
Behavioral and Electrocortical Response to a Sensorimotor Conflict in Individuals with Fibromyalgia
 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants and Ethical Statement
2.2. Study Design
2.3. Clinical Status and Tactile Acuity
2.4. Instrumentation and Stimuli
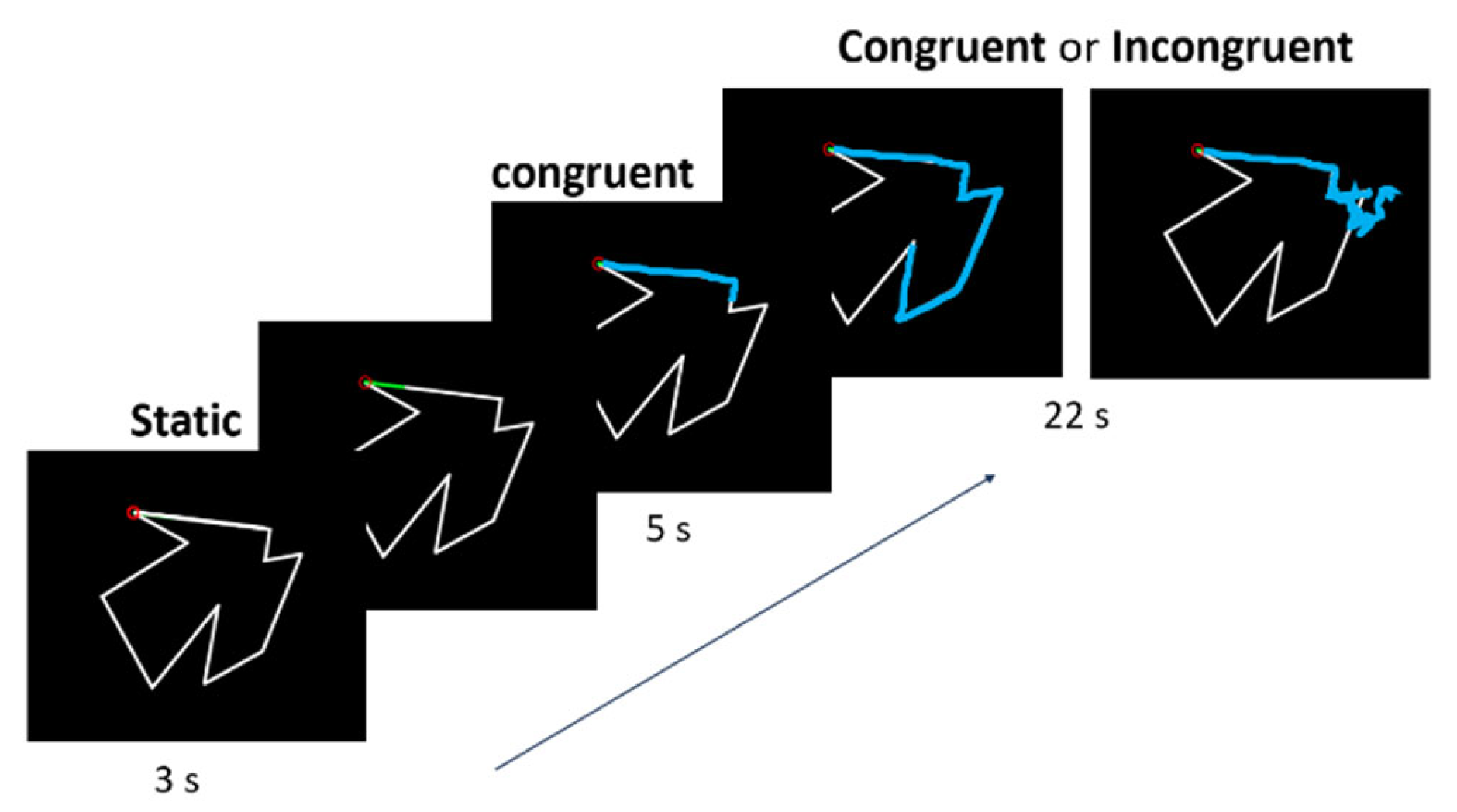
2.5. Experimental Task
2.5.1. Behavioral Measures
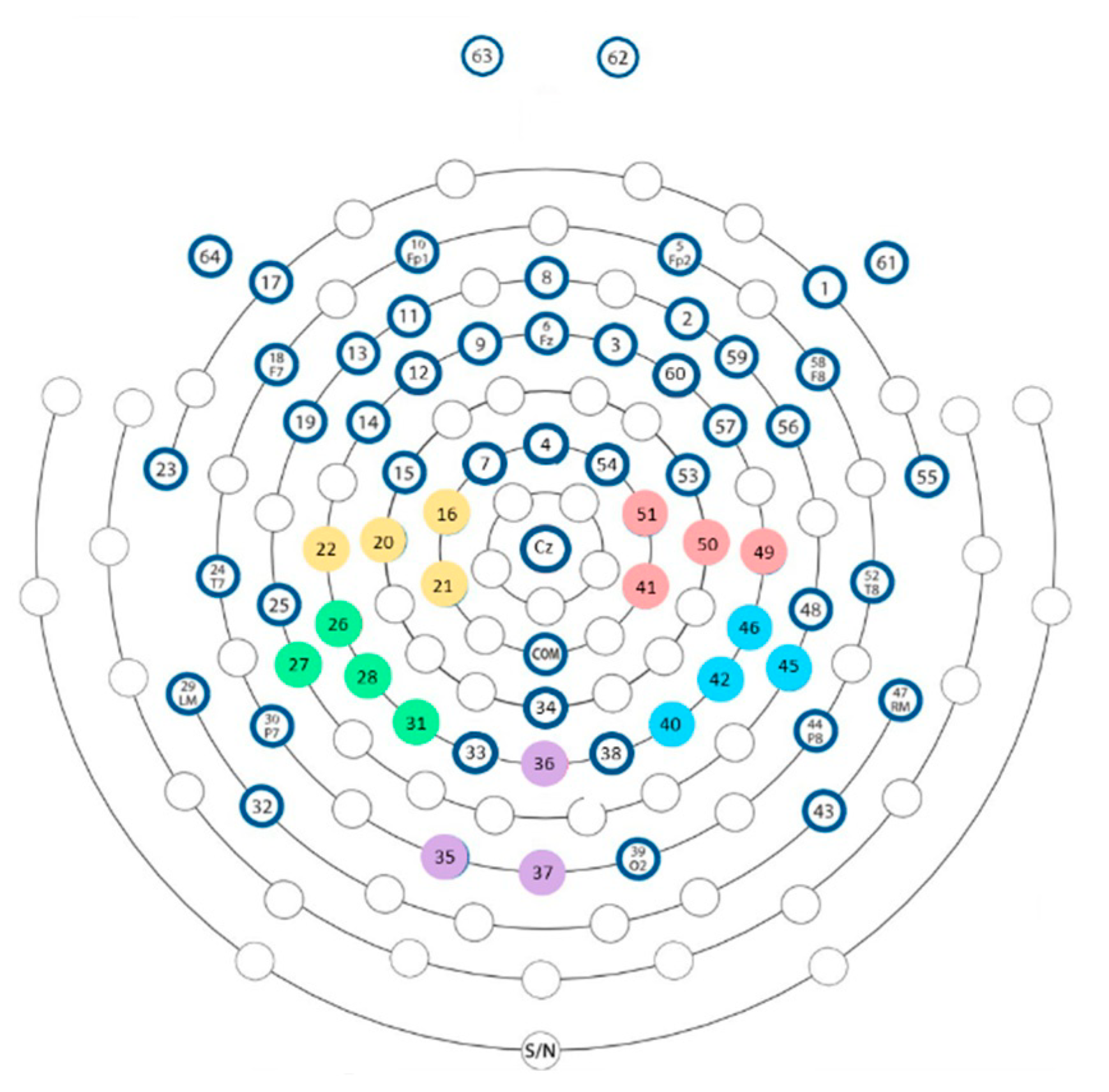
2.5.2. Electrocortical Activity Measures
2.6. Statistics
2.6.1. Clinical and Demographic Data
2.6.2. Behavioral Data
2.6.3. Electrocortical Activity Measures
3. Results
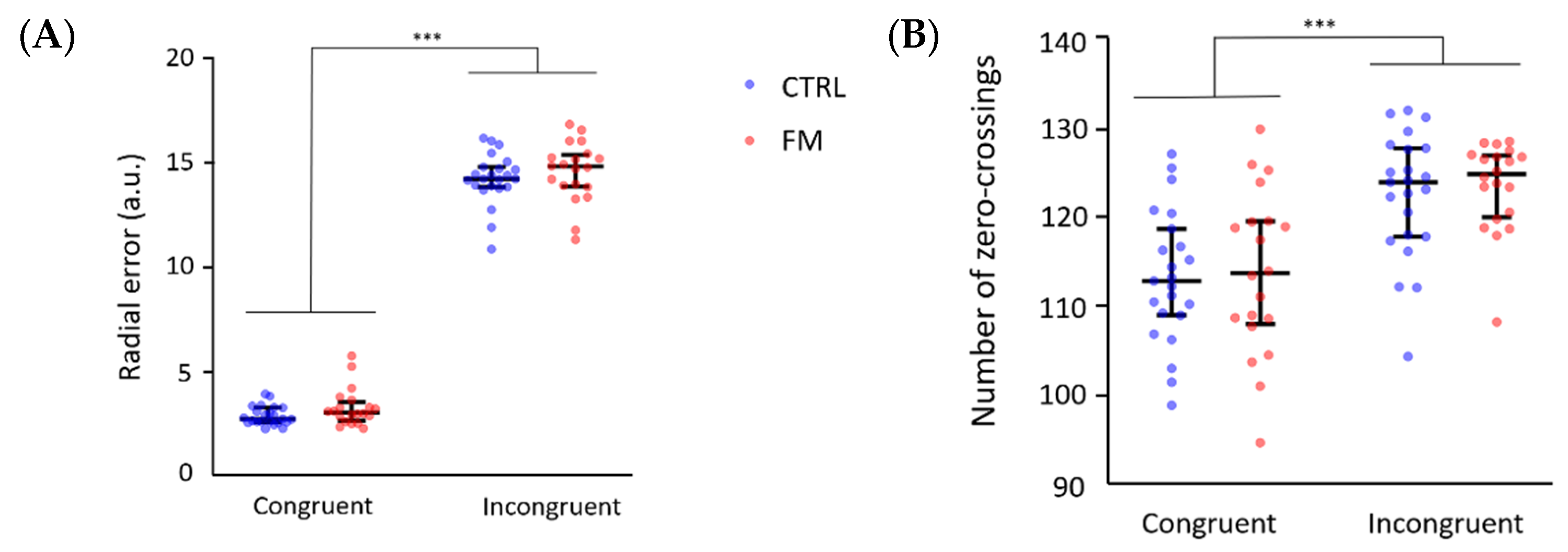
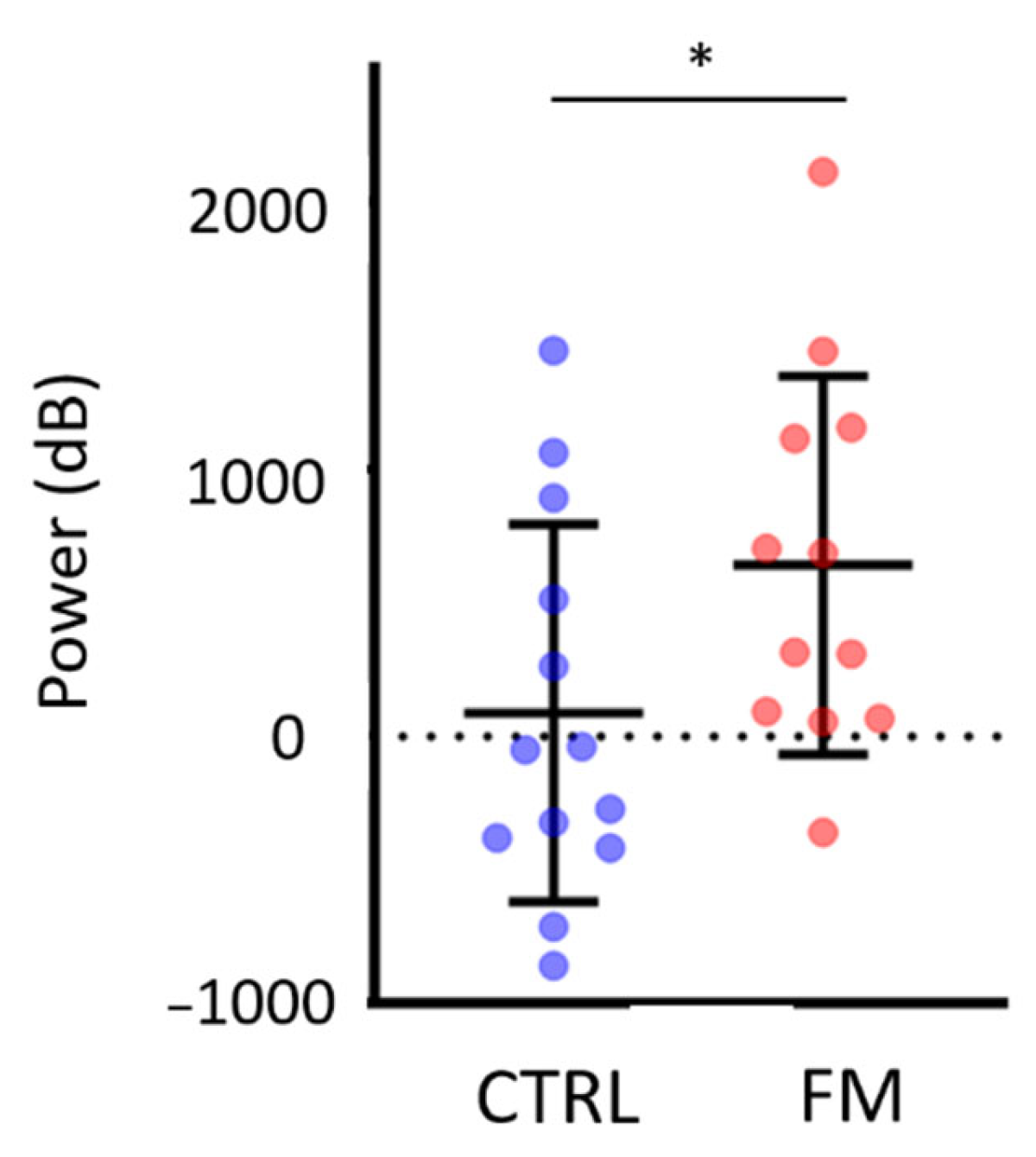
3.1. Behavioural Results
3.1.1. Results of the Complete Sample
