1. Introduction
Human movements are planned and executed by perceiving and integrating visual, somatosensory, and auditory sensory inputs [
1,
2]. The two-component, multiple process model of limb control proposes that effective limb control for goal-directed aiming movements includes integration of expected and actual sensory inputs from multiple sensory inputs. These sensory inputs include expected sensory consequences that are based on sensory memories. The model is based on extensive research on the role of vision for controlling upper limb goal-directed movements, as vision is a dominant source of information for aiming movements [
3,
4,
5].
The visual inputs from spatially accurate information to the central nervous system are a rich source of sensory information for movement control, including details of the limb, environment, and targets. A substantial body of literature has delved into the manipulation or elimination of each of these three components of visual input to elucidate their specific roles in movement planning and execution [
6,
7]. Studies that have focused on visual feedback related to the environmental cues and movement background have revealed that peripheral vision predominantly supplies online visual information about a limb relative to the environment, including the limb speed and direction as it approaches the target [
7,
8,
9].
In contrast, visual feedback of the limb via central vision holds particular significance for regulating spatial positioning of the limb, especially as the limb approaches the target vicinity during aiming movements. Additionally, contingent on the time available for processing aiming movement tasks, as well as the number of practice trials, central vision can contribute to real-time (i.e., online) or offline control of movement direction [
9].
The role of target vision has been studied independently and in conjunction with visual feedback of the limb. Removing visual feedback of the target leads to the movement unfolding based on the sensory memory of the target’s location [
10,
11,
12,
13,
14]. This sensory representation can be used for online corrections if visual feedback of the limb is present [
13]. Abahnini and Poteau (1999) demonstrated that even brief exposure to vision of the limb relative to the target significantly influences the planning of limb-target regulation later in the movement [
8]. Based on the aforementioned literature and in conjunction with the multiple process model (see [
2] for the multiple process model of limb control), vision of the limb serves as a reference point for movement planning and a crucial source of sensory input for online corrections.
Somatosensory information, and more specifically proprioception, has a complementary role in the control of human movement by providing information about the location of the limb in the space as well as the relative location of limbs to each other [
15,
16,
17,
18]. Thus, it is important to understand how these two major sources of information are integrated for voluntary limb control, including how humans adapt to changing sensory inputs. It is suggested that a combination of proprioception and visual inputs are the main sources of information for limb-target regulation. Thus, effective limb control includes integration of expected and actual sensory inputs from multiple sensory sources [
2]. Knowledge about the individual and integrative role of different sensory input is crucial for rehabilitation programs when either of these sensory inputs are deficient.
Previously, we used induced paresthesia to interrupt somatosensory inputs while also manipulating visual information by removing target vision upon movement initiation [
19,
20]. In this previous work [
19], we exclusively manipulated target vision. Thus, participants could combine available limb vision with their sensory representation of the target location. The presence of limb vision can allow for limb-target regulation based on the memory of the target position. Furthermore, we explored the removal of target vision both with and without induced paresthesia to better understand the changes in motor control strategies during rapid and accurate movement execution while experiencing paresthesia. That study aimed to uncover how participants adapt their movement control strategies in the presence of induced paresthesia and vision of the limb. Our findings indicated that the absence of target vision coupled with paresthesia adversely affected motor performance accuracy and efficiency. When vision of the target was removed and paresthesia was induced, participants had significantly higher bias for errors towards the midline of their body early during practice when compared to the no-paresthesia condition, or when vision of the target was available with paresthesia. Together, this pattern of results indicates that vision of the target and intact proprioception both contribute to limb-target corrections. We also found that participants did adapt to the effects of induced paresthesia with increased practice by adjusting their movement strategy to rely more on pre-planned movements. The strategy of pre-planning their movements included reducing their initial movement impulses and the need for online corrections to counter augmented neuromuscular noise.
While the strategy of pre-planning movements when experiencing induced paresthesia was successful when vision of the limb was still available, it is unclear if this strategy will still be effective if vision of the limb is also removed. The current study serves as a continuation of our prior research and introduces novelty by eliminating vision of the moving limb and surrounding environment, thereby accentuating the significance of afferent somatosensory input from the limb for successful target aiming movements. The overall aim was to assess if and how participants could adapt to the altered somatosensory sensory input when vision target and the limb were removed at movement initiation. In particular, if participants would be able to pre-plan their movement with disrupted somatosensory, target, and limb visual inputs. Two experiments were conducted to explore how humans adapt to the altered sensory input of the limb. In Experiment 1, we examined the implications of eliminating visual input of the entire visual environment (including the moving limb and target) combined with the introduction of temporarily induced paresthesia. This experimental setup effectively emulated the conditions someone with paresthesia might encounter while moving in a dark environment. The primary aim of this manipulation was to discern whether visual input, specifically pertaining to the limb, plays a pivotal role in enhancing movement performance when participants are exposed to induced paresthesia. We hypothesized that removing visual input from the entire visual environment would lead to larger and more pronounced differences between conditions with and without visual feedback, as compared to our previous study [
19]. This hypothesis stemmed from the understanding that removing limb-specific visual cues would prevent using limb-specific visual feedback to compensate for the induced paresthesia. However, we did expect proprioception information would be used to achieve some degree of online control when vision was removed [
2,
21]. Consequently, the presence of induced paresthesia was expected to disrupt these online control processes reliant on proprioception. Additionally, a comparison between early and late movement trials was conducted to probe participants’ adaptability to the somatosensory manipulation. Thus, a secondary objective of Experiment 1 was to investigate the changes in motor control strategies to these sensory manipulations.
Next, Experiment 2 was conducted to evaluate if augmented auditory feedback valuable to assist participants in updating their motor control strategies when experiencing disruptions in visual (target, limb, and environment) and somatosensory feedback. The auditory feedback provided participants with confirmation that their reaching had successfully acquired the target. Thus, the fundamental goal of Experiment 2 was to assess whether terminal auditory feedback could improve the development of accurate and efficient movements. We predicted participants would benefit from the addition of auditory feedback and would show reduced dependence on auditory versus visual feedback because auditory feedback is not the preferred modality [
22,
23].
3. Experiment 2
When paresthesia was induced in Experiment 1, participants were unable to update their movement strategies to account for the altered proprioceptive input. Disruptions in somatosensory input due to injury and disease are a common experience. Experiment 1 provided additional evidence that these changes impact the performance of the types of goal-directed aiming movements that are used daily when interacting with touchscreen technology. Therefore, understanding how to facilitate performance when somatosensory input is disrupted has both theoretical and practical implications. A growing literature in both neurotypical and neurodiverse populations have provided evidence that augmented auditory feedback can facilitate movement performance [
37,
38,
39]. When provided at or near target acquisition, augmented auditory likely works by providing confirmation to participants that they successfully acquired the target. Thus, the purpose of Experiment 2 was to assess if it is possible to supplement for the disrupted somatosensory feedback through providing this type of augmented auditory feedback at target acquisition.
