; Gil-Mohapel, J.; Christie, B.R. Anxiety- and depression-like behaviors are accompanied by an increase in oxidative stress in a rat model of fetal alcohol spectrum disorders: Protective effects of voluntary physical exercise. Neuropharmacology 2012, 62, 1607–1618. [Google Scholar] [CrossRef]
- Yamada, E.F.; Salgueiro, A.F.; Goulart, A.D.S.; Mendes, V.P.; Anjos, B.L.; Folmer, V.; da Silva, M.D. Evaluation of monosodium iodoacetate dosage to induce knee osteoarthritis: Relation with oxidative stress and pain. Int. J. Rheum. Dis. 2019, 22, 399–410. [Google Scholar] [CrossRef]
- Ferreira-Chamorro, P.; Redondo, A.; Riego, G.; Leánez, S.; Pol, O. Sulforaphane Inhibited the Nociceptive Responses, Anxiety- and Depressive-Like Behaviors Associated with Neuropathic Pain and Improved the Anti-allodynic Effects of Morphine in Mice. Front. Pharmacol. 2018, 9, 1332. [Google Scholar] [CrossRef] [PubMed]
- Cabarga, L.; Batallé, G.; Pol, O. Treatment with slow-releasing hydrogen sulfide donors inhibits the nociceptive and depressive-like behaviours accompanying chronic neuropathic pain: Endogenous antioxidant system activation. J. Psychopharmacol. 2020, 34, 737–749. [Google Scholar] [CrossRef] [PubMed]
- Abramson, S.B. Osteoarthritis and nitric oxide. Osteoarthr. Cartil. 2008, 16, S15–S20. [Google Scholar] [CrossRef] [Green Version]
- Leonidou, A.; Lepetsos, P.; Mintzas, M.; Kenanidis, E.; Macheras, G.; Tzetis, M.; Potoupnis, M.; Tsiridis, E. Inducible nitric oxide synthase as a target for osteoarthritis treatment. Expert Opin. Ther. Targets 2018, 22, 299–318. [Google Scholar] [CrossRef] [PubMed]
- Pelletier, J.P.; Jovanovic, D.V.; Lascau-Coman, V.; Fernandes, J.C.; Manning, P.T.; Connor, J.R.; Currie, M.G.; Martel-Pelletier, J. Selective inhibition of inducible nitric oxide synthase reduces progression of experimental osteoarthritis in vivo: Possible link with the reduction in chondrocyte apoptosis and caspase 3 level. Arthritis Rheum. 2000, 43, 1290–1299. [Google Scholar] [CrossRef]
- Gerasimova, E.; Lebedeva, J.; Yakovlev, A.; Zefirov, A.; Giniatullin, R.; Sitdikova, G. Mechanisms of hydrogen sulfide (H2S) action on synaptic transmission at the mouse neuromuscular junction. Neuroscience 2015, 303, 577–585. [Google Scholar] [CrossRef] [PubMed]
- Rong, W.; Kimura, H.; Grundy, D. The neurophysiology of hydrogen sulfide. Inflamm. Allergy Drug Targets 2011, 10, 109–117. [Google Scholar] [CrossRef]
- Wallace, J.L.; Vaughan, D.; Dicay, M.; MacNaughton, W.K.; de Nucci, G. Hydrogen Sulfide-Releasing Therapeutics: Translation to the Clinic. Antioxid. Redox Signal. 2018, 28, 1533–1540. [Google Scholar] [CrossRef]
- Guo, J.; Li, G.; Yang, L. Role of H2S in pain: Growing evidences of mystification. Eur. J. Pharmacol. 2020, 883, 173322. [Google Scholar] [CrossRef]
- Fox, B.; Schantz, J.T.; Haigh, R.; Wood, M.E.; Moore, P.K.; Viner, N.; Spencer, J.P.; Winyard, P.G.; Whiteman, M. Inducible hydrogen sulfide synthesis in chondrocytes and mesenchymal progenitor cells: Is H2S a novel cytoprotective mediator in the inflamed joint? J. Cell. Mol. Med. 2012, 16, 896–910. [Google Scholar] [CrossRef]
- Vela-Anero, Á.; Hermida-Gómez, T.; Gato-Calvo, L.; Vaamonde-García, C.; Díaz-Prado, S.; Meijide-Faílde, R.; Blanco, F.J.; Burguera, E.F. Long-term effects of hydrogen sulfide on the anabolic-catabolic balance of articular cartilage in vitro. Nitric Oxide 2017, 70, 42–50. [Google Scholar] [CrossRef] [Green Version]