3.1.2. Results of the Subsample of Participants for Which Electrocortical Activity Was Recorded
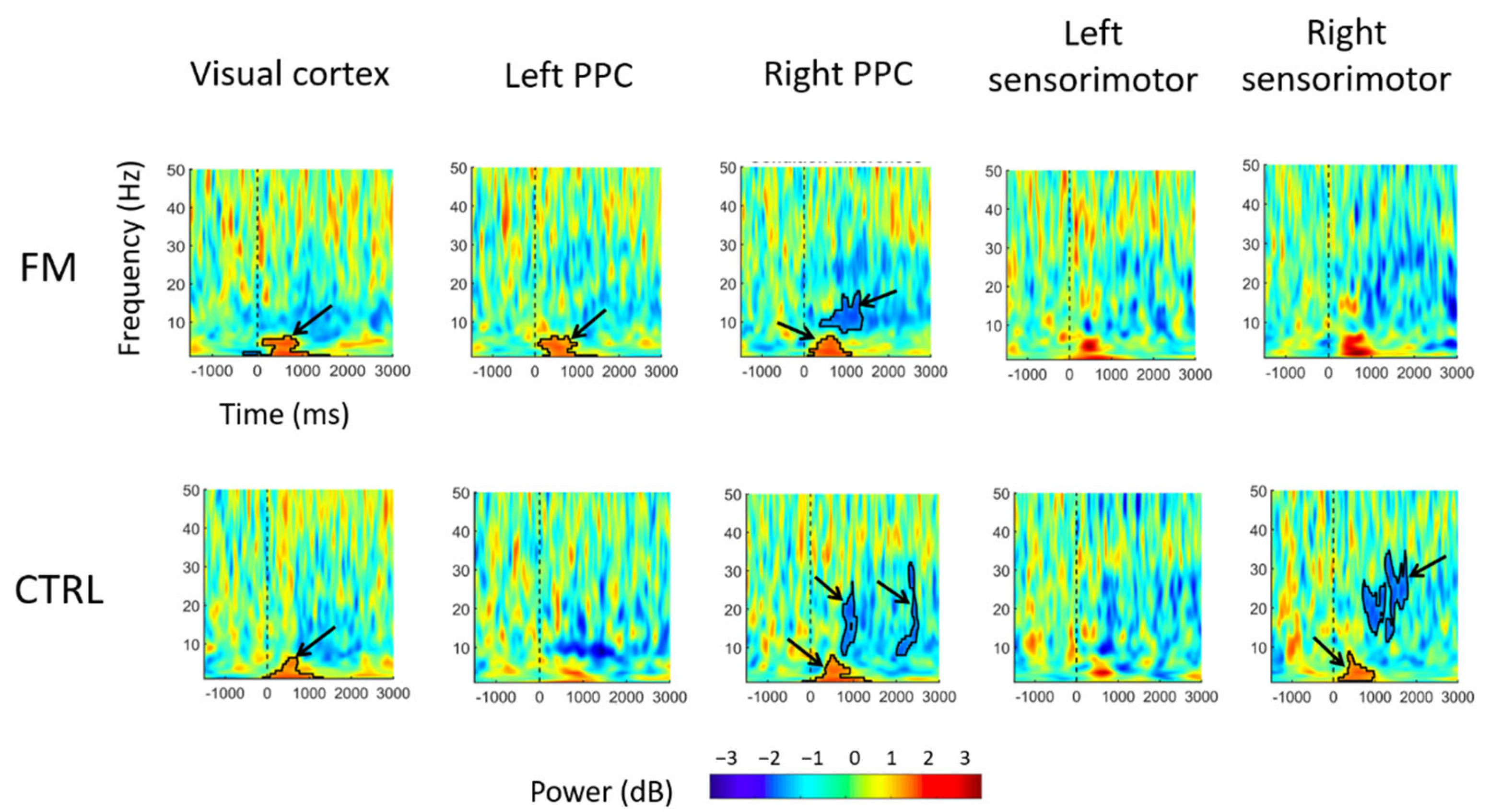
3.2. Electrocortical Activity Results
- Visual cortex: For both groups, a significant increase in theta (4–7 Hz) power in the Incongruent condition was observed within the first 1000 ms following sensorimotor conflict onset, compared with what was observed in the Congruent condition.
- PPC: In the left PPC, an increase in theta power was observed only for the FM group between 0 and 1000 ms after the onset of sensorimotor conflict in the Incongruent condition compared with the Congruent condition, whereas no significant difference was found for the Control group.In the right PPC, theta power increased between 0 and 1000 ms after conflict onset for both groups. A decrease in alpha power (8–12 Hz) was observed, from approximately 300 to 1200 ms after conflict onset for the FM group and from between 300 and 800 ms after conflict onset for the Control group. A decrease in beta power was also observed, with a cluster extending from approximately 2000 to 2500 ms after conflict onset in the Control group.
- Sensorimotor cortex: In the right sensorimotor cortex (ipsilateral to the tracing hand), there was a significant increase in theta power (a cluster extended from ~200 to ~1000 ms) after conflict onset, followed by a decrease in beta power (a cluster extended from ~500 to ~2000 ms), only for the Control group.
