


High-Definition Transcranial Direct Current Stimulation Improves Decision-Making Ability: A Study Based on EEG

Abstract
:1. Introduction
2. Materials and Methods
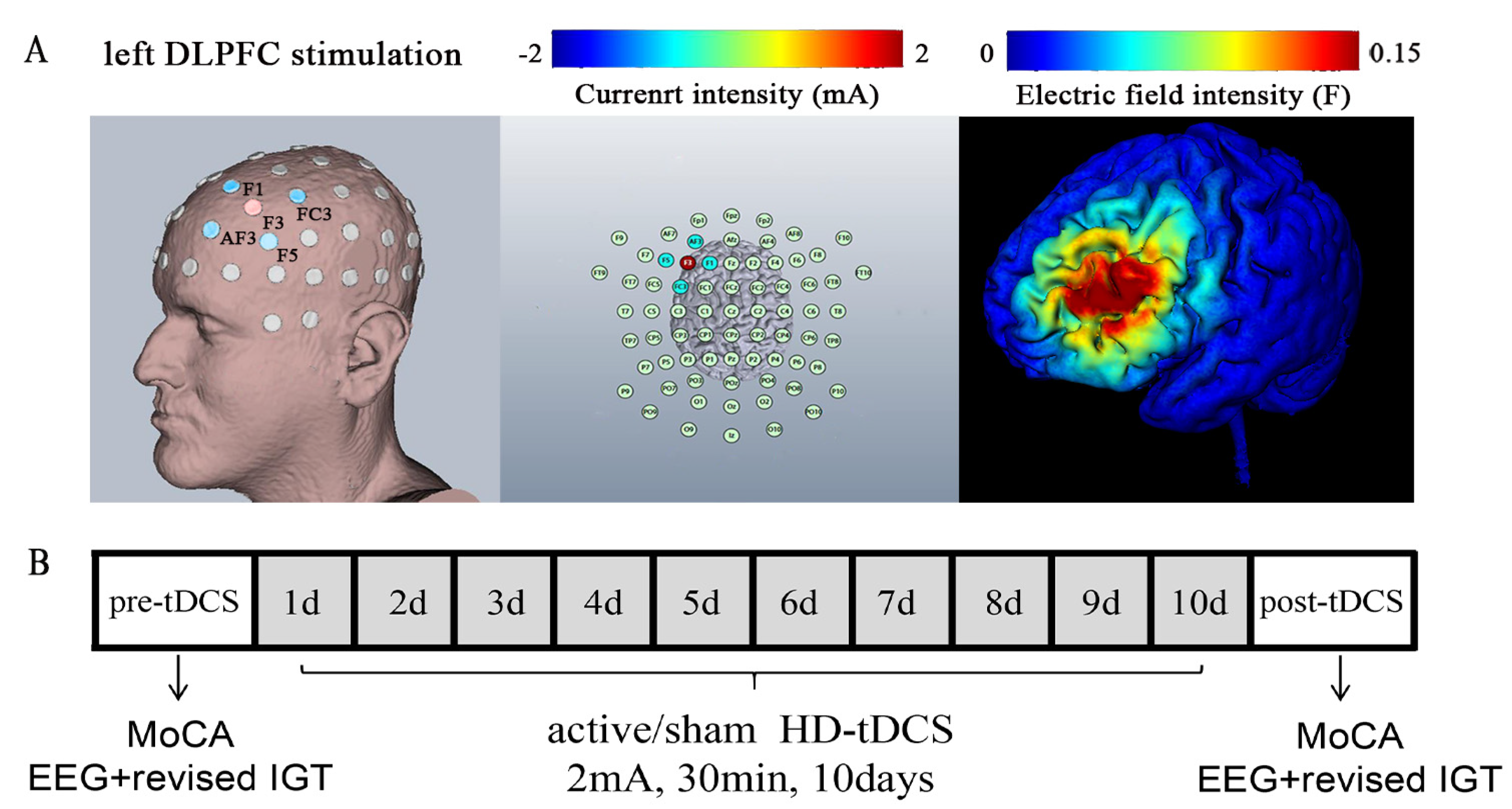
2.1. The Procedure
2.2. Participants
2.3. Task
2.4. High-Definition Transcranial Direct Current Stimulation
2.5. Electrophysiological Recording
2.6. Statistical Analysis
3. Results
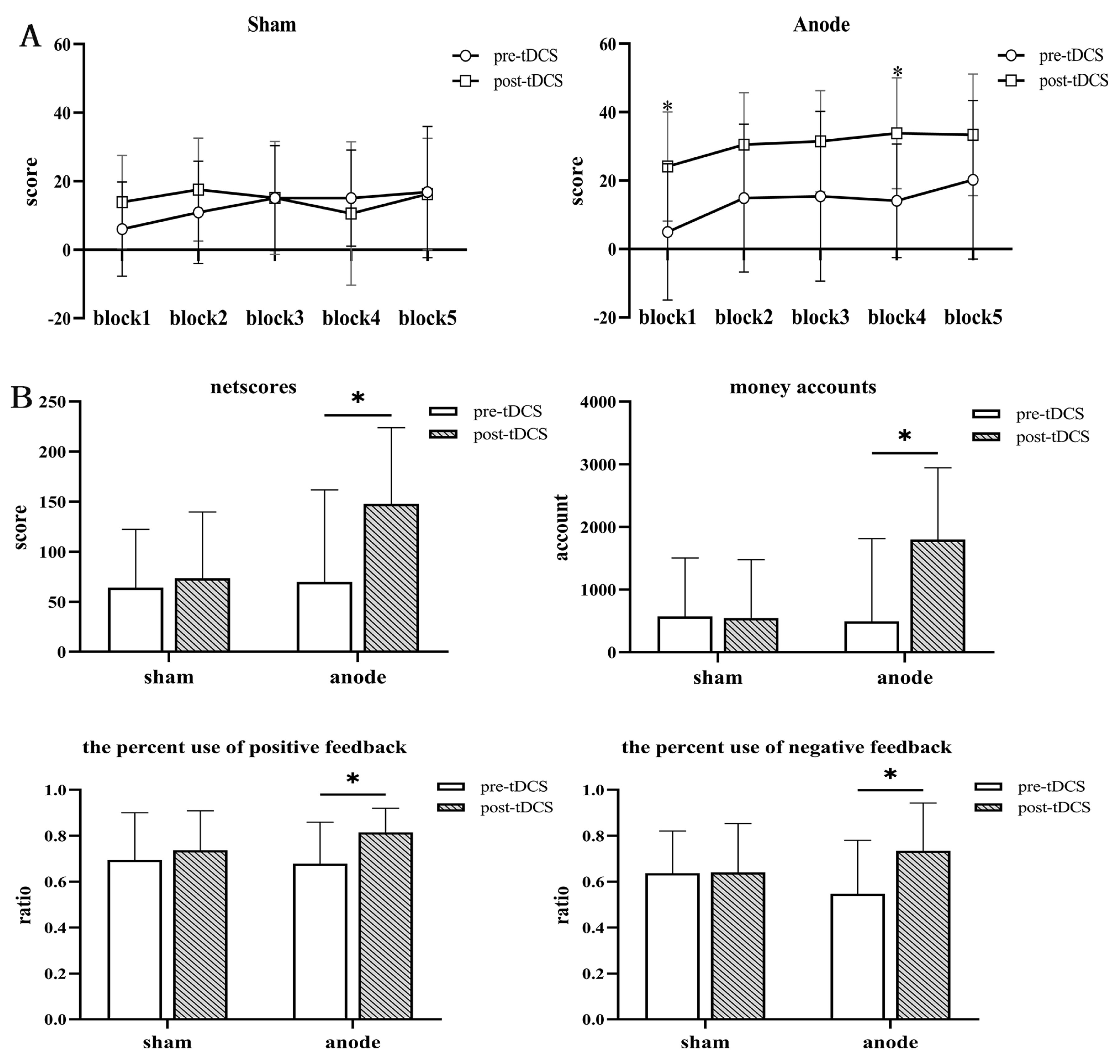
3.1. Demographic and Behavioral Performance
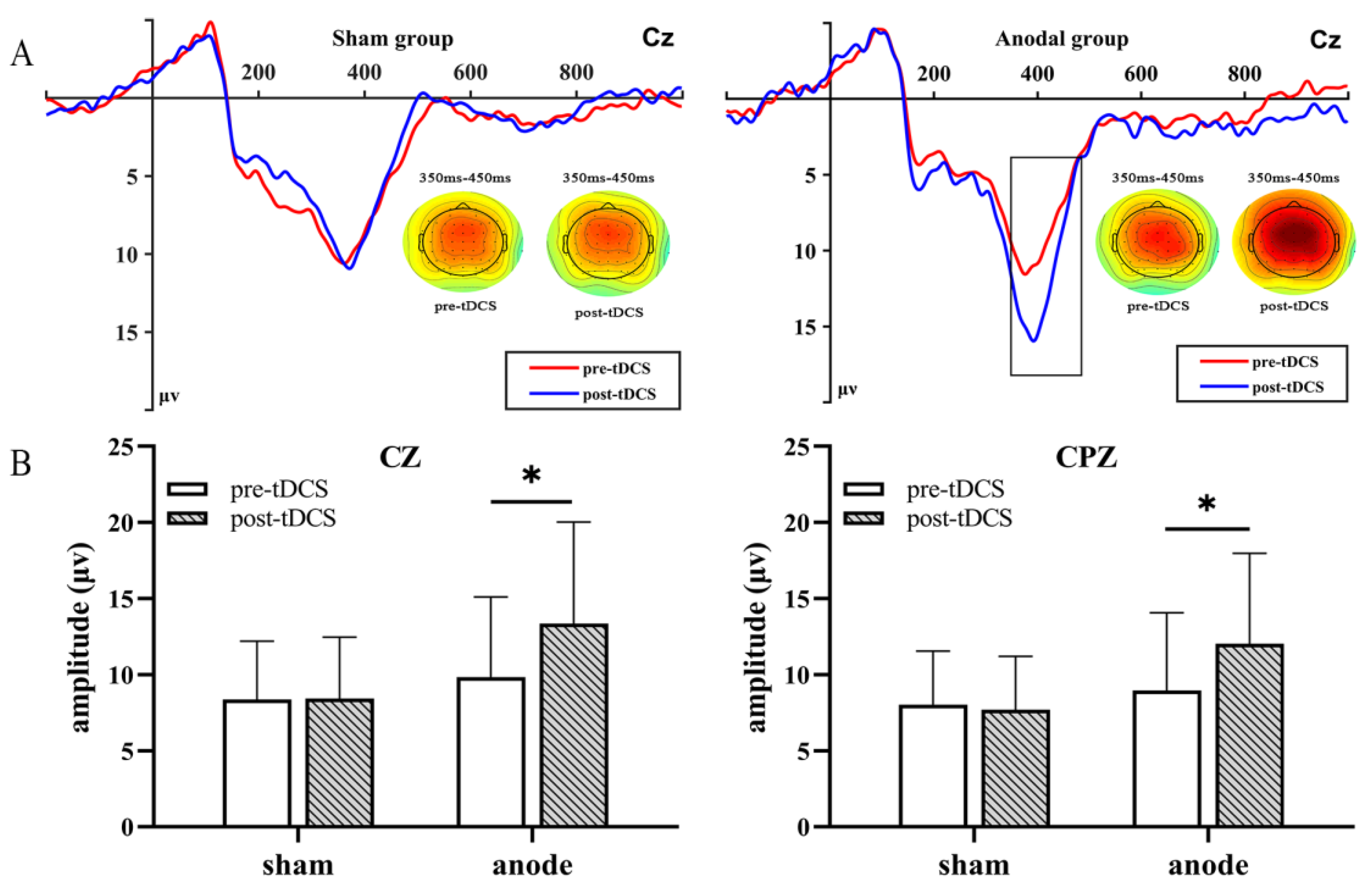
3.2. P300 Amplitude
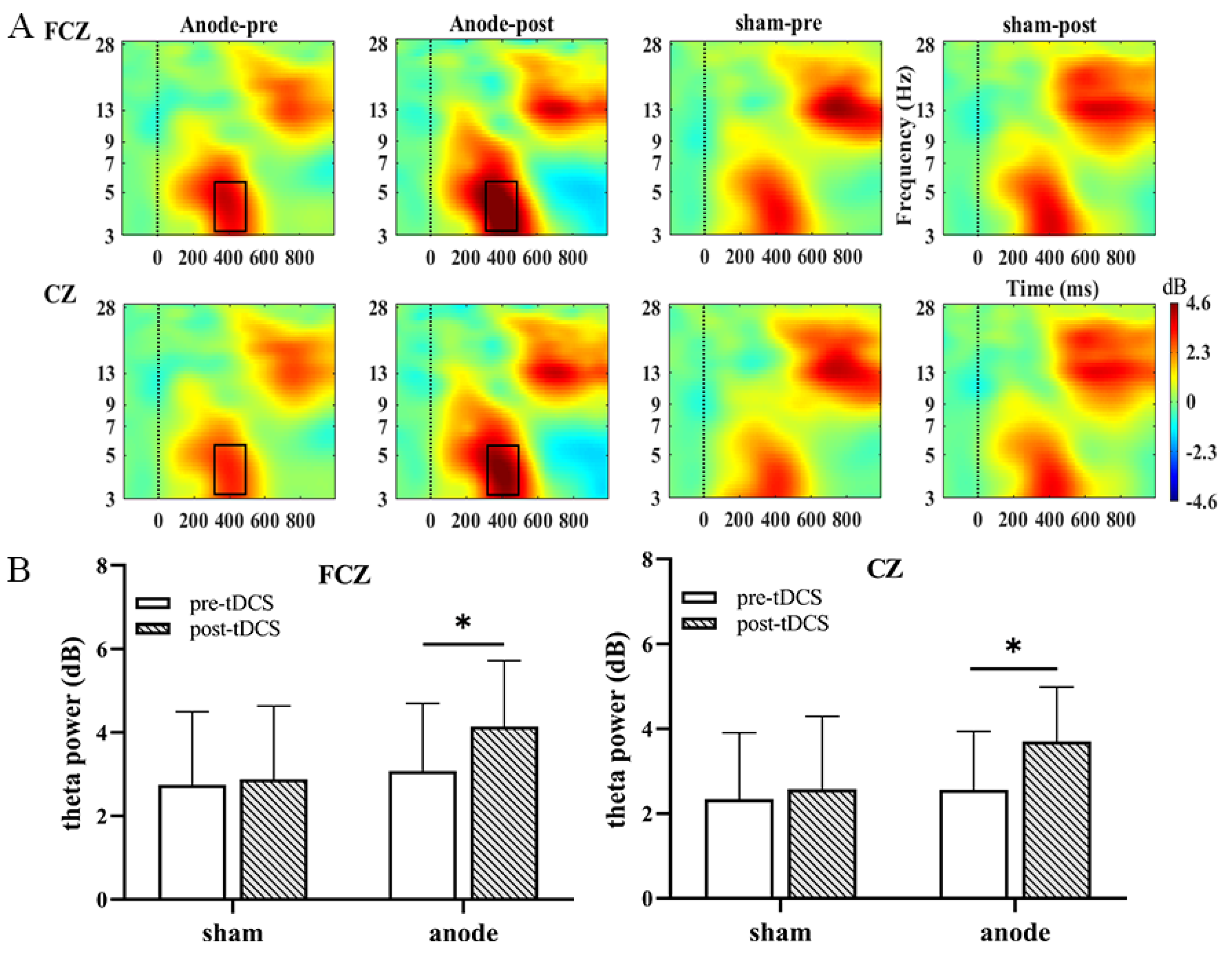
3.3. Theta Oscillations
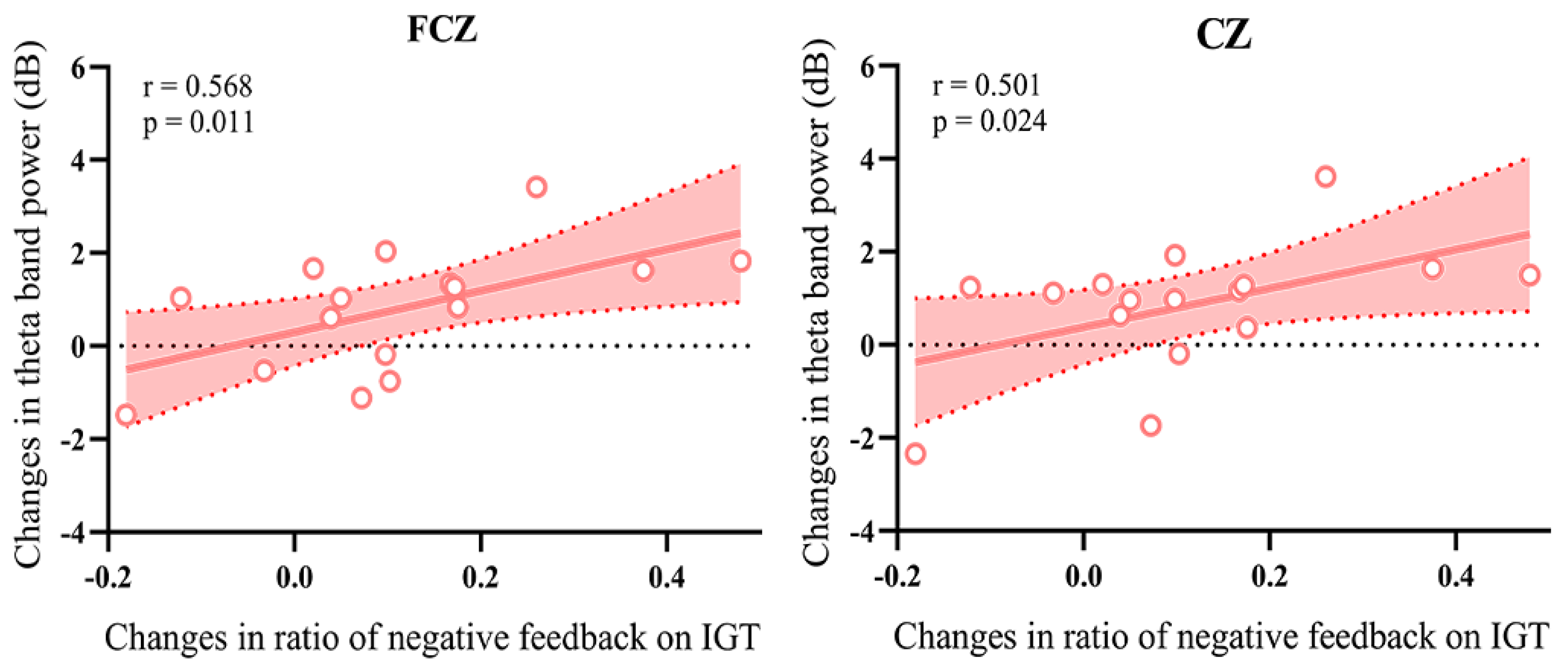
3.4. Correlation Analysis
3.5. Additional Analyses
3.6. Feasibility and Blinding Efficacy of the Procedure
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fecteau, S.; Pascual-Leone, A.; Zald, D.H.; Liguori, P.; Theoret, H.; Boggio, P.S.; Fregni, F. Activation of prefrontal cortex by transcranial direct current stimulation reduces appetite for risk during ambiguous decision making. J. Neurosci. 2007, 27, 6212–6218. [Google Scholar] [CrossRef]
- Liu, Y.W.; Chen, Z.H.; Luo, J.; Yin, M.Y.; Li, L.L.; Yang, Y.D.; Zheng, H.Q.; Liang, Z.H.; Hu, X.Q. Explore combined use of transcranial direct current stimulation and cognitive training on executive function after stroke. J. Rehabil. Med. 2021, 53, m162. [Google Scholar]
- Wang, L.; Wu, X.; Ji, G.J.; Xiao, G.; Xu, F.; Yan, Y.; Wu, Y.; Xi, C.; Chen, X.; Wang, K. Better modulation for risk decision-making after optimized magnetic stimulation. J. Neurosci. Res. 2021, 99, 858–871. [Google Scholar] [CrossRef] [PubMed]
- Soyata, A.Z.; Aksu, S.; Woods, A.J.; İşçen, P.; Saçar, K.T.; Karamürsel, S. Effect of transcranial direct current stimulation on decision making and cognitive flexibility in gambling disorder. Eur. Arch. Psychiatry Clin. Neurosci. 2019, 269, 275–284. [Google Scholar] [CrossRef] [PubMed]