We predicted that the addition of an auditory tone would facilitate improvements in movement efficiency when visual feedback was not available. In other words, the benefit of the auditory feedback would be greater when both visual and somatosensory feedback have been disrupted. However, when two sources of accurate feedback are available (i.e., vision and audition) then movements will be performed more accurately compared to when only one source of accurate feedback is available.
3.1. Methods
3.1.1. Participants
Fourteen healthy young adults (5 females, 9 males) with mean age of 22.7 ± 2.9 years participated in Experiment 2. Consistent with Experiment 1, participants had no neurological condition or orthopedic injury that would interfere with their performance of the task. All experimental procedures were consistent with the Declaration of Helsinki and were approved by the local ethics board. All participants provided signed informed consent prior to their participation in the experiment and received a base honorarium for their time.
3.1.2. Apparatus, Materials, Design, Procedure, and Analysis
The overall experimental set-up and design of Experiment 2 was identical to Experiment 1, with one difference: an auditory tone was introduced, sounding only when participants successfully landed on the target. The beep was presented immediately at target acquisition (i.e., when participants touched the target). This auditory feedback complemented the information about movement time and whether they hit or missed the target. The latter two types of feedback were the only ones provided in Experiment 1. Consistent with Experiment 1, vision and induced paresthesia were blocked and counterbalanced across participants. Thus, the experimental design remained the same, with the addition of auditory feedback for successful trials in all four conditions. The individual trial sequence also remained the same, with one exception. After participants completed each movement, an auditory tone was presented via standard computer speakers when that specific movement was accurate. The data analysis also followed the procedures outlined in Experiment 1. Please see Experiment 1 for details of the visual and somatosensory conditions.
3.2. Results
3.2.1. Monofilament Test