4. Discussion
4.1. Effect of the Sensorimotor Conflict on the Behavioral Measures
4.2. Effect of the Sensorimotor Conflict on the Electrocortical Activity
4.3. Clinical Implications
4.4. Limitations
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Crevecoeur, F.; Munoz, D.P.; Scott, S.H.; Crevecoeur, F.; Munoz, D.P.; Scott, S.H.; Crevecoeur, F.; Munoz, D.P.; Scott, S.H. Dynamic Multisensory Integration: Somatosensory Speed Trumps Visual Accuracy during Feedback Control. J. Neurosci. 2016, 36, 8598–8611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dieguez, S.; Lopez, C. The Bodily Self: Insights from Clinical and Experimental Research. Ann. Phys. Rehabil. Med. 2017, 60, 198–207. [Google Scholar] [CrossRef] [PubMed]
- Bouffard, J.; Gagné, M.; Mercier, C. Effect of Painful and Non-Painful Sensorimotor Manipulations on Subjective Body Midline. Front. Hum. Neurosci. 2013, 7, 77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gandevia, S.C.; Phegan, C.M.L. Perceptual Distortions of the Human Body Image Produced by Local Anaesthesia, Pain and Cutaneous Stimulation. J. Physiol. 1999, 514, 609–616. [Google Scholar] [CrossRef] [PubMed]
- Valenzuela-Moguillansky, C. Pain and Body Awareness: An Exploration of the Bodily Experience of Persons Suffering from Fibromyalgia. Constr. Found. 2013, 8, 339–350. [Google Scholar]
- Moseley, G.L. Distorted Body Image in Complex Regional Pain Syndrome. Neurology 2005, 65, 773. [Google Scholar] [CrossRef]
- Peltz, E.; Seifert, F.; Lanz, S.; Müller, R.; Maihöfner, C. Impaired Hand Size Estimation in CRPS. J. Pain 2011, 12, 1095–1101. [Google Scholar] [CrossRef]
- Moseley, G.L. I Can’t Find It! Distorted Body Image and Tactile Dysfunction in Patients with Chronic Back Pain. Pain 2008, 140, 239–243. [Google Scholar] [CrossRef]
- Frettlöh, J.; Hüppe, M.; Maier, C. Severity and Specificity of Neglect-like Symptoms in Patients with Complex Regional Pain Syndrome (CRPS) Compared to Chronic Limb Pain of Other Origins. Pain 2006, 124, 184–189. [Google Scholar] [CrossRef]
- Galer, B.S.; Jensen, M. Neglect-like Symptoms in Complex Regional Pain Syndrome: Results of a Self-Administered Survey. J. Pain Symptom Manag. 1999, 18, 213–217. [Google Scholar] [CrossRef]
- Heredia-Jimenez, J.; Orantes-Gonzalez, E.; Soto-Hermoso, V.M. Variability of Gait, Bilateral Coordination, and Asymmetry in Women with Fibromyalgia. Gait Posture 2016, 45, 41–44. [Google Scholar] [CrossRef] [PubMed]
- Pérez-De-Heredia-Torres, M.; Martínez-Piédrola, R.M.; Cigarán-Méndez, M.; Ortega-Santiago, R.; Fernández-De-Las-Peñas, C. Bilateral Deficits in Fine Motor Control Ability and Manual Dexterity in Women with Fibromyalgia Syndrome. Exp. Brain Res. 2013, 226, 137–143. [Google Scholar] [CrossRef] [PubMed]
- Eken, A.; Gökçay, D.; Yılmaz, C.; Baskak, B.; Baltacı, A.; Kara, M. Association of Fine Motor Loss and Allodynia in Fibromyalgia: An FNIRS Study. J. Mot. Behav. 2018, 50, 664–676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCabe, C.S.; Cohen, H.; Blake, D.R. Somaesthetic Disturbances in Fibromyalgia Are Exaggerated by Sensory—Motor Conflict: Implications for Chronicity of the Disease? Rheumatology 2007, 46, 1587–1592. [Google Scholar] [CrossRef] [Green Version]
- Lebar, N.; Danna, J.; Moré, S.; Mouchnino, L.; Blouin, J. On the Neural Basis of Sensory Weighting: Alpha, Beta and Gamma Modulations during Complex Movements. Neuroimage 2017, 150, 200–212. [Google Scholar] [CrossRef]
- Brun, C.; Gagné, M.; McCabe, C.S.; Mercier, C. Motor and Sensory Disturbances Induced by Sensorimotor Conflicts during Passive and Active Movements in Healthy Participants. PLoS ONE 2018, 13, e0203206. [Google Scholar] [CrossRef]
- Brun, C.; McCabe, C.S.; Mercier, C. The Contribution of Motor Commands to the Perturbations Induced by Sensorimotor Conflicts in Fibromyalgia. Neuroscience 2020, 434, 55–65. [Google Scholar] [CrossRef]
- Bultitude, J.H.; Petrini, K. Altered Visuomotor Integration in Complex Regional Pain Syndrome. Behav. Brain Res. 2021, 397, 112922. [Google Scholar] [CrossRef]
- Vittersø, A.D.; Buckingham, G.; Ten Brink, A.F.; Halicka, M.; Proulx, M.J.; Bultitude, J.H. Characterising sensorimotor adaptation in Complex Regional Pain Syndrome. Cortex 2021, 140, 157–178. [Google Scholar] [CrossRef]
- Brun, C.; Giorgi, N.; Pinard, A.M.; Gagné, M.; McCabe, C.S.; Mercier, C. Exploring the Relationships Between Altered Body Perception, Limb Position Sense, and Limb Movement Sense in Complex Regional Pain Syndrome. J. Pain 2019, 20, 17–27. [Google Scholar] [CrossRef] [Green Version]
- Dagenais, M.; Brun, C.; Ohayon, A.; Mercier, C. Virtual Reality in Fibromyalgia: Does Altering Visual Feedback Impact on Pain and Movement During Reaching? Front. Virtual Real. 2021, 2, 681034. [Google Scholar] [CrossRef]
- Brun, C.; Mercier, C.; Grieve, S.; Palmer, S.; Bailey, J.; McCabe, C.S. Sensory Disturbances Induced by Sensorimotor Conflicts Are Higher in Complex Regional Pain Syndrome and Fibromyalgia Compared to Arthritis and Healthy People, and Positively Relate to Pain Intensity. Eur. J. Pain 2019, 23, 483–494. [Google Scholar] [CrossRef] [PubMed]
- Bernier, P.M.; Burle, B.; Vidal, F.; Hasbroucq, T.; Blouin, J. Direct Evidence for Cortical Suppression of Somatosensory Afferents during Visuomotor Adaptation. Cereb. Cortex 2009, 19, 2106–2113. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.-J. Neurophysiological and Computational Principles of Cortical Rhythms in Cognition. Physiol. Rev. 2010, 90, 1195–1268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krebber, M.; Harwood, J.; Spitzer, B.; Keil, J.; Senkowski, D. Visuotactile Motion Congruence Enhances Gamma-Band Activity in Visual and Somatosensory Cortices. Neuroimage 2015, 117, 160–169. [Google Scholar] [CrossRef] [PubMed]
- Engel, A.K.; Senkowski, D.; Schneider, T.R. Multisensory Integration through Neural Coherence; CRC Press/Taylor & Francis: Boca Raton, FL, USA, 2012; ISBN 9781439812174. [Google Scholar]