- Romero Lauro, L.J.; Rosanova, M.; Mattavelli, G.; Convento, S.; Pisoni, A.; Opitz, A.; Bolognini, N.; Vallar, G. TDCS increases cortical excitability: Direct evidence from TMS–EEG. Cortex 2014, 58, 99–111. [Google Scholar] [CrossRef]
- Guo, H.; Zhang, Z.; Da, S.; Sheng, X.; Zhang, X. High-definition transcranial direct current stimulation (HD-tDCS) of left dorsolateral prefrontal cortex affects performance in Balloon Analogue Risk Task (BART). Brain Behav. 2018, 8, e884. [Google Scholar] [CrossRef] [PubMed]
- Brunoni, A.R.; Nitsche, M.A.; Bolognini, N.; Bikson, M.; Wagner, T.; Merabet, L.; Edwards, D.J.; Valero-Cabre, A.; Rotenberg, A.; Pascual-Leone, A.; et al. Clinical research with transcranial direct current stimulation (tDCS): Challenges and future directions. Brain Stimul. 2012, 5, 175–195. [Google Scholar] [CrossRef] [Green Version]
- Nitsche, M.A.; Paulus, W. Excitability changes induced in the human motor cortex by weak transcranial direct current stimulation. J. Physiol. 2000, 527, 633–639. [Google Scholar] [CrossRef]
- McGuire, J.T.; Botvinick, M.M. Prefrontal cortex, cognitive control, and the registration of decision costs. Proc. Natl. Acad. Sci. USA 2010, 107, 7922–7926. [Google Scholar] [CrossRef] [Green Version]
- Lu, H.; Gong, Y.; Huang, P.; Zhang, Y.; Guo, Z.; Zhu, X.; You, X. Effect of Repeated Anodal HD-tDCS on Executive Functions: Evidence from a Pilot and Single-Blinded fNIRS Study. Front. Human Neurosci. 2021, 14, 583730. [Google Scholar] [CrossRef]
- Hogeveen, J.; Grafman, J.; Aboseria, M.; David, A.; Bikson, M.; Hauner, K.K. Effects of High-Definition and Conventional tDCS on Response Inhibition. Brain Stimul. 2016, 9, 720–729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patel, H.; Naish, K.; Soreni, N.; Amlung, M. The Effects of a Single Transcranial Direct Current Stimulation Session on Impulsivity and Risk Among a Sample of Adult Recreational Cannabis Users. Front. Human Neurosci. 2022, 16, 758285. [Google Scholar] [CrossRef] [PubMed]
