1. Introduction
Shellfish (mollusks and crustaceans) species are the key dietary source of omega-3 (n-3) very-long-chain (VLC)-polyunsaturated fatty acids (PUFAs), mainly eicosapentaenoic (EPA; C20:5n3), and docosahexaenoic acids (DHA; C22:6n3), which play a crucial role in reducing the risk of cancer and cardiovascular diseases (CVD). Moreover, DHA-derived specialized pro-resolving mediators (SPMs, e.g., protectins, resolvins, and maresins) are critically crucial for neonatal brain development and mental and cognitive development [
1] and neuroprotection [
2,
3].
Though with the richness of VLC-n-3 PUFAs, shellfish are considered a vital component of a healthy diet, the high cholesterol content of some species is generally cited as a reason to limit their intake. However, in addition to cholesterol, shellfish species contain a significant amount of other sterols (called non-cholesterols sterols; NCS); some are unique to marine species [
4], derived from the food they consume (e.g., microalgae) and from endogenous metabolism. Plant-derived sterols (called phytosterols) are well known to reduce low-density plasma lipoprotein-cholesterol (LDL-C) levels [
5,
6,
7] and thereby lower the risk of CVD. Moreover, as antioxidants, phytosterols are well-known to scavenge harmful reactive oxygen species (ROS) [
8]. Furthermore, animal and human studies have demonstrated the anticancer [
9] and anti-inflammatory [
10] effects of phytosterols. Considering these facts, consuming a phytosterol-rich diet may provide health benefits.
Tocopherols and tocotrienols (collectively known as tocols, tocochromanols, or vitamin E) are key components of plant- and animal-derived lipids, which scavenge ROS, thus protecting lipids from oxidative degradation. Given their functional role in controlling cellular oxidative stress, a diet rich in tocols minimizes the incidence of cancer, CVD, and neurodegenerative diseases [
11,
12,
13].
To the best of our knowledge, only a few detailed studies are available on the sterol composition of shellfish species [
4], while most studies reported only cholesterol and few other sterols, such as sitosterol, campesterol, or stigmasterol [
14,
15,
16], while many shellfish species contain still other sterols at significant levels. Moreover, culturing conditions and dietary factors can affect the sterol content of shellfish species harvested from different natural geographical locations [
4]. Similarly, the detailed composition of many shellfish species commonly consumed in Korea is not available. Furthermore, similarly to sterols, fatty acids’ composition in shellfish may vary with season [
17], sex (e.g., male vs. female crabs) [
18], culturing conditions (e.g., wild vs. cultured, and diet quality) [
19,
20], and several other factors [
21].
Considering the above, the present investigation aims to analyze the composition and contents of the major lipophilic compounds, including sterols, fatty acids, and tocols, of shellfish species, commonly consumed in Korea. Sterols were analyzed by gas chromatography (GC)-mass spectrometry (MS), fatty acids by GC-flame ionization detection (FID) and GC-MS, and tocols by high-performance liquid chromatography (HPLC)-diode-array detection (DAD). Moreover, to explore the antitumoral activities of the lipophilic constituents of shellfish species, the cytotoxicity potentials of the extracted lipids were determined against five different human cancer cells, including colon carcinoma (HCT116), epithelial melanoma (A2058), glioblastoma multiforme (T98G), lung carcinoma (A549), and adenocarcinoma (HeLa). The fatty acids, sterols, and tocols composition, and the cytotoxicity data reported herein may help explore shellfish species’ nutritional and anticancer potentials.
2. Materials and Methods
2.1. Reagents, Standards, and Raw Material
Authentic standards of sterols including cholesterol (>99% purity), cholestanol (>99% purity), brassicasterol (>98% purity), ergosterol (quality level of MQ300), and campesterol (90% purity) and fatty acids (37-Component FAME Mix, CRM47885, quality level of MQ100) were obtained from Merck Ltd., Seoul, South Korea. A mixed-tocols solution containing δ-, γ-, β-, and α-tocotrienol and δ-, γ-, β-, and α-tocopherol (purity adjusted) was purchased from ChromaDex, Inc. (Irvine, CA, USA). All organic solvents used for the extractions were HPLC grade and obtained from Daejung Chemicals and Metals Co., Ltd., Siheung-si, Korea.