- Balslev, D.; Christensen, L.O.D.; Lee, J.H.; Law, I.; Paulson, O.B.; Miall, R.C. Enhanced Accuracy in Novel Mirror Drawing after Repetitive Transcranial Magnetic Stimulation-Induced Proprioceptive Deafferentation. J. Neurosci. 2004, 24, 9698–9702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goldenkoff, E.R.; McGregor, H.R.; Mergos, J.; Gholizadeh, P.; Bridenstine, J.; Brown, M.J.N.; Vesia, M. Reversal of Visual Feedback Modulates Somatosensory Plasticity. Neuroscience 2021, 452, 335–344. [Google Scholar] [CrossRef]
- Yoon, H.C.; Lee, K.H.; Huh, D.C.; Lee, J.H.; Lee, D.H. Effects of Repetitive Transcranial Magnetic Stimulation on the Somatosensory Cortex during Prism Adaptation. Percept. Mot. Ski. 2014, 118, 491–506. [Google Scholar] [CrossRef]
- Perfetti, B.; Moisello, C.; Landsness, E.C.; Kvint, S.; Lanzafame, S.; Onofrj, M.; di Rocco, A.; Tononi, G.; Felice Ghilardi, M. Modulation of Gamma and Theta Spectral Amplitude and Phase Synchronization Is Associated with the Development of Visuo-Motor Learning. J. Neurosci. 2011, 31, 14810–14819. [Google Scholar] [CrossRef] [Green Version]
- Zama, T.; Takahashi, Y.; Shimada, S. Simultaneous EEG-NIRS Measurement of the Inferior Parietal Lobule During a Reaching Task with Delayed Visual Feedback. Front. Hum. Neurosci. 2019, 13, 301. [Google Scholar] [CrossRef]
- Wolfe, F.; Clauw, D.J.; Fitzcharles, M.A.; Goldenberg, D.L.; Häuser, W.; Katz, R.L.; Mease, P.J.; Russell, A.S.; Russell, I.J.; Walitt, B. 2016 Revisions to the 2010/2011 Fibromyalgia Diagnostic Criteria. Semin. Arthritis Rheum. 2016, 46, 319–329. [Google Scholar] [CrossRef] [PubMed]
- Rehm, S.E.; Koroschetz, J.; Gockel, U.; Brosz, M.; Freynhagen, R.; Tölle, T.R.; Baron, R. A Cross-Sectional Survey of 3035 Patients with Fibromyalgia: Subgroups of Patients with Typical Comorbidities and Sensory Symptom Profiles. Rheumatology 2010, 49, 1146–1152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kleykamp, B.A.; Ferguson, M.C.; McNicol, E.; Bixho, I.; Arnold, L.M.; Edwards, R.R.; Fillingim, R.; Grol-Prokopczyk, H.; Turk, D.C.; Dworkin, R.H. The Prevalence of Psychiatric and Chronic Pain Comorbidities in Fibromyalgia: An ACTTION Systematic Review. Semin. Arthritis Rheum. 2021, 51, 166–174. [Google Scholar] [CrossRef] [PubMed]
- Weir, P.T.; Harlan, G.A.; Nkoy, F.L.; Jones, S.S.; Hegmann, K.T.; Gren, L.H.; Lyon, J.L. The Incidence of Fibromyalgia and Its Associated Comorbidities: A Population-Based Retrospective Cohort Study Based on International Classification of Diseases, 9th Revision Codes. J. Clin. Rheumatol. 2006, 12, 124–128. [Google Scholar] [CrossRef] [PubMed]
- Lebar, N.; Bernier, P.M.; Guillaume, A.; Mouchnino, L.; Blouin, J. Neural Correlates for Task-Relevant Facilitation of Visual Inputs during Visually-Guided Hand Movements. Neuroimage 2015, 121, 39–50. [Google Scholar] [CrossRef]
- Oldfield, R.C. The Assessment and Analysis of Handedness: The Edinburgh Inventory. Neuropsychologia 1971, 9, 97–113. [Google Scholar] [CrossRef]
- Ransil, B.J.; Schachter, S.C. Test-Retest Reliability of the Edinburgh Handedness Inventory and Global Handedness Preference Measurements, and Their Correlation. Percept. Mot. Ski. 1994, 79, 1355–1372. [Google Scholar] [CrossRef]
- Veale, J.F. Edinburgh Handedness Inventory—Short Form: A Revised Version Based on Confirmatory Factor Analysis. Laterality 2014, 19, 164–177. [Google Scholar] [CrossRef]
- Krakauer, J.W.; Ghilardi, M.F.; Mentis, M.; Barnes, A.; Veytsman, M.; Eidelberg, D.; Ghez, C. Differential Cortical and Subcortical Activations in Learning Rotations and Gains for Reaching: A PET Study. J. Neurophysiol. 2004, 91, 924–933. [Google Scholar] [CrossRef]
- Balslev, D.; Nielsen, F.Å.; Paulson, O.B.; Law, I. Right Temporoparietal Cortex Activation during Visuo-Proprioceptive Conflict. Cerebral Cortex 2005, 15, 166–169. [Google Scholar] [CrossRef]
- Wolfe, F.; Smythe, H.A.; Yunus, M.B.; Bennett, R.M.; Bombardier, C.; Goldenberg, D.L.; Tugwell, P.; Campbell, S.M.; Abeles, M.; Clark, P. The American College of Rheumatology 1990 Criteria for the Classification of Fibromyalgia. Report of the Multicenter Criteria Committee. Arthritis Rheum. 1990, 33, 160–172. [Google Scholar] [CrossRef] [PubMed]
- Wolfe, F.; Clauw, D.J.; Fitzcharles, M.A.; Goldenberg, D.L.; Katz, R.S.; Mease, P.; Russell, A.S.; Russell, I.J.; Winfield, J.B.; Yunus, M.B. The American College of Rheumatology Preliminary Diagnostic Criteria for Fibromyalgia and Measurement of Symptom Severity. Arthritis Care Res. 2010, 62, 600–610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cleeland, C.S.; Ryan, K.M. Pain Assessment: Global Use of the Brief Pain Inventory. Ann. Acad. Med. Singap. 1994, 23, 129–138. [Google Scholar] [PubMed]
- Poundja, J.; Fikretoglu, D.; Guay, S.; Brunet, A. Validation of the French Version of the Brief Pain Inventory in Canadian Veterans Suffering from Traumatic Stress. J. Pain Symptom Manag. 2007, 33, 720–726. [Google Scholar] [CrossRef] [PubMed]
- Moberg, E. Two-Point Discrimination Test. Scand. J. Rehabil. Med. 1990, 22, 127–134. [Google Scholar] [CrossRef]
- Martínez, E.; Guillen, V.; Buesa, I.; Azkue, J.J. A Distorted Body Schema and Susceptibility to Experiencing Anomalous Somatosensory Sensations in Fibromyalgia Syndrome. Clin. J. Pain 2019, 35, 887–893. [Google Scholar] [CrossRef]
- Lajoie, Y.; Paillard, J.; Teasdale, N.; Bard, C.; Fleury, M.; Forget, R.; Lamarre, Y. Mirror Drawing in a Deafferented Patient and Normal Subjects Visuoproprioceptive Conflict. Neurology 1992, 42, 1104–1106. [Google Scholar] [CrossRef]
- Brainard, D.H. The Psychophysics Toolbox. Spat. Vis. 1997, 10, 433–436. [Google Scholar] [CrossRef] [Green Version]
- Ofori, E.; Coombes, S.A.; Vaillancourt, D.E. 3D Cortical Electrophysiology of Ballistic Upper Limb Movement in Humans. Neuroimage 2015, 115, 30–41. [Google Scholar] [CrossRef] [Green Version]