- Aytekin, K.; Erhan, S.Ş.; Erişgin, Z.; Esenyel, C.Z.; Takır, S. Intra-articular injection of hydrogen sulfide decreased the progression of gonarthrosis. Can. J. Physiol. Pharmacol. 2019, 97, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Vaamonde-García, C.; Burguera, E.F.; Vela-Anero, Á.; Hermida-Gómez, T.; Filgueira-Fernández, P.; Fernández-Rodríguez, J.A.; Meijide-Faílde, R.; Blanco, F.J. Intraarticular Administration Effect of Hydrogen Sulfide on an In Vivo Rat Model of Osteoarthritis. Int. J. Mol. Sci. 2020, 21, 7421. [Google Scholar] [CrossRef] [PubMed]
- Batallé, G.; Cabarga, L.; Pol, O. The Inhibitory Effects of Slow-Releasing Hydrogen Sulfide Donors in the Mechanical Allodynia, Grip Strength Deficits, and Depressive-Like Behaviors Associated with Chronic Osteoarthritis Pain. Antioxidants 2020, 9, 31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yue, L.; Ma, L.Y.; Cui, S.; Liu, F.Y.; Yi, M.; Wan, Y. Brain-derived neurotrophic factor in the infralimbic cortex alleviates inflammatory pain. Neurosci. Lett. 2017, 655, 7–13. [Google Scholar] [CrossRef]
- Thompson, J.M.; Neugebauer, V. Amygdala Plasticity and Pain. Pain. Res. Manag. 2017, 2017, 8296501. [Google Scholar] [CrossRef] [Green Version]
- Russell, M.D.; Barrick, T.R.; Howe, F.A.; Sofat, N. Reduced anterior cingulate grey matter volume in painful hand osteoarthritis. Rheumatol. Int. 2018, 38, 1429–1435. [Google Scholar] [CrossRef] [Green Version]
- Kummer, K.K.; Mitrić, M.; Kalpachidou, T.; Kress, M. The Medial Prefrontal Cortex as a Central Hub for Mental Comorbidities Associated with Chronic Pain. Int. J. Mol. Sci. 2020, 21, 3440. [Google Scholar] [CrossRef]
- Chaplan, S.R.; Bach, F.W.; Pogrel, J.W.; Chung, J.M.; Yaksh, T.L. Quantitative assessment of tactile allodynia in the rat paw. J. Neurosci. Methods 1994, 53, 55–63. [Google Scholar] [CrossRef]
- Montilla-García, A.; Tejada, M.A.; Perazzoli, G.; Entrena, J.M.; Portillo-Salido, E.; Fernández-Segura, E.; Cañizares, F.J.; Cobos, E.J. Grip strength in mica with joint inflammation: A rheumatology function test sensitive to pain and analgesia. Neuropharmacology 2017, 125, 231–242. [Google Scholar] [CrossRef] [PubMed]
- Federman, N.; de la Fuente, V.; Zalcman, G.; Corbi, N.; Onori, A.; Passananti, C.; Romano, A. Nuclear factor κB-dependent histone acetylation is specifically involved in persistent forms of memory. J. Neurosci. 2013, 33, 7603–7614. [Google Scholar] [CrossRef] [PubMed]
- Porsolt, R.D.; Le Pichon, M.; Jalfre, M. Depression: A new animal model sensitive to antidepressant treatments. Nature 1977, 266, 730–732. [Google Scholar] [CrossRef]
- Steru, L.; Chermat, R.; Thierry, B.; Simon, P. The tail suspension test: A new method for screening antidepressants in mice. Psychopharmacology 1985, 85, 367–370. [Google Scholar] [CrossRef]
- Walf, A.A.; Frye, C.A. The use of the elevated plus maze as an assay of anxiety-related behavior in rodents. Nat. Protoc. 2007, 2, 322–328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bai, X.; Batallé, G.; Pol, O. The Anxiolytic and Antidepressant Effects of Diallyl Disulfide and GYY4137 in Animals with Chronic Neuropathic Pain. Antioxidants 2021, 10, 1074. [Google Scholar] [CrossRef]
- Berryman, C.; Stanton, T.R.; Jane Bowering, K.; Tabor, A.; McFarlane, A.; Lorimer Moseley, G. Evidence for working memory deficits in chronic pain: A systematic review and meta-analysis. Pain 2013, 154, 1181–1196. [Google Scholar] [CrossRef]