A total of eleven shellfish species consumed in Korea were procured from the Garak market, Songpa-gu, Seoul, in October 2019 (
Table 1). Two kilograms of each species were brought to the lab, the edible flesh was manually separated and homogenized using a food processor, and 30-g portion was precisely aliquoted to falcon tubes and stored at −80 °C until analysis.
2.2. Extraction of Crude Lipids (Lipophilic Compounds)
Crude lipids, containing lipophilic compounds, were extracted from the edible portions of the studied shellfish species following our optimized protocol [
22], with minor modifications; it was initially based on a previous report [
23]. The detailed extraction procedure is illustrated in
Appendix A. The butylated hydroxytoluene (BHT:
w/
v; synthetic antioxidant) was added to the extraction solvent to minimize the degradation of lipophilic compounds [
24]. The crude lipids utilized for the cell culture analysis were extracted without using the BHT. The extracted crude lipids were utilized to analyze their fatty acids, tocols, and sterols, and to determine their anticancer potentials, as shown in
Appendix A. Tocols were analyzed by HPLC without hydrolysis, as it can degrade the tocols [
23]. The crude lipids were converted to fatty acid methyl esters (FAMEs) using a commercially available boron trifluoride-methanol solution (14% in methanol; Merck Ltd., Seoul, South Korea) as per the manufacturer’s guidelines, with minor modifications (
Appendix B). Similarly, before the GC-MS analysis, the crude lipids were hydrolyzed for sterol analysis, as mentioned in
Appendix B [
23].
2.3. HPLC Analysis of Tocols