- Russo, R.; Twyman, P.; Cooper, N.R.; Fitzgerald, P.B.; Wallace, D. When you can, scale up: Large-scale study shows no effect of tDCS in an ambiguous risk-taking task. Neuropsychologia 2017, 104, 133–143. [Google Scholar] [CrossRef] [PubMed]
- Dubreuil-Vall, L.; Gomez-Bernal, F.; Villegas, A.C.; Cirillo, P.; Surman, C.; Ruffini, G.; Widge, A.S.; Camprodon, J.A. Transcranial Direct Current Stimulation to the Left Dorsolateral Prefrontal Cortex Improves Cognitive Control in Patients with Attention-Deficit/Hyperactivity Disorder: A Randomized Behavioral and Neurophysiological Study. Biol. Psychiatry Cogn. Neurosci. Neuroimaging 2021, 6, 439–448. [Google Scholar] [CrossRef]
- Solomons, C.D.; Shanmugasundaram, V. Transcranial direct current stimulation: A review of electrode characteristics and materials. Med. Eng. Phys. 2020, 85, 63–74. [Google Scholar] [CrossRef]
- Song, S.; Zilverstand, A.; Gui, W.; Li, H.; Zhou, X. Effects of single-session versus multi-session non-invasive brain stimulation on craving and consumption in individuals with drug addiction, eating disorders or obesity: A meta-analysis. Brain Stimul. 2019, 12, 606–618. [Google Scholar] [CrossRef] [PubMed]
- Filmer, H.L.; Varghese, E.; Hawkins, G.E.; Mattingley, J.B.; Dux, P.E. Improvements in Attention and Decision-Making Following Combined Behavioral Training and Brain Stimulation. Cereb. Cortex 2016, 27, 3675–3682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horvath, J.C.; Forte, J.D.; Carter, O. Quantitative Review Finds No Evidence of Cognitive Effects in Healthy Populations From Single-session Transcranial Direct Current Stimulation (tDCS). Brain Stimul. 2015, 8, 535–550. [Google Scholar] [CrossRef]