As in Experiment 1, baseline monofilament test results were analyzed and compared with post-stimulation using the Wilcoxon signed-rank test. The findings revealed that participants sensed thicker filaments after the application of stimulation, in contrast to their baseline measurements (Z = −3.329,
p = 0.001,
Table 2).
3.2.2. Temporal Measurements
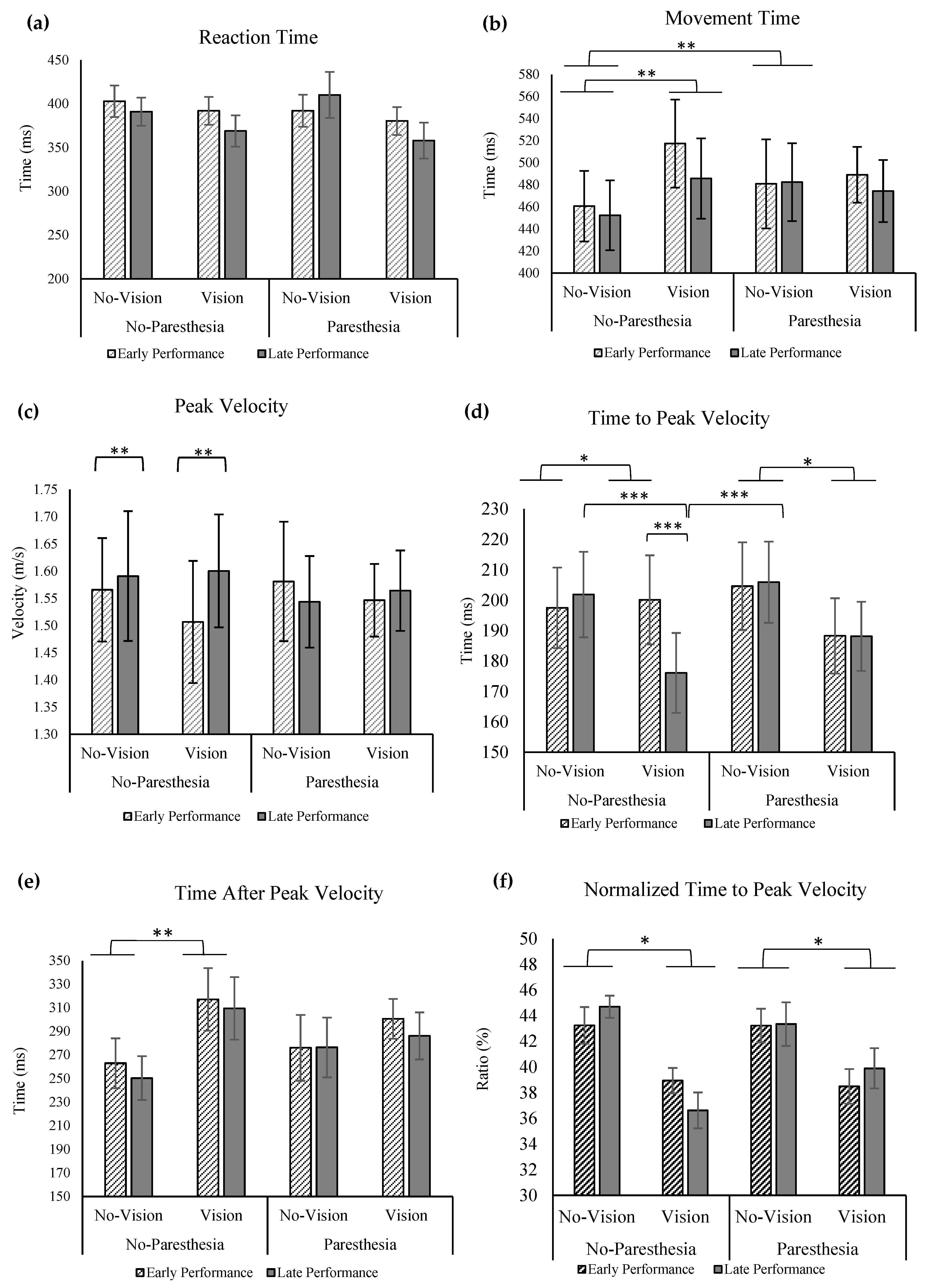
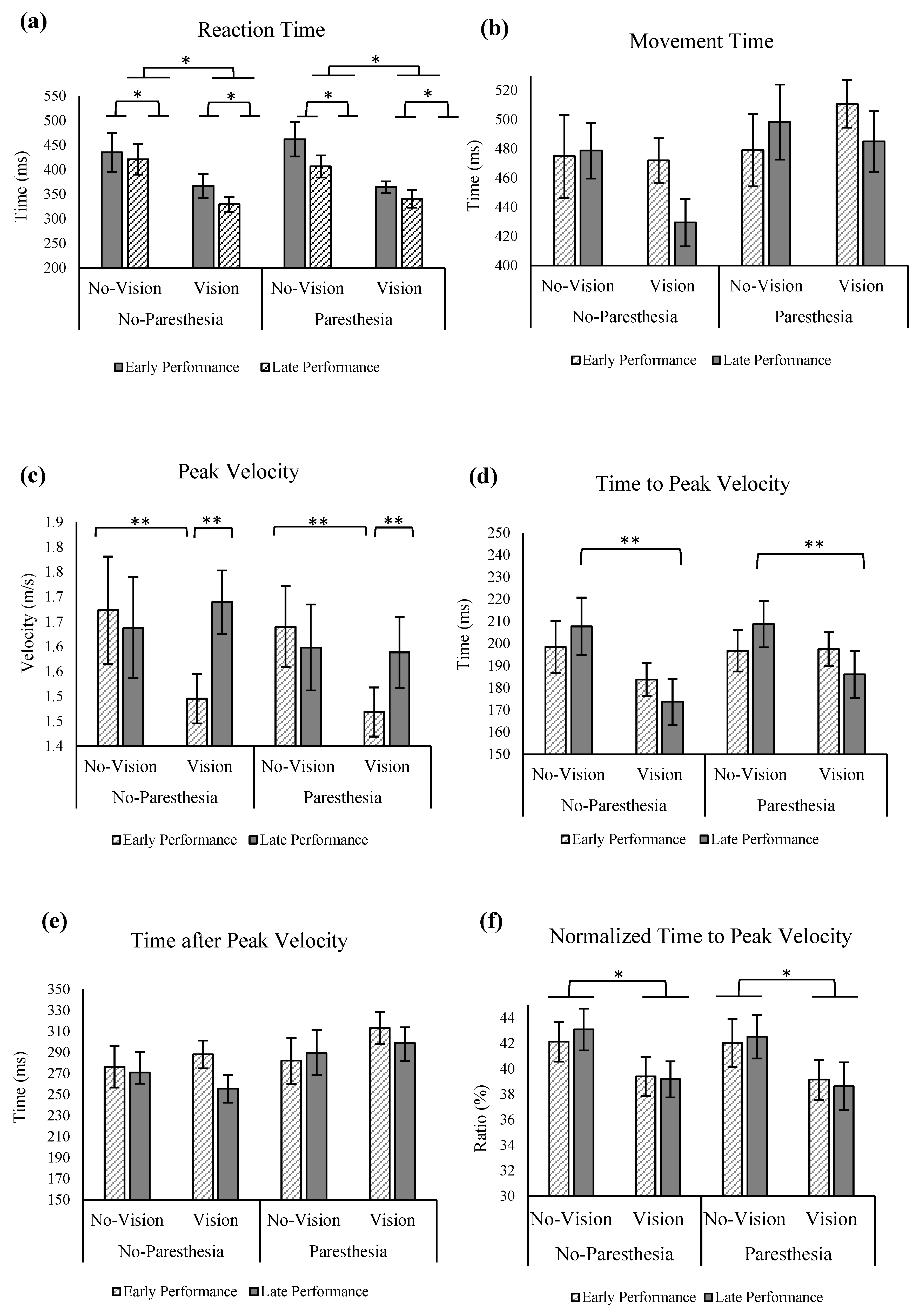
No significant main effects or interactions were found for the factors of vision, practice, or paresthesia for MT (
Figure 5a). For the outcome of RT, significant main effects were found for vision (F (1,13) = 12.936;
p = 0.003, ηp
2 = 0.499) and practice (F (1,13) = 16.283;
p = 0.001, ηp
2 = 0.556,
Figure 5b). Reaction times were found to be significantly longer in the no-vision condition compared to the vision condition, as well as during early performance compared to late performance.
Regarding peak velocity (PV), a significant interaction between practice and vision was observed, F (1,13) = 7.307,
p = 0.018, ηp
2 = 0.36. Tukey’s HSD analysis revealed that while PV was significantly lower with full vision compared with no vision in early performance, with more practice in the late performance trials, PV with full vision increased and became closer to the PV value in the no vision in late performance (
Figure 5c). Statistical analysis for time to peak velocity (ttPV) revealed a significant main effect for vision: ttPV was significantly longer when vision was removed, F (1,13) = 6.234;
p = 0.027, ηp
2 = 0.324. Also, there was significant interact ion of factors of vision and practice (F (1,13) = 6.388;
p = 0.025, ηp
2 = 0.329). Tukey’s HSD test showed that only in the late performance with full vision did participants have significantly shorter ttPV compared to when vision was removed; however, the ttPV in these two vision conditions was not significantly different in the early performance (
Figure 5d).
No significant main effects or interactions were found for the factors of vision, practice, or paresthesia for time after peak velocity (taPV;
Figure 5e). When ttPV outcome was normalized with MT, statistical analysis showed a main effect of vision, F (1,13) = 6.364;
p = 0.025, ηp
2 = 0.329; that is, participants spent a larger percentage of their movement time before PV when vision was removed compared to a full-vision condition (
Figure 5f).