The chromatographic separation of tocols was achieved utilizing an HPLC system (Model 1100; Agilent Technologies Canada, Inc., Mississauga, ON, Canada) equipped with a YMC C
30 column (250 × 4.6 mm, 5 μm; YMC, Wilmington, NC, USA) and a DAD. The solvent system, gradient elution pattern, flow rate, and detection wavelengths were used as previously optimized [
25].
2.4. FAMEs Determinations by Gas Chromatography (GC)-Flame Ionization Detection (FID) and GC-Mass Spectrometry (MS)
FAMEs were quantitatively analyzed by GC (Agilent 7890B, Agilent Technologies, Santa Clara, CA, USA) equipped with an FID and an SP-2560 capillary column (100 m, 0.20 μm film thickness, 0.25 mm ID; Merck KGaA, Darmstadt, Germany). The thermal program and other FID parameters were used as optimized in recent investigations [
26]. For precise identification, compounds’ mass spectra were recorded through the GS-MS system (QP2010 SE; Shimadzu, Japan), following a thermal program of GC-FID analysis. The identities of FAMEs were confirmed by comparing their fragmentation pattern with the authentic standards.
2.5. Calculation of Fat Quality Indices
Fat-quality indices, including total polyunsaturated fatty acids (PUFAs)/total saturated fatty acids (SFAs), n-3 PUFAs/n-6 PUFAs, hypocholesterolemic (h)/hypercholesterolemic (H) fatty acids ratios, atherogenic index (AI), and thrombogenic index (TI), were calculated from the fatty acid-profiling results [
27,
28,
29,
30]. The h/H, AI, and TI were calculated as per the following equations:
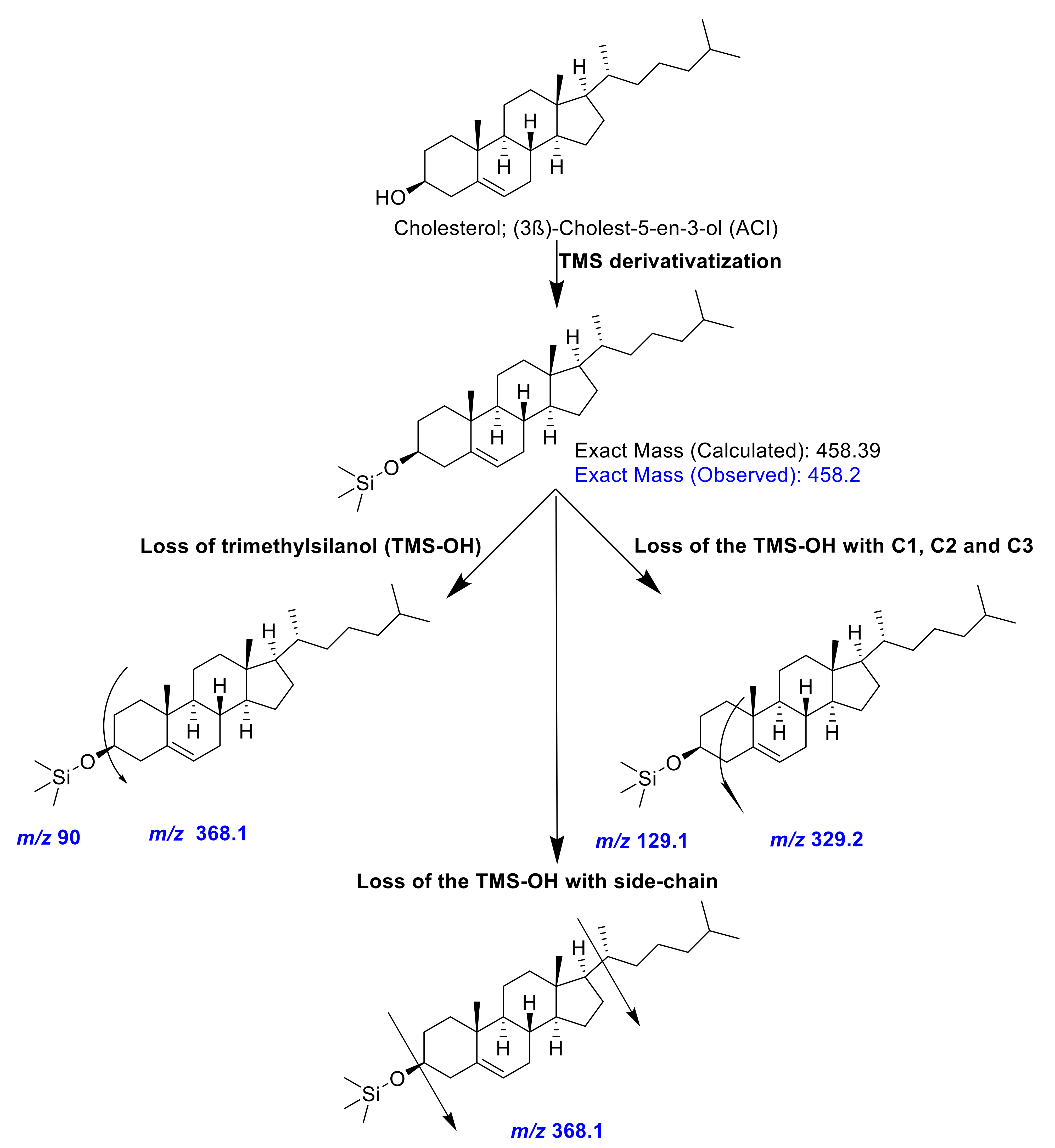
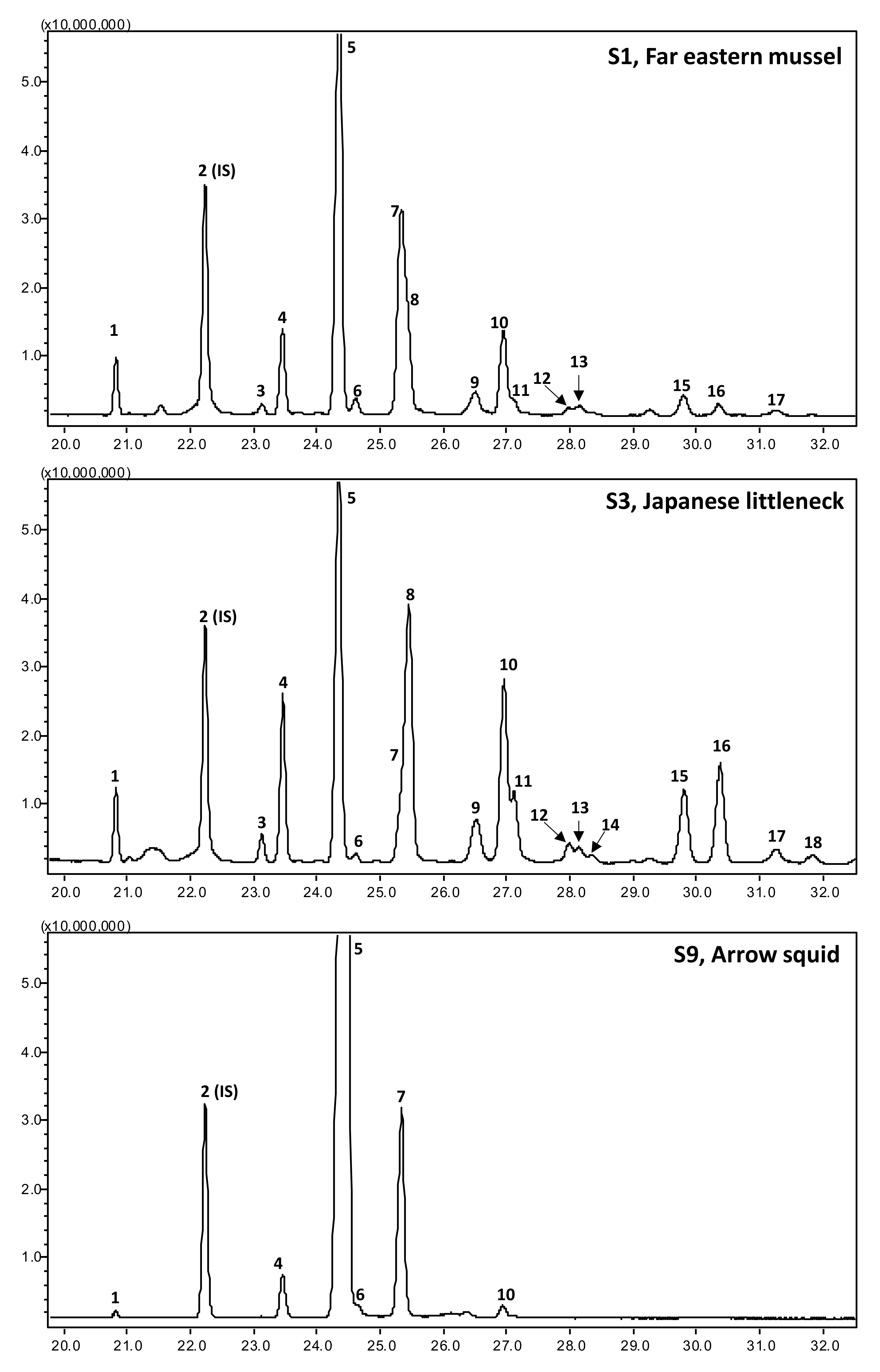
2.6. GC-MS Analysis of Sterols