- Molavi, P.; Aziziaram, S.; Basharpoor, S.; Atadokht, A.; Nitsche, M.A.; Salehinejad, M.A. Repeated transcranial direct current stimulation of dorsolateral-prefrontal cortex improves executive functions, cognitive reappraisal emotion regulation, and control over emotional processing in borderline personality disorder: A randomized, sham-controlled, parallel-group study. J. Affect. Disord. 2020, 274, 93–102. [Google Scholar]
- Datta, A.; Bansal, V.; Diaz, J.; Patel, J.; Reato, D.; Bikson, M. Gyri-precise head model of transcranial direct current stimulation: Improved spatial focality using a ring electrode versus conventional rectangular pad. Brain Stimul. 2009, 2, 201–207. [Google Scholar] [CrossRef] [Green Version]
- Edwards, D.; Cortes, M.; Datta, A.; Minhas, P.; Wassermann, E.M.; Bikson, M. Physiological and modeling evidence for focal transcranial electrical brain stimulation in humans: A basis for high-definition tDCS. Neuroimage 2013, 74, 266–275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuo, H.; Bikson, M.; Datta, A.; Minhas, P.; Paulus, W.; Kuo, M.; Nitsche, M.A. Comparing Cortical Plasticity Induced by Conventional and High-Definition 4 × 1 Ring tDCS: A Neurophysiological Study. Brain Stimul. 2013, 6, 644–648. [Google Scholar] [CrossRef]
- Wang, W.; Zhu, Y.; Wang, L.; Mu, L.; Zhu, L.; Ding, D.; Ren, Z.; Yang, D.; Tang, H.; Zhang, L.; et al. High-frequency repetitive transcranial magnetic stimulation of the left dorsolateral prefrontal cortex reduces drug craving and improves decision-making ability in methamphetamine use disorder. Psychiatry Res. 2022, 317, 114904. [Google Scholar] [CrossRef]
- Shao, R.; Sun, D.; Lee, T.M. The interaction of perceived control and Gambler’s fallacy in risky decision making: An fMRI study. Hum. Brain Mapp. 2016, 37, 1218–1234. [Google Scholar] [CrossRef]
- Ernst, M.; Paulus, M.P. Neurobiology of Decision Making: A Selective Review from a Neurocognitive and Clinical Perspective. Biol. Psychiatry 2005, 58, 597–604. [Google Scholar] [CrossRef]