- Flash, T.; Hogans, N. The Coordination of Arm Movements: An Experimentally Confirmed Mathematical Model. J. Neurosci. 1985, 5, 1688–1703. [Google Scholar] [CrossRef] [Green Version]
- Delorme, A.; Makeig, S. EEGLAB: An Open Source Toolbox for Analysis of Single-Trial EEG Dynamics Including Independent Component Analysis. J. Neurosci. Methods 2004, 134, 9–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyakoshi, M.; Schmitt, L.M.; Erickson, C.A.; Sweeney, J.A.; Pedapati, E.V. Can We Push the “Quasi-Perfect Artifact Rejection” Even Closer to Perfection? Front. Neuroinform. 2021, 14, 597079. [Google Scholar] [CrossRef] [PubMed]
- Cohen, M.X. Analyzing Neural Time Series Data: Theory and Practice; MIT Press: Cambridge, MA, USA, 2014. [Google Scholar]
- Jensen, O.; Spaak, E.; Zumer, J.M. Human Brain Oscillations: From Physiological Mechanisms to Analysis and Cognition. In Magnetoencephalography; Springer: Berlin/Heidelberg, Germany, 2014; Volume 9783642330, ISBN 9783642330452. [Google Scholar]
- Watanabe, T.; Mima, T.; Shibata, S.; Kirimoto, H. Midfrontal Theta as Moderator between Beta Oscillations and Precision Control. Neuroimage 2021, 235, 118022. [Google Scholar] [CrossRef] [PubMed]
- Struber, L.; Baumont, M.; Barraud, P.A.; Nougier, V.; Cignetti, F. Brain Oscillatory Correlates of Visuomotor Adaptive Learning. Neuroimage 2021, 245, 118645. [Google Scholar] [CrossRef]
- Pfurtscheller, G.; Lopes Da Silva, F.H. Event-Related EEG/MEG Synchronization and Desynchronization: Basic Principles. Clin. Neurophysiol. 1999, 110, 1842–1857. [Google Scholar] [CrossRef]
- Stančák, A., Jr. The Electroencephalographic Beta Synchronization Following Extension and Flexion Finger Movements in Humans. Neurosci. Lett. 2000, 284, 41–44. [Google Scholar] [CrossRef]
- Noguchi, K.; Gel, Y.R.; Brunner, E.; Konietschke, F. NparLD: An R Software Package for the Nonparametric Analysis of Longitudinal Data in Factorial Experiments. J. Stat. Softw. 2012, 50, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Vargha, A.; Delaney, H.D. A Critique and Improvement of the “CL” Common Language Effect Size Statistics of McGraw and Wong. J. Educ. Behav. Stat. 2000, 25, 101–132. [Google Scholar]
- Maris, E.; Oostenveld, R. Nonparametric Statistical Testing of EEG- and MEG-Data. J. Neurosci. Methods 2007, 164, 177–190. [Google Scholar] [CrossRef]
- Nichols, T.E.; Holmes, A.P. Nonparametric Permutation Tests for Functional Neuroimaging: A Primer with Examples. Hum. Brain Mapp. 2001, 15, 1–25. [Google Scholar] [CrossRef] [Green Version]
- Clower, D.M.; Boussaoud, D. Selective Use of Perceptual Recalibration versus Visuomotor Skill Acquisition. J. Neurophysiol. 2000, 84, 2703–2708. [Google Scholar] [CrossRef] [PubMed]
- Van Beers, R.J.; Wolpert, D.M.; Haggard, P. When Feeling Is More Important Than Seeing in Sensorimotor Adaptation. Curr. Biol. 2002, 12, 834–837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lemieux, L.G.; Simoneau, M.; Tessier, J.F.; Billot, M.; Blouin, J.; Teasdale, N. Balance Control Interferes with the Tracing Performance of a Pattern with Mirror-Reversed Vision in Older Persons. Age 2014, 36, 823–837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, K.; Körding, K. Uncertainty of Feedback and State Estimation Determines the Speed of Motor Adaptation. Front. Comput. Neurosci. 2010, 4, 11. [Google Scholar] [CrossRef] [Green Version]
- Bultitude, J.H.; Juravle, G.; Spence, C. Tactile Gap Detection Deteriorates during Bimanual Symmetrical Movements under Mirror Visual Feedback. PLoS ONE 2016, 11, e0146077. [Google Scholar] [CrossRef] [Green Version]
- Cavanagh, J.F.; Frank, M.J. Frontal Theta as a Mechanism for Cognitive Control. Trends Cogn. Sci. 2014, 18, 414–421. [Google Scholar] [CrossRef] [Green Version]
- Beldzik, E.; Ullsperger, M.; Domagalik, A.; Marek, T. Conflict- and Error-Related Theta Activities Are Coupled to BOLD Signals in Different Brain Regions. Neuroimage 2022, 256, 119264. [Google Scholar] [CrossRef]
- Nigbur, R.; Ivanova, G.; Stürmer, B. Theta Power as a Marker for Cognitive Interference. Clin. Neurophysiol. 2011, 122, 2185–2194. [Google Scholar] [CrossRef]
- Alsuradi, H.; Park, W.; Eid, M. Midfrontal Theta Oscillation Encodes Haptic Delay. Sci. Rep. 2021, 11, 17074. [Google Scholar] [CrossRef]
- Roa Romero, Y.; Keil, J.; Balz, J.; Gallinat, J.; Senkowski, D.; Romero, R.Y.; Reduced, S.D. Reduced Frontal Theta Oscillations Indicate Altered Crossmodal Prediction Error Processing in Schizophrenia. J. Neurophysiol. 2016, 116, 1396–1407. [Google Scholar] [CrossRef] [Green Version]
- Rangel-Pacheco, A.; Lew, B.J.; Schantell, M.D.; Frenzel, M.R.; Eastman, J.A.; Wiesman, A.I.; Wilson, T.W. Altered Fronto-Occipital Connectivity during Visual Selective Attention in Regular Cannabis Users. Psychopharmacology 2021, 238, 1351–1361. [Google Scholar] [CrossRef] [PubMed]
- Arrighi, P.; Bonfiglio, L.; Minichilli, F.; Cantore, N.; Carboncini, M.C.; Piccotti, E.; Rossi, B.; Andre, P. EEG Theta Dynamics within Frontal and Parietal Cortices for Error Processing during Reaching Movements in a Prism Adaptation Study Altering Visuo-Motor Predictive Planning. PLoS ONE 2016, 11, e0150265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pastötter, B.; Dreisbach, G.; Bäuml, K.H.T. Dynamic Adjustments of Cognitive Control: Oscillatory Correlates of the Conflict Adaptation Effect. J. Cogn. Neurosci. 2013, 25, 2167–2178. [Google Scholar] [CrossRef] [PubMed]
- Kieffaber, P.D.; Hershaw, J.; Sredl, J.; West, R. Electrophysiological Correlates of Error Initiation and Response Correction. Neuroimage 2016, 128, 158–166. [Google Scholar] [CrossRef] [PubMed]
- Sipp, A.R.; Gwin, J.T.; Makeig, S.; Ferris, D.P. Loss of Balance during Balance Beam Walking Elicits a Multifocal Theta Band Electrocortical Response. J. Neurophysiol. 2013, 110, 2050–2060. [Google Scholar] [CrossRef] [Green Version]