Sterols were analyzed after silylation, utilizing a QP2010 SE GC-MS equipped with a fused silica Rxi-5ms column (30 m, 0.5-μm film thickness, 0.25-mm ID; Restek Corporation, Bellefonte, PA, USA). Helium was used as a carrier gas, maintained at the pressure control flow of 33.5 cm/min (7.8-mL/min total flow). The injector and MS ion source were precisely maintained at 260, while the MS interface was maintained at 280 °C. The column oven temperature was kept at 120 °C for 1 min, then progressively increased to 300 °C with a linear increase of 15 °C/min, and held at 300 °C for 27 min [
31]. One µL of samples and standards were injected in a 1:5 split ratio. Their fragmentation pattern was compared with authentic standards and reference databases (NIST08, NIST08S, and Wiley9).
2.7. Total Equivalent Antioxidant Capacities (TEAC)
The total equivalent antioxidant capacities (TEAC) of the lipids extracted from the shellfish species was determined using 1,1-diphenyl-2-picrylhydrazyl- (DPPH
•) and 2,2′-azino-bis(3-ethylbenzothiazoline-6-sulphonic acid)- (ABTS
•+) scavenging assays following our previously optimized procedures [
32], originally based on the studies of Thaipong et al. [
33] with minor modifications.
Briefly, for the ABTS+• decolorization assay, 1950 µL of freshly prepared ABTS+• solution was allowed to react with 50 µL (1.5 mg) of extracted lipids for 2 h in the dark. Later, the absorbance was measured at 734 nm, using a spectrophotometer (UV-2550, Shimadzu, Japan). A linear standard calibration curve was prepared using Trolox as standard in the range of 0.01–10 µg/mL. The results are expressed as the mg-of-Trolox equivalents (TE)/g of the lipids.