- Blankenstein, N.E.; Peper, J.S.; Crone, E.A.; van Duijvenvoorde, A.C.K. Neural Mechanisms Underlying Risk and Ambiguity Attitudes. J. Cogn. Neurosci. 2017, 29, 1845–1859. [Google Scholar] [CrossRef] [Green Version]
- Khani, A.; Rainer, G. Neural and neurochemical basis of reinforcement-guided decision making. J. Neurophysiol. 2016, 116, 724–741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.; Gao, M.; Shi, J.; Ye, H.; Chen, S. Modulating the Activity of the DLPFC and OFC Has Distinct Effects on Risk and Ambiguity Decision-Making: A tDCS Study. Front. Psychol. 2017, 8, 1417. [Google Scholar] [CrossRef] [Green Version]
- Xue, G.; Juan, C.H.; Chang, C.F.; Lu, Z.L.; Dong, Q. Lateral prefrontal cortex contributes to maladaptive decisions. Proc. Natl. Acad. Sci. USA 2012, 109, 4401–4406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mengarelli, F.; Spoglianti, S.; Avenanti, A.; di Pellegrino, G. Cathodal tDCS Over the Left Prefrontal Cortex Diminishes Choice-Induced Preference Change. Cereb. Cortex 2015, 25, 1219–1227. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Xue, G.; Potenza, M.N.; Zhang, J.T.; Yao, Y.W.; Xia, C.C.; Lan, J.; Ma, S.S.; Fang, X.Y. Dissociable neural processes during risky decision-making in individuals with Internet-gaming disorder. Neuroimage Clin. 2017, 14, 741–749. [Google Scholar] [CrossRef] [PubMed]
- Manes, F.; Sahakian, B.; Clark, L.; Rogers, R.; Antoun, N.; Aitken, M.; Robbins, T. Decision-making processes following damage to the prefrontal cortex. Brain 2002, 125, 624–639. [Google Scholar] [CrossRef] [Green Version]
- Salatino, A.; Miccolis, R.; Gammeri, R.; Ninghetto, M.; Belli, F.; Nobili, M.; Mouraux, A.; Ricci, R. Improvement of Impulsivity and Decision Making by Transcranial Direct Current Stimulation of the Dorsolateral Prefrontal Cortex in a Patient with Gambling Disorder. J. Gambl. Stud. 2022, 38, 627–634. [Google Scholar] [CrossRef] [PubMed]