- Contreras-Vidal, J.L.; Kerick, S.E. Independent Component Analysis of Dynamic Brain Responses during Visuomotor Adaptation. Neuroimage 2004, 21, 936–945. [Google Scholar] [CrossRef]
- Morís Fernández, L.; Torralba, M.; Soto-Faraco, S. Theta Oscillations Reflect Conflict Processing in the Perception of the McGurk Illusion. Eur. J. Neurosci. 2018, 48, 2630–2641. [Google Scholar] [CrossRef]
- Frith, C.D.; Blakemore, S.J.; Wolpert, D.M. Abnormalities in the Awareness and Control of Action. Philos. Trans. R. Soc. B Biol. Sci. 2000, 355, 1771–1788. [Google Scholar] [CrossRef] [Green Version]
- Scott, S.H. Optimal Feedback Control and the Neural Basis of Volitional Motor Control. Nat. Rev. Neurosci. 2004, 5, 532–544. [Google Scholar] [CrossRef]
- Shadmehr, R.; Smith, M.A.; Krakauer, J.W. Error Correction, Sensory Prediction, and Adaptation in Motor Control. Annu. Rev. Neurosci. 2010, 33, 89–108. [Google Scholar] [CrossRef] [Green Version]
- Kim, O.A.; Forrence, A.D.; McDougle, S.D. Motor Learning without Movement. Proc. Natl. Acad. Sci. USA 2022, 119, e2204379119. [Google Scholar] [CrossRef] [PubMed]
- Benyamini, M.; Demchenko, I.; Zacksenhouse, M. Error Related EEG Potentials Evoked by Visuo-Motor Rotations. Brain Res. 2021, 1769, 147606. [Google Scholar] [CrossRef] [PubMed]
- Savoie, F.A.; Thénault, F.; Whittingstall, K.; Bernier, P.M. Visuomotor Prediction Errors Modulate EEG Activity Over Parietal Cortex. Sci. Rep. 2018, 8, 12513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mazzoni, P.; Krakauer, J.W. An Implicit Plan Overrides an Explicit Strategy during Visuomotor Adaptation. J. Neurosci. 2006, 26, 3642–3645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diedrichsen, J.; Hashambhoy, Y.; Rane, T.; Shadmehr, R. Neural Correlates of Reach Errors. J. Neurosci. 2005, 25, 9919–9931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mutha, P.K.; Sainburg, R.L.; Haaland, K.Y. Critical Neural Substrates for Correcting Unexpected Trajectory Errors and Learning from Them. Brain 2011, 134, 3644–3658. [Google Scholar] [CrossRef] [Green Version]
- Newport, R.; Brown, L.; Husain, M.; Mort, D.; Jackson, S.R. The Role of the Posterior Parietal Lobe in Prism Adaptation: Failure to Adapt to Optical Prisms in a Patient with Bilateral Damage to Posterior Parietal Cortex. Cortex 2006, 42, 720–729. [Google Scholar] [CrossRef]
- Amassian, V.E.; Cracco, R.Q.; Maccabee, P.J.; Cracco, J.B.; Rudell, A.; Eberle, L. Suppression of Visual Perception by Magnetic Coil Stimulation of Human Occipital Cortex. Electroencephalogr. Clin. Neurophysiol./Evoked Potentials Sect. 1989, 74, 459–462. [Google Scholar] [CrossRef]
- Ruben, J.; Schwiemann, J.; Deuchert, M.; Meyer, R.; Krause, T.; Curio, G.; Villringer, K.; Kurth, R.; Villringer, A. Somatotopic Organization of Human Secondary Somatosensory Cortex. Cereb. Cortex 2001, 11, 463–473. [Google Scholar] [CrossRef]
- Pasalar, S.; Ro, T.; Beauchamp, M.S. TMS of Posterior Parietal Cortex Disrupts Visual Tactile Multisensory Integration. Eur. J. Neurosci. 2010, 31, 1783–1790. [Google Scholar] [CrossRef] [Green Version]
- Augière, T.; Desjardins, A.; Paquette Raynard, E.; Brun, C.; Pinard, A.M.; Simoneau, M.; Mercier, C. Tactile Detection in Fibromyalgia: A Systematic Review and a Meta-Analysis. Front. Pain Res. 2021, 2, 116. [Google Scholar] [CrossRef] [PubMed]
- Del Percio, C.; Babiloni, C.; Bertollo, M.; Marzano, N.; Iacoboni, M.; Infarinato, F.; Lizio, R.; Stocchi, M.; Robazza, C.; Cibelli, G.; et al. Visuo-Attentional and Sensorimotor Alpha Rhythms Are Related to Visuo-Motor Performance in Athletes. Hum. Brain Mapp. 2009, 30, 3527–3540. [Google Scholar] [CrossRef] [PubMed]
- Jensen, O.; Mazaheri, A. Shaping Functional Architecture by Oscillatory Alpha Activity: Gating by Inhibition. Front. Hum. Neurosci. 2010, 4, 186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfurtscheller, G. Central Beta Rhythm during Sensorimotor Activities in Man. Electroencephalogr. Clin. Neurophysiol. 1981, 51, 253–264. [Google Scholar] [CrossRef]
- Kilavik, B.E.; Zaepffel, M.; Brovelli, A.; MacKay, W.A.; Riehle, A. The Ups and Downs of Beta Oscillations in Sensorimotor Cortex. Exp. Neurol. 2013, 245, 15–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goodale, M.A.; Milner, A.D. Separate Visual Pathways for Perception and Action. Trends Neurosci. 1992, 15, 20–25. [Google Scholar] [CrossRef] [PubMed]
- Cañal-Bruland, R.; Voorwald, F.; Wielaard, K.; van der Kamp, J. Dissociations between Vision for Perception and Vision for Action Depend on the Relative Availability of Egocentric and Allocentric Information. Atten. Percept. Psychophys. 2013, 75, 1206–1214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rossetti, Y.; Pisella, L.; McIntosh, R.D. Rise and Fall of the Two Visual Systems Theory. Ann. Phys. Rehabil. Med. 2017, 60, 130–140. [Google Scholar] [CrossRef]
- Wolfe, F.; Walitt, B.; Perrot, S.; Rasker, J.J.; Häuser, W. Fibromyalgia Diagnosis and Biased Assessment: Sex, Prevalence and Bias. PLoS ONE 2018, 13, e0203755. [Google Scholar] [CrossRef] [Green Version]
- Limanowski, J. Precision Control for a Flexible Body Representation. Neurosci. Biobehav. Rev. 2021, 34, 104401. [Google Scholar] [CrossRef]
- Ulus, Y.; Akyol, Y.; Tander, B.; Bilgici, A.; Kuru, Ö. Knee Proprioception and Balance in Turkish Women and without Fibromyalgia Syndrome. Turk. J. Phys. Med. Rehabil./Turk. Fiz. Tip Ve Rehabil. Derg. 2013, 59, 128–132. [Google Scholar] [CrossRef]
- Bardal, E.M.; Roeleveld, K.; Johansen, T.O.; Mork, P.J. Upper Limb Position Control in Fibromyalgia. BMC Musculoskelet. Disord. 2012, 13, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toprak Celenay, S.; Mete, O.; Coban, O.; Oskay, D.; Erten, S. Trunk Position Sense, Postural Stability, and Spine Posture in Fibromyalgia. Rheumatol. Int. 2019, 39, 2087–2094. [Google Scholar] [CrossRef] [PubMed]