- Tyrtyshnaia, A.; Manzhulo, I. Neuropathic Pain Causes Memory Deficits and Dendrite Tree Morphology Changes in Mouse Hippocampus. J. Pain Res. 2020, 13, 345–354. [Google Scholar] [CrossRef]
- Negrete, R.; García Gutiérrez, M.S.; Manzanares, J.; Maldonado, R. Involvement of the dynorphin/KOR system on the nociceptive, emotional and cognitive manifestations of joint pain in mice. Neuropharmacology 2017, 116, 315–327. [Google Scholar] [CrossRef] [Green Version]
- Carcolé, M.; Zamanillo, D.; Merlos, M.; Fernández-Pastor, B.; Cabañero, D.; Maldonado, R. Blockade of the Sigma-1 Receptor Relieves Cognitive and Emotional Impairments Associated to Chronic Osteoarthritis Pain. Front. Pharmacol. 2019, 10, 468. [Google Scholar] [CrossRef] [Green Version]
- Cao, H.; Zang, K.K.; Han, M.; Zhao, Z.Q.; Wu, G.C.; Zhang, Y.Q. Inhibition of p38 mitogen-activated protein kinase activation in the rostral anterior cingulate cortex attenuates pain-related negative emotion in rats. Brain Res. Bull. 2014, 107, 79–88. [Google Scholar] [CrossRef] [PubMed]
- Graeff, F.G.; Del-Ben, C.M. Neurobiology of panic disorder: From animal models to brain neuroimaging. Neurosci. Biobehav. Rev. 2008, 32, 1326–1335. [Google Scholar] [CrossRef] [PubMed]
- Barroso, J.; Wakaizumi, K.; Reis, A.M.; Baliki, M.; Schnitzer, T.J.; Galhardo, V.; Apkarian, A.V. Reorganization of functional brain network architecture in chronic osteoarthritis pain. Hum. Brain Mapp. 2021, 42, 1206–1222. [Google Scholar] [CrossRef] [PubMed]
- Sheng, J.; Liu, S.; Wang, Y.; Cui, R.; Zhang, X. The Link between Depression and Chronic Pain: Neural Mechanisms in the Brain. Neural Plast. 2017, 2017, 9724371. [Google Scholar] [CrossRef] [PubMed]
- Karimi, S.A.; Hosseinmardi, N.; Janahmadi, M.; Sayyah, M.; Hajisoltani, R. The protective effect of hydrogen sulfide (H2S) on traumatic brain injury (TBI) induced memory deficits in rats. Brain Res. Bull. 2017, 134, 177–182. [Google Scholar] [CrossRef]
- Chen, W.L.; Xie, B.; Zhang, C.; Xu, K.L.; Niu, Y.Y.; Tang, X.Q.; Zhang, P.; Zou, W.; Hu, B.; Tian, Y. Antidepressant-like and anxiolytic-like effects of hydrogen sulfide in behavioral models of depression and anxiety. Behav. Pharmacol. 2013, 24, 590–597. [Google Scholar] [CrossRef]
- Krause-Utz, A.; Winter, D.; Schriner, F.; Chiu, C.D.; Lis, S.; Spinhoven, P.; Bohus, M.; Schmahl, C.; Elzinga, B.M. Reduced amygdala reactivity and impaired working memory during dissociation in borderline personality disorder. Eur. Arch. Psychiatry Clin. Neurosci. 2018, 268, 401–415. [Google Scholar] [CrossRef]
- Martínez-Torres, N.I.; Vázquez-Hernández, N.; Martín-Amaya-Barajas, F.L.; Flores-Soto, M.; González-Burgos, I. Ibotenic acid induced lesions impair the modulation of dendritic spine plasticity in the prefrontal cortex and amygdala, a phenomenon that underlies working memory and social behavior. Eur. J. Pharmacol. 2021, 896, 173883. [Google Scholar] [CrossRef]
- Zhao, F.; Lei, F.; Yan, X.; Zhang, S.; Wang, W.; Zheng, Y. Protective Effects of Hydrogen Sulfide Against Cigarette Smoke Exposure-Induced Placental Oxidative Damage by Alleviating Redox Imbalance via Nrf2 Pathway in Rats. Cell. Physiol. Biochem. 2018, 48, 1815–1828. [Google Scholar] [CrossRef]