- Zhu, C.; Yu, F.; Ye, R.; Chen, X.; Dong, Y.; Li, D.; Zhang, L.; Li, D.; Wang, K. External error monitoring in subclinical obsessive-compulsive subjects: Electrophysiological evidence from a Gambling Task. PLoS ONE 2014, 9, e90874. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; Wang, L.; Yu, F.; Ji, G.J.; Xiao, G.; Feifei, X.; Chunyan, Z.; Xingui, C.; Wang, K. Intermittent Theta Burst Stimulation (iTBS) as an Optimal Treatment for Schizophrenia Risk Decision: An ERSP Study. Front. Psychiatry 2021, 12, 594102. [Google Scholar] [CrossRef]
- Li, X.; Lu, Z.; D’Argembeau, A.; Ng, M.; Bechara, A. The Iowa Gambling Task in fMRI images. Human Brain Mapp. 2009, 31, 410–423. [Google Scholar] [CrossRef] [Green Version]
- Ouerchefani, R.; Ouerchefani, N.; Allain, P.; Ben Rejeb, M.R.; Le Gall, D. Relationships between executive function, working memory, and decision-making on the Iowa Gambling Task: Evidence from ventromedial patients, dorsolateral patients, and normal subjects. J. Neuropsychol. 2018, 13, 432–461. [Google Scholar] [CrossRef] [PubMed]
- Gilmore, C.S.; Dickmann, P.J.; Nelson, B.G.; Lamberty, G.J.; Lim, K.O. Transcranial Direct Current Stimulation (tDCS) paired with a decision-making task reduces risk-taking in a clinically impulsive sample. Brain Stimul. 2018, 11, 302–309. [Google Scholar] [CrossRef]
- Ehrhardt, S.E.; Filmer, H.L.; Wards, Y.; Mattingley, J.B.; Dux, P.E. The influence of tDCS intensity on decision-making training and transfer outcomes. J. Neurophysiol. 2021, 125, 385–397. [Google Scholar] [CrossRef]
- Jiang, Y.; Cui, C.; Liu, M.; Zhang, X. Capture or suppression? Attentional allocation upon reward and loss-associated nonsalient distractors are supported by distinct neural mechanisms: An EEG study. Neuropsychologia 2021, 157, 107879. [Google Scholar] [CrossRef]