- Bardal, E.M.; Roeleveld, K.; Ihlen, E.; Mork, P.J. Micro Movements of the Upper Limb in Fibromyalgia: The Relation to Proprioceptive Accuracy and Visual Feedback. J. Electromyogr. Kinesiol. 2016, 26, 1–7. [Google Scholar] [CrossRef]
- Gracely, R.H.; Petzke, F.; Wolf, J.M.; Clauw, D.J. Functional Magnetic Resonance Imaging Evidence of Augmented Pain Processing in Fibromyalgia. Arthritis Rheum. 2002, 46, 1333–1343. [Google Scholar] [CrossRef]
- Fagerlund, A.J.; Iversen, M.; Ekeland, A.; Moen, C.M.; Aslaksen, P.M. Blame It on the Weather? The Association between Pain in Fibromyalgia, Relative Humidity, Temperature and Barometric Pressure. PLoS ONE 2019, 14, e0216902. [Google Scholar] [CrossRef] [Green Version]
- Rau, C.; Plewnia, C.; Hummel, F.; Gerloff, C. Event-Related Desynchronization and Excitability of the Ipsilateral Motor Cortex during Simple Self-Paced Finger Movements. Clin. Neurophysiol. 2003, 114, 1819–1826. [Google Scholar] [CrossRef]
- Schnitzler, A.; Salmelin, R.; Salenius, S.; Jousmäki, V.; Hari, R. Tactile Information from the Human Hand Reaches the Ipsilateral Primary Somatosensory Cortex. Neurosci. Lett. 1995, 200, 25–28. [Google Scholar] [CrossRef]
- Cheyne, D.; Gaetz, W.; Garnero, L.; Lachaux, J.P.; Ducorps, A.; Schwartz, D.; Varela, F.J. Neuromagnetic Imaging of Cortical Oscillations Accompanying Tactile Stimulation. Cogn. Brain Res. 2003, 17, 599–611. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Participant | Sex | Age (years) | Currently Working? | Pain Duration (years) | BPI: Pain Severity | BPI: Pain Interference | Pharmacological Treatments | Non-Pharmacological Treatments | Current Comorbidities |
|---|---|---|---|---|---|---|---|---|---|
| S02 | F | 66 | no | 32 | 4.8 ± 0.8 | 6.4 ± 1.2 | Acetaminophen, tridural, tramacet, amitriptyline, gabapentine | Migraines | |
| S04 | F | 56 | no | 11 | 6.5 ± 1.7 | 3.4 ± 3 | Naproxen | Physiotherapy, osteopathy | Hypothyroidism, coeliac disease |
| S06 | F | 32 | yes | 7 | 3.5 ± 1.7 | 1.6 ± 1.2 | Amitriptyline, duloxetine, bisoprolol | Psychotherapy, massage, acupuncture, TENS | Tachycardia, chronic fatigue syndrome |
| S14 | F | 59 | no | 3 | 7 ± 0.7 | 5.6 ± 2.8 | Venlafaxine | Meditation | Epstein-Barr virus, irritable bowel syndrome |
| S15 | F | 20 | yes | 3 | 4.8 ± 1.1 | 5 ± 1.7 | Diclofenac, duloxetine | Psychotherapy, acupuncture, massage, nutritionist | Restless leg syndrome, arthritis, irritable bowel syndrome, migraines, generalized anxiety disorder, borderline personality disorder, eating disorder, attention and hyperactivity disorder, triple X syndrome |
| S16 | F | 34 | yes | 21 | 4 ± 1.2 | 2.1 ± 2.6 | Cyclobenzaprine | Psychotherapy | Arthritis, borderline personality disorder, migraines, endometriosis, post-traumatic stress disorder, hyperlaxity |
| S21 | F | 64 | no | 12 | 4.8 ± 0.8 | 4.6 ± 1.8 | Acetaminophen, codeine | Physiotherapy | Eczema, asthma, irritable bowel syndrome, arthritis, Raynaud’s disease |
| S39 | F | 52 | no | 10 | 3 ± 1.2 | 0.6 ± 0.9 | Pregabalin, acetaminophen | Massage, chiropractic | Aerophagia |
| S01 | F | 45 | yes | 31 | 5.5 ± 1.5 | 5.4 ± 1.8 | Pregabalin, naproxen, amitriptyline, escitaloprame | Physiotherapy, meditation | Slipped disc, hypothyroidism, chronic fatigue syndrome, kinesiophobia |
| S03 | F | 51 | yes | 46 | 4.5 ± 0.9 | 5 ± 1.9 | None | Physiotherapy, psychotherapy | Biliary cirrhosis, hypothyroidism, generalized anxiety disorder, post-traumatic stress disorder, type 2 diabetes, obesity, sleep apnea, asthma, migraines, chronic fatigue syndrome, depression |
| S05 | F | 21 | yes | 9 | 4 ± 0.7 | 0.9 ± 0.8 | Aventyl | Physiotherapy, massage | Irritable bowel syndrome, migraines |
| S08 | F | 23 | yes | 5 | 5.3 ± 2.3 | 3.6 ± 2.8 | Acetaminophen, cyclobenzaprine | Massage | Attention disorder, post-traumatic stress disorder |
| S11 | F | 39 | no | 39 | 1.5 ± 1.1 | 1.6 ± 1.2 | None | Chiropractic, osteopathy, massage | |
| S12 | F | 51 | no | 6 | 5.8 ± 1.8 | 7 ± 1.2 | Flexeril | Massage | Depression |
| S13 | F | 41 | yes | 41 | 7 ± 0.7 | 5.1 ± 3.1 | Pregabalin, celebrex, flexeril, cannabis | Psychotherapy, physiotherapy, osteopathy | Depression, rhumatoid arthritis, irritable bowel syndrome, migraines |
| S17 | F | 48 | no | 10 | 5 ± 2 | 5.3 ± 1.7 | Restoril | Psychotherapy, osteopathy, physiotherapy, acupuncture | Sclero-atrophic lichen |
| S19 | F | 48 | no | 9 | 4.3 ± 1.5 | 5.4 ± 2.6 | None | Osteopathy | |
| S22 | F | 24 | yes | 10 | 4.8 ± 1.1 | 4 ± 2.4 | Pregabalin | Osteopathy | Hyperactivity disorder, hypothyroidism, irritable bowel syndrome, asthma, migraines |
| S23 | F | 37 | yes | 12 | 6 ± 1 | 4.9 ± 1.4 | Ibuprofen, acetaminophen, decontractyl | Massage, chiropractic, osteopathy | Type I diabetes |
| S26 | F | 66 | no | 21 | 4 ± 1.6 | 4.9 ± 2.4 | None | Psychotherapy, physiotherapy, meditation | Irritable bowel syndrome, osteoporosis, chronic rhinitis |
| Median ± IQR | 10.5 ± 15 | 4.8 ± 1.6 | 4.9 ± 2.3 | ||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Augière, T.; Simoneau, M.; Brun, C.; Pinard, A.M.; Blouin, J.; Mouchnino, L.; Mercier, C. Behavioral and Electrocortical Response to a Sensorimotor Conflict in Individuals with Fibromyalgia. Brain Sci. 2023, 13, 931. https://doi.org/10.3390/brainsci13060931
Augière T, Simoneau M, Brun C, Pinard AM, Blouin J, Mouchnino L, Mercier C. Behavioral and Electrocortical Response to a Sensorimotor Conflict in Individuals with Fibromyalgia. Brain Sciences. 2023; 13(6):931. https://doi.org/10.3390/brainsci13060931
Chicago/Turabian StyleAugière, Tania, Martin Simoneau, Clémentine Brun, Anne Marie Pinard, Jean Blouin, Laurence Mouchnino, and Catherine Mercier. 2023. "Behavioral and Electrocortical Response to a Sensorimotor Conflict in Individuals with Fibromyalgia" Brain Sciences 13, no. 6: 931. https://doi.org/10.3390/brainsci13060931