For the DPPH•-scavenging activities, 1950 µL of freshly prepared DPPH• solution (0.1 mM) was allowed to react with 50 µL (1.5 mg) of extracted lipids for 80 min in the dark. Later, the absorbance was measured at 517 nm using the spectrophotometer. A linear standard calibration curve was prepared using Trolox as standard in the range of 0.01–10 µg/mL. The results are expressed as the mg-of-Trolox equivalents (TE)/g of the lipids.
2.8. Cytotoxicity Studies
The HCT116 (Colorectal carcinoma), A2058 (Melanoma), A549 (lung carcinoma), T98G (Glioblastoma multiforme), and Hela (Adenocarcinoma) cell lines were purchased from the American Type Culture Collection (ATCC, Manassas, VI, USA). Cells were cultured in McCoy’s 5A Medium, Dulbecco’s Modified Eagle’s Medium, or Minimum Essential Medium Eagle, supplemented with 10% FBS, 1 mM sodium pyruvate, and 100 units/mL penicillin/streptomycin, accordingly, and incubated in a 5%-CO2 incubator at 37 °C. Prior to treatment, cells were seeded at 3 × 103 cells/per well density in 96-well plate and incubated overnight. Treatments were performed by carefully washing the cells with Dubelcco’s Phosphate Buffered Saline twice and adding lipids in media without FBS to the washed wells. Lipids were dissolved at a concentration of 20 mg/mL in DMSO, and cells were treated at concentrations ranging from 10 to 100 µg/mL in 0.5% DMSO for 24 h. Cytotoxicity was assayed by adding 10 µL of WST-1 EZ-cytox (DoGen, Suwon, Korea) per well with 90 µL of media without FBS and incubated at 37 °C for 40 min. The absorbance at 450 nm and reference wavelength at 600 nm were measured using a Microplate Spectrophotometer (Biotek, VT, USA). Experiments were performed in three technical repeats for more than three biological repeats. The most representative biological repeats were used to calculated IC50 concentration by mean of the AAT-Bioquest® online tool.
2.9. Statistical Analysis and Quality Control
A total of three replicate extraction and analysis were performed for each shellfish species. The results were analyzed using IBM SPSS statistics (version 25; IBM Corp., Armonk, NY, USA) employing a one-way analysis of variance (ANOVA), considering a significance level of 0.05 (Turkey HSD).
The use of the GC-MS method for quantifying sterols was recently validated in terms of accuracy, linearity, precision, and stability [
26]. The recovery of sterols was precisely monitored and normalized using 5β-cholestan-3α-ol as an internal standard.
4. Conclusions
The results of the present investigation indicated that the contents and composition of sterols, fatty acids, and tocopherols varied significantly among shellfish species. The highest n-3 PUFAs were recorded from arrow squid and pacific oysters, accounting for 53.2% and 53.0% of total fatty acids, respectively. However, the highest cholesterol content was recorded in arrow squid (154.4 mg/100 g; 92.6% of total sterols). In contrast, in the Japanese littleneck, Yesso scallop, and common orient clam, cholesterol was just 17.1%, 18.3%, and 18.9% of total sterols, respectively, which makes them the richest source of NCSs. The fat-quality indices indicated that lipids obtained from the Yesso scallop and Gazmi crab are least hypercholesteremic, atherogenic, and thrombogenic.
Lipids extracted from shellfish species showed ABTS+•-and DPPH+•-scavenging activities. The highest ABTS+•-scavenging activities were recorded from the far eastern mussel, while the lipids from the Gazami crab showed the highest DPPH+•-scavenging activities.
In the cytotoxic studies, lipids extracted from shellfish species showed varied levels of cytotoxicity against the studied cancer cells. The lipids extracted from the Argentine red shrimp showed the highest cytotoxicity against glioblastoma multiforme (T98G) cells, with an IC50 of 12.3 µg/mL. In contrast, lipids of the long arm octopus were the most cytotoxic to lung carcinoma (AS49) cells, with an IC50 of 13.3 µg/mL.
The composition of major lipophilic compounds and cytotoxicity data against several cancer types of cancer cells reported herein may be helpful in exploring the nutritional and anticancer potential of shellfish species.
The presence of a wide range of sterols (including isomeric forms) in shellfish species makes the qualitative analysis complicated. In the future, investigation with the aid of NMR could provide further information to confirm the identities. Moreover, sampling from several locations of the country can provide more comprehensive information on the contents of these nutritionally vital lipophilic constituents and their anticancer potential.


 ,
, 










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}