- Leicht, G.; Troschütz, S.; Andreou, C.; Karamatskos, E.; Ertl, M.; Naber, D.; Mulert, C.; Barnes, G.R. Relationship between oscillatory neuronal activity during reward processing and trait impulsivity and sensation seeking. PLoS ONE 2013, 8, e83414. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Y.; Yi, W.; Cheng, J.; Li, Q. Common and distinct electrophysiological correlates of feedback processing during risky and ambiguous decision making. Neuropsychologia 2020, 146, 107526. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Selva, J.M.; Muñoz, M.A.; Sánchez-Navarro, J.P.; Walteros, C.; Montoya, P. Time Course of the Neural Activity Related to Behavioral Decision-Making as Revealed by Event-Related Potentials. Front. Behav. Neurosci. 2019, 13, 191. [Google Scholar] [CrossRef] [Green Version]
- Cui, J.; Chen, Y.; Wang, Y.; Shum, D.H.K.; Chan, R.C.K. Neural correlates of uncertain decision making: ERP evidence from the Iowa Gambling Task. Front. Human Neurosci. 2013, 7, 776. [Google Scholar] [CrossRef] [Green Version]
- Luft, C.D.B.; Nolte, G.; Bhattacharya, J. High-Learners Present Larger Mid-Frontal Theta Power and Connectivity in Response to Incorrect Performance Feedback. J. Neurosci. 2013, 33, 2029–2038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christie, G.J.; Tata, M.S. Right frontal cortex generates reward-related theta-band oscillatory activity. Neuroimage 2009, 48, 415–422. [Google Scholar] [CrossRef]
- Minhas, P.; Bansal, V.; Patel, J.; Ho, J.S.; Diaz, J.; Datta, A.; Bikson, M. Electrodes for high-definition transcutaneous DC stimulation for applications in drug delivery and electrotherapy, including tDCS. J. Neurosci. Methods 2010, 190, 188–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Li, J. High-definition transcranial stimulation over the dorsolateral prefrontal cortex alters the sunk cost effect: A mental accounting framework. J. Neurosci. 2022, 42, 6770–6781. [Google Scholar] [CrossRef]