3.2.3. Spatial Measurements
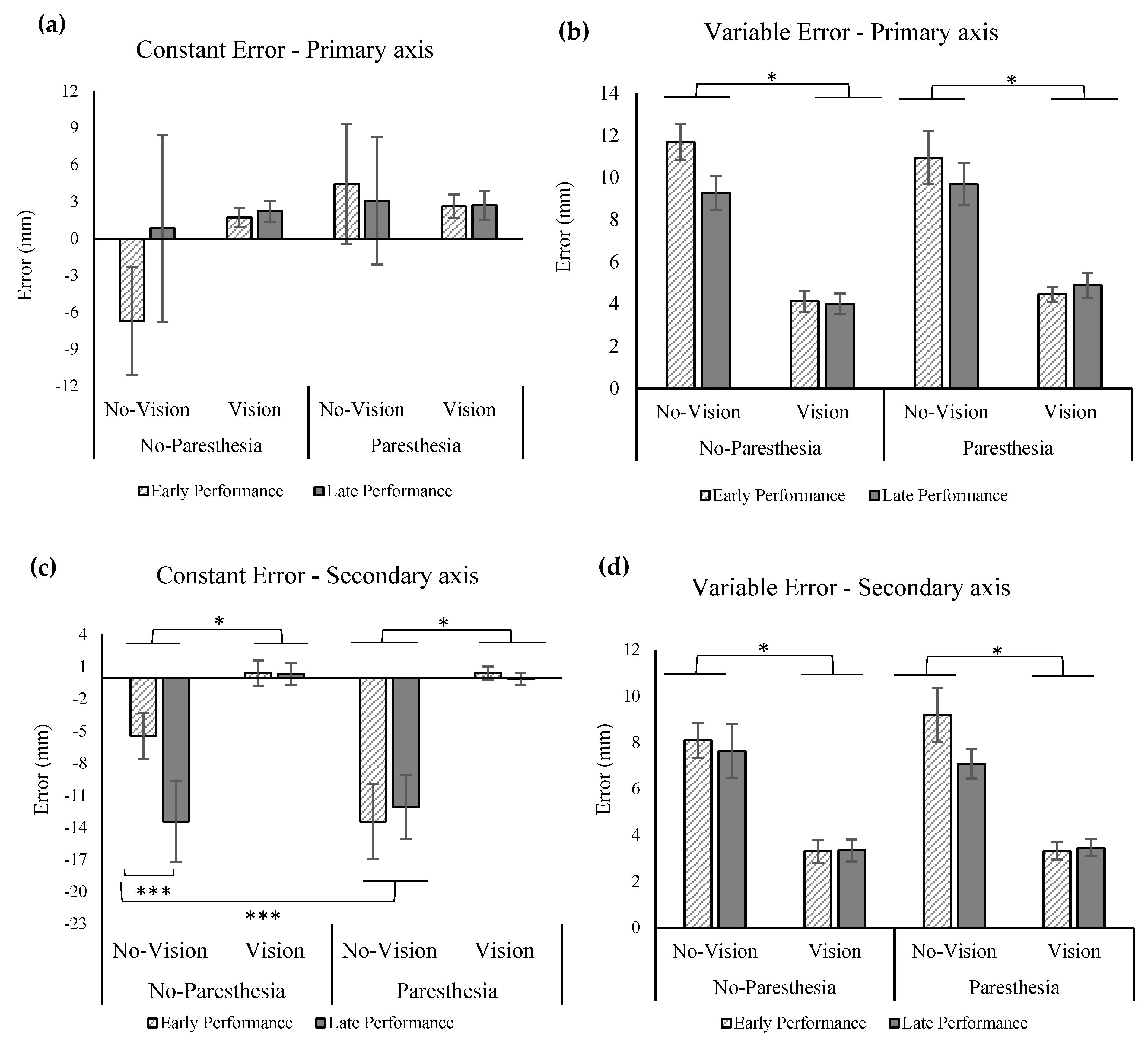
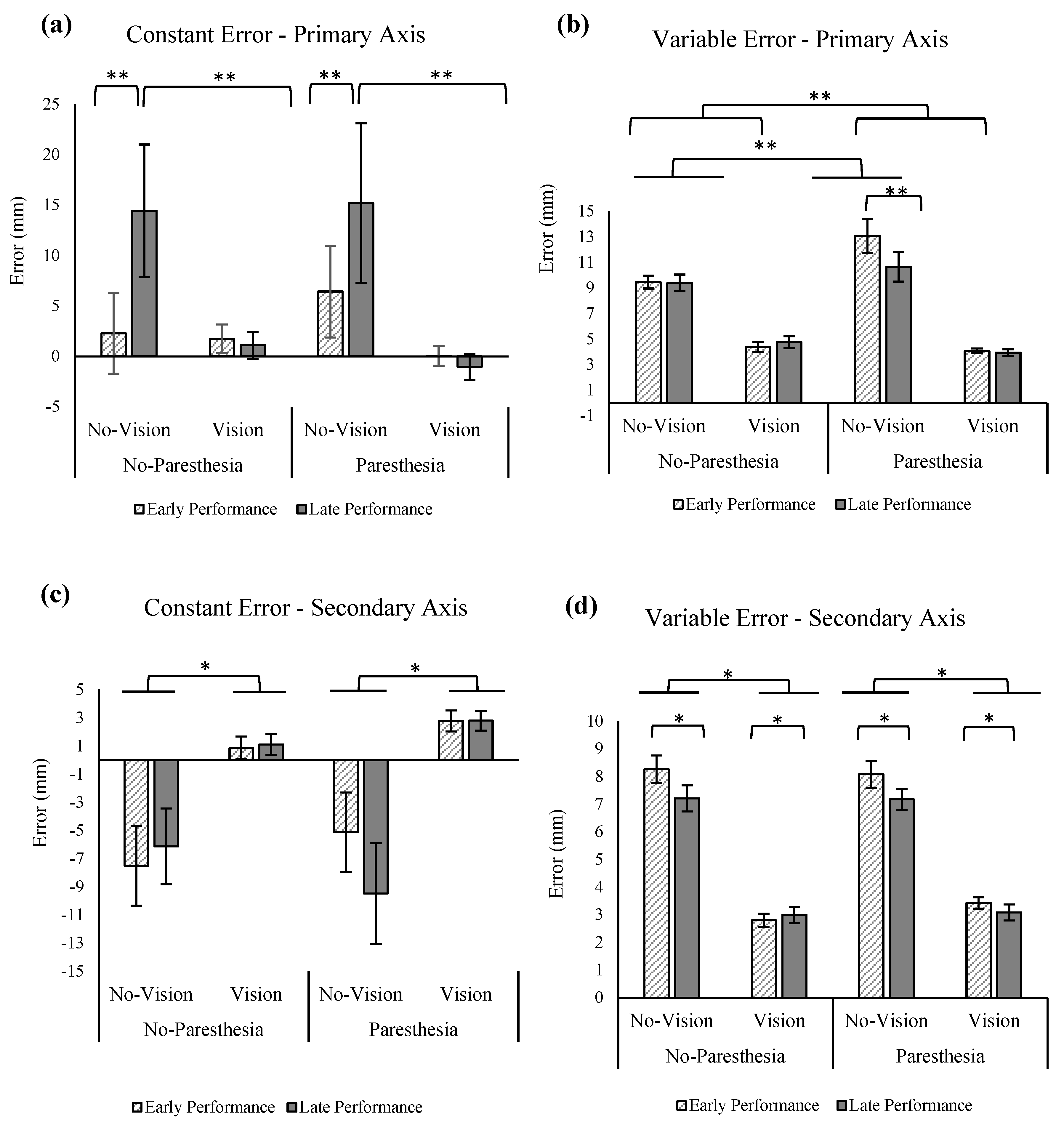
The ANOVA for the outcome of CE in the primary axis of movement showed a significant main effect for practice, F (1,13) = 4.713,
p = 0.049, ηp2 = 0.266, as well as a significant interaction for the factors of vision and practice, F (1,13) = 5.715,
p = 0.033, ηp
2 = 0.305 (
Figure 6a). Tukey’s HSD showed significantly larger overshoots for the no-vision condition when compared with the vision condition in the late performance. However, the CE in the primary axis in these two conditions was not significantly different in the early performance. Also, according to Tukey’s HSD, comparisons of late versus early performance in the no-vision conditions found a significant increase in CE late in performance. Results of the statistical analysis for VE showed significant main effect for vision, F (1,13) = 133.209,
p = 0.000, ηp
2 = 0.911 (
Figure 6b) and interaction of vision and paresthesia, F (1,13) = 5.308,
p = 0.038, ηp
2 = 0.290, as well as an interaction between paresthesia and practice (F (1,13) = 6.841;
p = 0.021, ηp
2 = 0.345). Tukey’s HSD for the interaction of vision and paresthesia showed significantly larger VE for the no-vision condition when compared with the full-vision condition in both with and without paresthesia conditions. Additionally, comparisons of with and without paresthesia trials when vision was blocked showed significant increase in VE when paresthesia was present. Post hoc testing using Tukey’s HSD for interaction of paresthesia and practice showed that participants had significantly larger VE in the presence of paresthesia when compared with the no-paresthesia condition only in the early performance; however, the VE in the primary axis in these two conditions was not significantly different in the late performance (
Figure 6b). Additionally, comparison of VE in the early and late performance of the paresthesia condition showed significantly higher VE in the early performance.