- Sun, H.; Qi, L.; Wang, S.; Li, X.; Li, C. Hydrogen sulfide is expressed in the human and the rat cultured nucleus pulposus cells and suppresses apoptosis induced by hypoxia. PLoS ONE 2018, 13, e0192556. [Google Scholar] [CrossRef]
- Chen, M.; Pritchard, C.; Fortune, D.; Kodi, P.; Grados, M. Hydrogen sulfide: A target to modulate oxidative stress and neuroplasticity for the treatment of pathological anxiety. Expert Rev. Neurother. 2020, 20, 109–121. [Google Scholar] [CrossRef] [PubMed]
- Zeng, J.; Li, S.; Zhang, C.; Huang, G.; Yu, C. The Mechanism of Hyperalgesia and Anxiety Induced by Remifentanil: Phosphorylation of GluR1 Receptors in the Anterior Cingulate Cortex. J. Mol. Neurosci. 2018, 65, 93–101. [Google Scholar] [CrossRef] [PubMed]
- Darvish-Ghane, S.; Quintana, C.; Beaulieu, J.M.; Martin, L.J. D1 receptors in the anterior cingulate cortex modulate basal mechanical sensitivity threshold and glutamatergic synaptic transmission. Mol. Brain 2020, 13, 121. [Google Scholar] [CrossRef] [PubMed]
- Xue, M.; Zhou, S.B.; Liu, R.H.; Chen, Q.Y.; Zhuo, M.; Li, X.H. NMDA Receptor-Dependent Synaptic Depression in Potentiated Synapses of the Anterior Cingulate Cortex of adult Mice. Mol. Pain 2021, 17, 17448069211018045. [Google Scholar] [CrossRef] [PubMed]
- Su, S.; Li, M.; Wu, D.; Cao, J.; Ren, X.; Tao, Y.X.; Zang, W. Gene Transcript Alterations in the Spinal Cord, Anterior Cingulate Cortex, and Amygdala in Mice Following Peripheral Nerve Injury. Front. Cell Dev. Biol. 2021, 9, 634810. [Google Scholar] [CrossRef]
- Liu, J.; Chen, L.; Chen, X.; Hu, K.; Tu, Y.; Lin, M.; Huang, J.; Liu, W.; Wu, J.; Qiu, Z.; et al. Modulatory effects of different exercise modalities on the functional connectivity of the periaqueductal grey and ventral tegmental area in patients with knee osteoarthritis: A randomised multimodal magnetic resonance imaging study. Br. J. Anaesth. 2019, 123, 506–518. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Fox, B.; Keeble, J.; Salto-Tellez, M.; Winyard, P.G.; Wood, M.E.; Moore, P.K.; Whiteman, M. The complex effects of the slow-releasing hydrogen sulfide donor GYY4137 in a model of acute joint inflammation and in human cartilage cells. J. Cell Mol. Med. 2013, 17, 365–376. [Google Scholar] [CrossRef] [Green Version]
- Qabazard, B.; Masocha, W.; Khajah, M.; Phillips, O.A. H2S donor GYY4137 ameliorates paclitaxel-induced neuropathic pain in mice. Biomed. Pharmacother. 2020, 127, 110210. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Batallé, G.; Bai, X.; Pouso-Vázquez, E.; Roch, G.; Rodríguez, L.; Pol, O. The Recovery of Cognitive and Affective Deficiencies Linked with Chronic Osteoarthritis Pain and Implicated Pathways by Slow-Releasing Hydrogen Sulfide Treatment. Antioxidants 2021, 10, 1632. https://doi.org/10.3390/antiox10101632
Batallé G, Bai X, Pouso-Vázquez E, Roch G, Rodríguez L, Pol O. The Recovery of Cognitive and Affective Deficiencies Linked with Chronic Osteoarthritis Pain and Implicated Pathways by Slow-Releasing Hydrogen Sulfide Treatment. Antioxidants. 2021; 10(10):1632. https://doi.org/10.3390/antiox10101632
Chicago/Turabian StyleBatallé, Gerard, Xue Bai, Enric Pouso-Vázquez, Gerard Roch, Laura Rodríguez, and Olga Pol. 2021. "The Recovery of Cognitive and Affective Deficiencies Linked with Chronic Osteoarthritis Pain and Implicated Pathways by Slow-Releasing Hydrogen Sulfide Treatment" Antioxidants 10, no. 10: 1632. https://doi.org/10.3390/antiox10101632