Consistent with Experiment 1, results of CE in the secondary axis showed significant main effect for vision, F (1,13) = 18.62,
p = 0.001, ηp
2 = 0.589, indicating larger undershoot errors when vision was removed (
Figure 6c). Results of VE in the secondary axis were similar to the primary axis; that is, there was a significant main effect for vision indicating more variability when vision was removed (F (1,13) = 232.338;
p = 0.000, ηp
2 = 0.947;
Figure 6d). Additionally, a main effect for practice was observed (F (1,13) = 6.711;
p = 0.022, ηp
2 = 0.340;
Figure 6d), indicating that participants exhibited smaller VE in the secondary axis with more practice.
3.2.4. Movement Trajectories
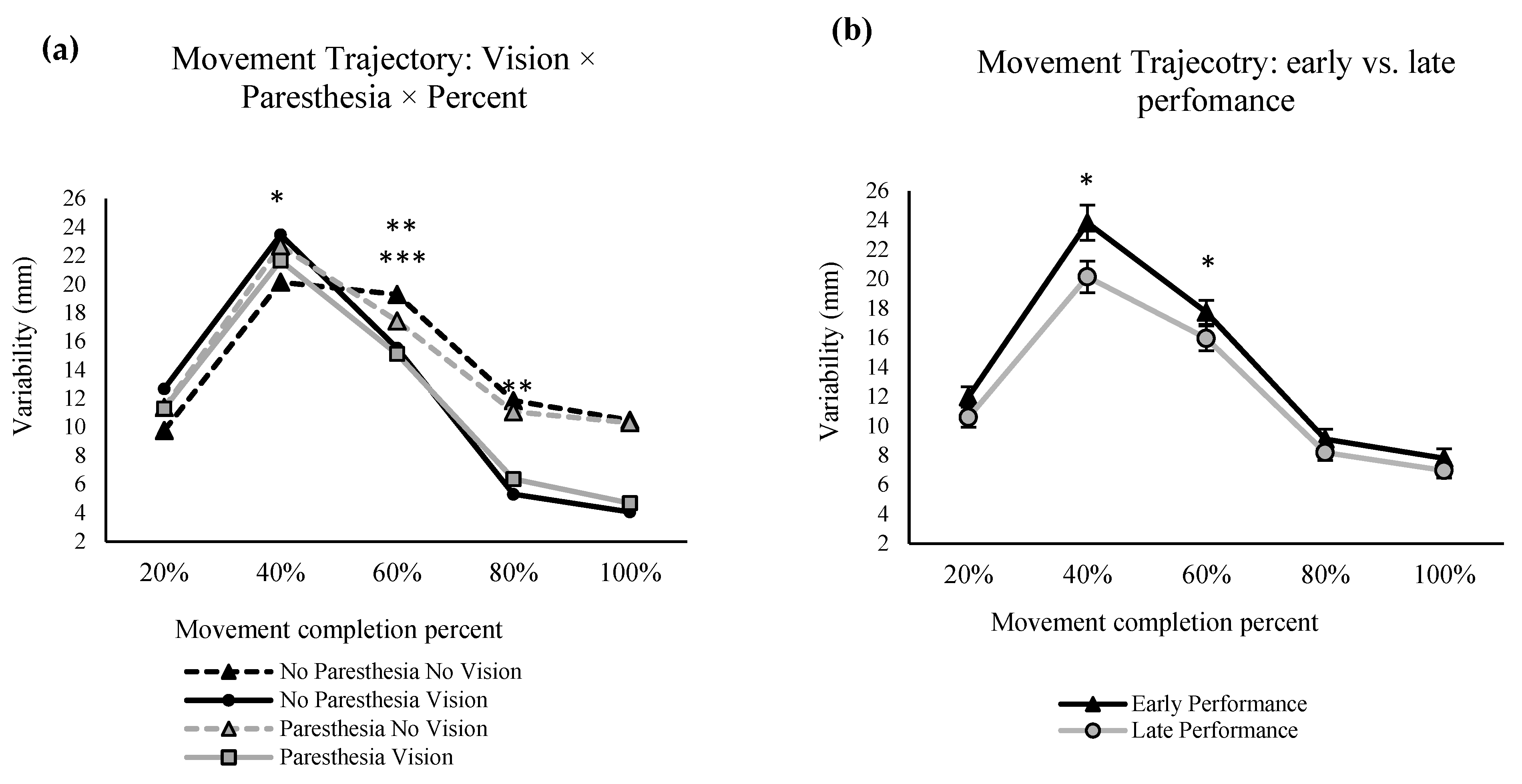
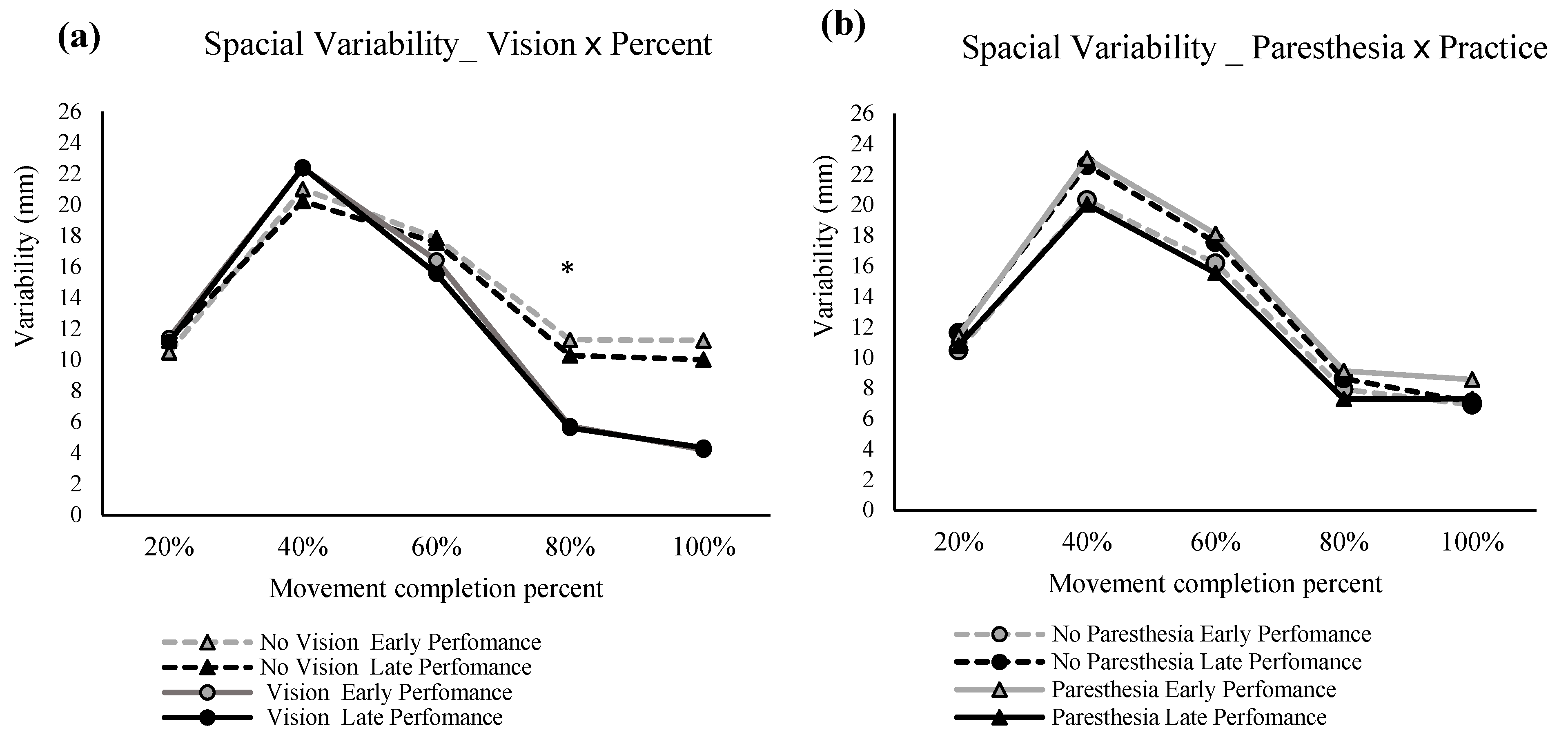
The repeated measures ANOVA for the spatial variability in the primary movement axis showed significant main effects for percentage of the movement, F (1.372,17.842) = 81.553,
p = 0.000, ηp
2 = 0.863, and vision F (1,13) = 6.840,
p = 0.021, ηp
2 = 0.345 (
Figure 7a). Also, there was a significant interaction between factors of percentage of movement and vision, F (1.824,23.712) = 16.316,
p = 0.000, ηp
2 = 0.557 (
Figure 7a), and another interaction between the paresthesia and practice, F (1,13) = 13.578,
p = 0.003, ηp
2 = 0.511 (
Figure 7b). Post hoc analysis for the interaction of percentage of movement and vision showed that at 80% of the movement time, no-vision conditions had significantly higher variability compared with the full-vision condition. Post hoc analysis for the interaction of practice and paresthesia showed that, in early performance, the presence of paresthesia significantly increased the spatial variability (
Figure 7b). Also, when paresthesia was present, trajectories were more variable in early performance when compared with the late performance.
3.3. Discussion
3.3.1. Effect of Sensory Manipulation on Movement Strategy
Participants in Experiment 2 adapted their RT, PV, and ttPV to accommodate the changing visual and somatosensory inputs. RT, PV, and ttPV are all measures that are associated with adjusting movement preparation [
2,
31,
40]. Specifically, with practice, participants increased their PV from early to late performance when vision was available. This increase in PV likely reflects a more confident movement plan that may also result in more efficient movements. That is, in early performance, participants reduced their PV to achieve more accuracy. As they practiced more, they improved their movement planning, as shown by shorter RTs combined with higher PV with vision and late performance [
41]. Moreover, consistent with Experiment 1, ttPV was significantly shorter than the ttPV in the no-vision condition. This pattern indicates that only when vision was available did more practice lead to more forceful movement initiation, which led to a higher PV and shorter ttPV. When vision was obstructed, participants reduced their PV to mitigate the spatial variability of their movements and improve the endpoint accuracy of their limb movements [
42]. More forceful movement initiation only when vision was available would be expected as participants could use visual feedback during movement execution to adjust for any increase in the variability of the initial movement impulse [
2,
41].
3.3.2. Effect of Sensory Manipulation on Endpoint Accuracy
The analysis of constant error in the primary axis showed an expected effect for vision, whereby target aiming was more accurate when vision was available. The improved accuracy was also achieved within the same movement time as trials without vision. When vision was removed at movement initiation, there was a clear effect on endpoint accuracy. However, somewhat unexpectedly, participants displayed larger overshoot errors in the primary axis with more practice when compared with their early performance. Overshoot errors are not typically reported in the literature; with increased practice without vision, one would typically expect participants to undershoot the target. It is possible that the relatively high index of difficulty combined with the auditory feedback, and the task instructions designed to motivate participants to challenge themselves to move as quickly and accurately as possible, combined to encourage participants to take on a more forceful movement strategy.
Consistent with the above explanation, the significant interaction for vision and paresthesia for VE in the primary axis showed that lack of visual input led to even larger VE in the primary axis when paresthesia was present. In addition, the interaction of practice and paresthesia in the VE for the primary axis showed that when compared with the no-paresthesia condition, induced paresthesia led to significantly larger VE early in performance; however, with practice, participants improved their performance to the VE of the no-paresthesia condition. The interaction of practice and paresthesia for the movement trajectories also showed a similar result. That is, when paresthesia was induced, then more practice led to significantly less spatial variability. However, this same pattern was not seen for the no-paresthesia condition. The lack of improvement with practice for the no-paresthesia group likely occurred because participants were accustomed to integrating proprioceptive input from the limb and therefore did not need to adjust their movement strategies in the no-paresthesia condition. Finally, the interaction of percentage of movement time and vision indicates that visual feedback was being used to improve endpoint accuracy in the vision conditions, as at 80 precent of movement time there was reduced spatial variability in the vision condition. This reduction in spatial variability late in the movement is reported to represent the implementation of online correction based on the available visual feedback.
3.3.3. Sensory Manipulation and Practice
With more practice, participants in Experiment 2 showed significantly higher PV only when vision was available (
Figure 5c). Also, when compared with no-vision conditions, ttPV became significantly shorter during late performance with vision (
Figure 5d). This pattern suggests a shift in movement control strategy and planning. That is, moving from primarily pre-planned movements to a greater reliance on online control mechanisms when participants had access to their intrinsic sensory inputs. In contrast, participants did not alter their movement control strategy in response to changes in somatosensory input (induced paresthesia). These findings are consistent with the outcomes of Experiment 1. In Experiment 2, there was a notable positive effect of practice as observed in the results of VE in the primary axis in the no-vision condition and in the presence of paresthesia (
Figure 6b); participants improved their accuracy by reducing their VE with more practice. This improvement in the endpoint variability could be attributed to the augmented auditory feedback in Experiment 2 (please see the general discussion for the direct comparison of the endpoint variability for the participants of the two experiments). Another effect of practice was seen in larger CE with more practice where in conditions without vision participants had more overshoot errors (
Figure 6a). Auditory stimuli have been reported to have an activating effect [
43] on movement performance, and therefore it is possible that the lack of visual feedback combined with auditory feedback led to more forceful movement impulses that resulted in target overshoots [
44].
4. General Discussion
4.1. Adaptability and Practice
In both experiments, participants benefited most from practice when their intrinsic feedback was unaltered. That is, for both experiments, participants improved their movement strategy throughout practice by using a shorter ttPV and higher PV in the no-paresthesia and full-vision condition specifically. Thus, changes to movement strategies with practice that helped participants achieve movements that were both fast and accurate occurred primarily with unaltered intrinsic sensory inputs. That said, we did find evidence that participants adjusted their movement strategies when either or both visual and somatosensory feedback were disrupted.
4.2. Paresthesia and Movement Strategy
Overall, participants updated their movement strategies to account for the sensory inputs that were available to them. The results of Experiment 1 indicate participants updated their movement strategy according to if vision of the target and moving limb was or was not present, but only in the no-paresthesia condition. In contrast, when paresthesia was induced, participants did not adjust their movement time or time after peak velocity movement strategies. The lack of changes to MT and taPV when paresthesia was induced suggests that participants were not able to make use of online corrections to their limb trajectories as they normally would. Once again, this highlights the multisensory nature of these online corrections in that the corrections require both visual and proprioceptive inputs [
2].
Based on the findings of Experiment 1, the combination of induced paresthesia and no visual feedback led to a significant movement bias towards the body midline. The findings also provide evidence for the idea that the contribution of proprioceptive input for movement accuracy is indeed larger in the absence of visual feedback. Specifically, the bias was greater with paresthesia when vision was removed. With intact sensory processing (i.e., without paresthesia and with full vision), participants had the longest MTs. In conjunction with improved endpoint accuracy, this pattern of results indicates that participants used the available sensory information and spent more time implementing online corrections to the limb trajectory. Notably, vision in the presence of induced paresthesia did not lead to longer MTs, presumably because the participants had less sensory feedback available to use for online movement corrections [
2]. The longest MT in the intact sensory input condition was associated with the shortest time to peak velocity, especially with more practice. In the condition without vision and with paresthesia, the longer movement time (
Figure 2b), accompanied by a marginal reduction in movement trajectory variability after 40% of the movement (which corresponds to approximately PV, and the onset of the limb-target correction phase;
Figure 4a), as compared to the condition without vision and without paresthesia, could be explained by the principles of the optimized sub-movement model [
42,
45,
46]. That is, due to the absence of availability of the two major sources of sensory input (visual and proprioceptive feedback), participants had to exchange a fast and forceful movement, which would lead to larger movement trajectory variability, for a more controlled movement, resulting in overall slower movements with less variability in movement trajectory. This choice of movement strategy fits with a ‘play it safe’ strategy and could account for the observed slower movements with longer movement times. In summary, the temporal movement findings are all in agreement that with full sensory information available and more practice, participants developed a new movement strategy. The differences in the temporal parameters in paresthesia versus no paresthesia conditions with full vision available signifies the role of proprioception for informing both the movement strategy and efficacy, even when visual information of the limb, target, and environment is available.
Experiment 2 added augmented auditory feedback upon target acquisition. When auditory feedback was present then participants updated how they planned their movements (as seen through changes in RT, with a lack of any changes taPV) [
23,
37]. Changes in the time needed to initiate the movement (RT) as well as to execute the initial movement impulse (ttPV) are evidence of more efficient movement planning. Consistent with previous literature [
47,
48], it appears that the auditory feedback allowed participants to complete each movement more efficiently, freeing up processing resources to plan the next movement more effectively and efficiently. Further evidence of the improved planning includes the reduced trial-to-trial VE when paresthesia was induced and vision of the movement environment was removed.
4.3. Paresthesia and Movement Accuracy
The preceding section elaborated on the evidence found in both Experiments 1 and 2 for the significant contribution of proprioceptive input to the movement planning and strategies adopted by participants of both experiments. However, a similar effect was not found for movement accuracy. Specifically, the presence of visual feedback could compensate for the sensory feedback deficiencies caused by induced paresthesia for limb-target regulation after about 60% of the total movement time. This finding is supported by the results of the outcomes of CE (especially in the secondary axis) and VE (in both primary and secondary axes), which showed that vision was the most important source of sensory feedback for endpoint accuracy (
Figure 3 and
Figure 6). Furthermore, movement trajectory variability findings (
Figure 4 and
Figure 7) also displayed less variability after 60% into the movement in the full vision conditions, regardless of paresthesia. These findings are consistent with the literature in signifying the role of visual feedback as the dominant and most reliable source of spatial information for movements such as goal-directed aiming tasks [
2,
3,
5], especially when participants are aware that visual information will be available to them [
2,
31]. The results of the present experiments demonstrate the key contribution of vision of the moving limb. Specifically, participants had difficulty updating their motor control strategies when both somatosensory and visual inputs were disrupted. In the context of models of limb control, such as the two-component multiple process model, this finding demonstrates that a minimum amount of (unaltered) sensory input is required for participants to update their movement strategies using pre-existing internal models of their limb movements.
4.4. Auditory Feedback and Movement Strategy
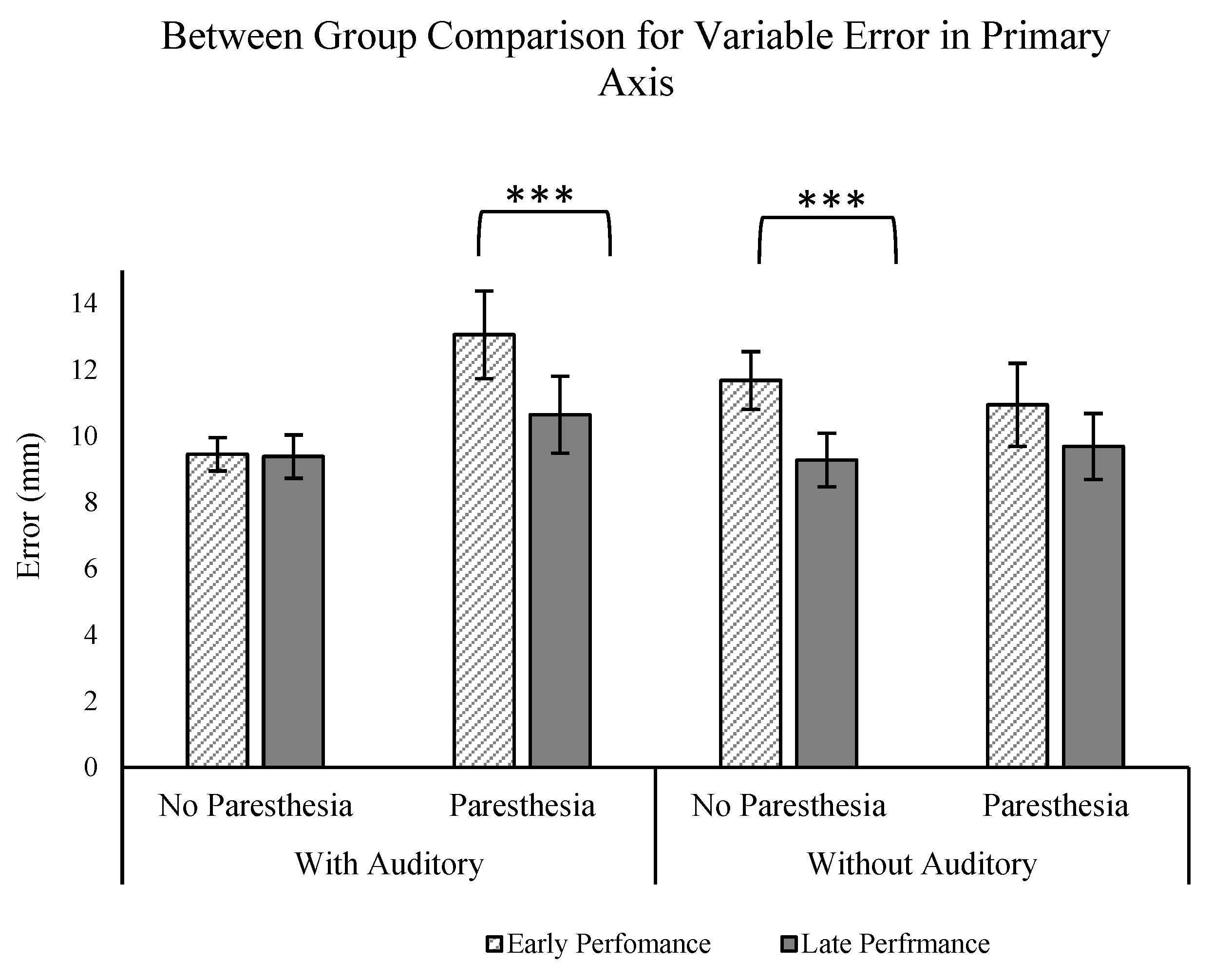
To further explore the influence of auditory feedback on movement performance and strategy under conditions of disrupted visual and somatosensory feedback, we conducted a mixed-design ANOVA (2 groups (auditory vs. no auditory) × 2 paresthesia (paresthesia vs. no paresthesia) × 2 practice (early vs. late performance)). We focused on comparison of performance outcomes for participants of Experiment 1 and Experiment 2, exclusively comparing conditions in which visual feedback was obstructed (no-vision conditions).
Our analysis centered on specific performance metrics aimed at assessing the impact of auditory feedback on both movement accuracy (CE, VE) and strategy (ttPV/MT, MT). The only significant finding was for the VE in the primary axis. The ANOVA analysis revealed a significant three-way interaction for the factors of paresthesia, group, and practice (F (1,24) = 5.547;
p = 0.027, ηp
2 = 0.188;
Figure 8). Interestingly, the post hoc analyses revealed that only for the group who received auditory feedback was there a significant effect of practice (comparison of early vs. late trials) with paresthesia. That is, with paresthesia, participants had less VE with more practice only when auditory feedback was provided, while the same practice effect was not found with paresthesia in the group without auditory feedback. Also, the group without auditory feedback exhibited a significant practice effect (characterized by decreased endpoint variability) when paresthesia was absent, which was not the case for the group that received auditory feedback. This absence of practice effect for the group who received auditory feedback could be attributed to a potential ceiling effect influenced by the presence of auditory feedback.
Taken together, the present experiments provide further evidence that humans can improve movement consistency with augmented auditory feedback, which is especially relevant when intrinsic feedback is most disrupted. Notably, the present experiment extends previous findings from clinical populations to an experimental model of induced paresthesia. That is, when all other aspects of movement control remain typical, the temporary disruptions of somatosensory feedback led to identifiable differences in movement performance and the movement strategies used to achieve that performance. Consistent with previous research with clinical populations, improvements in movement performance with the addition of auditory feedback were seen through changes in movement planning. Interestingly, the strategy of adapting to induced paresthesia through changes in movement planning was also seen in our previous research where vision of the target was removed, but vision of the limb was still available. In the future, it will be interesting to assess if additional differences are evident if participants have more time to adapt to the induced paresthesia.
It should be noted that another way to interpret the combined analysis is that auditory feedback supplemented for practice through a reduction in endpoint variability (but no change in other movement parameters). It has been proposed that augmented auditory feedback can free up the central nervous system for other processing—namely, improvements in movement planning for the subsequent movement [
23,
48,
49]. Practice is another means by which sensory processing becomes more efficient, and these changes were seen through in the present experiment through more efficient taPV and improved endpoint accuracy with practice.
6. Conclusions
We report that temporarily induced paresthesia influenced visuomotor control in an otherwise neurotypical adult group of participants. This finding is consistent with humans using proprioception of their current limb position to update movement control based on current and expected sensory consequences. When no auditory feedback was presented, then changes were observed primarily during movement execution (i.e., MT and time after PV), suggesting that participants were trying to adjust movement control by adapting parameters of their movement execution. In contrast, when auditory feedback of target acquisition was presented in Experiment 2, then participants instead updated how they planned through movements through changes in RT and time to PV.
The present experiments provide additional evidence that humans are able to flexibly adapt their movement strategies according to the presence and quality of different sources of intrinsic and augmented sensory feedback. Specifically, how humans adapt their approach to movement control depends on the combination of intrinsic and augmented feedback available. We report that participants updated their approach to movement planning or execution in specific ways that allowed them to achieve the task goals according to the sensory conditions. For example, through changing movement plans and/or taking time to update the movement trajectory based on available somatosensory and/or visual feedback. In other words, how humans change their movement strategy depends on the available sources of sensory input. The somatosensory manipulation used in the present experiments is relevant to various clinical populations who experience sensations such as paresthesia in the upper limb, including individuals with conditions such as stroke, spinal cord injury, or diabetes. The findings of this study can contribute to our understanding of how humans compensate for disrupted sensory inputs as well as how augmented auditory feedback may help to compensate for these disruptions.












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}