- Choi, H.; Park, J.; Yang, Y.M. Whitening Technique Based on Gram-Schmidt Orthogonalization for Motor Imagery Classification of Brain-Computer Interface Applications. Sensors 2022, 22, 6042. [Google Scholar] [CrossRef]
- Helfrich, R.F.; Knight, R.T. Cognitive neurophysiology: Event-related potentials. Handb. Clin. Neurol. 2019, 160, 543–558. [Google Scholar]
- Cui, T.; Wang, P.P.; Liu, S.; Zhang, X. P300 amplitude and latency in autism spectrum disorder: A meta-analysis. Eur. Child Adoles. Psy. 2017, 26, 177–190. [Google Scholar] [CrossRef]
- Rawls, E.; Miskovic, V.; Moody, S.N.; Lee, Y.; Shirtcliff, E.A.; Lamm, C. Feedback-Related Negativity and Frontal Midline Theta Reflect Dissociable Processing of Reinforcement. Front. Human Neurosci. 2020, 13, 452. [Google Scholar] [CrossRef] [Green Version]
- Correll, J.; Mellinger, C.; Pedersen, E.J. Flexible approaches for estimating partial eta squared in mixed-effects models with crossed random factors. Behav. Res. Methods 2022, 54, 1626–1642. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Lin, Y.; Gao, M.; Jin, X. Effect of Modulating Activity of DLPFC and Gender on Search Behavior: A tDCS Experiment. Front. Human Neurosci. 2018, 12, 325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ouellet, J.; McGirr, A.; Van den Eynde, F.; Jollant, F.; Lepage, M.; Berlim, M.T. Enhancing decision-making and cognitive impulse control with transcranial direct current stimulation (tDCS) applied over the orbitofrontal cortex (OFC): A randomized and sham-controlled exploratory study. J. Psychiatr. Res. 2015, 69, 27–34. [Google Scholar] [CrossRef]
- Paulus, M.P.; Rogalsky, C.; Simmons, A.; Feinstein, J.S.; Stein, M.B. Increased activation in the right insula during risk-taking decision making is related to harm avoidance and neuroticism. Neuroimage 2003, 19, 1439–1448. [Google Scholar] [CrossRef]
- Hsu, M.; Bhatt, M.; Adolphs, R.; Tranel, D.; Camerer, C.F. Neural systems responding to degrees of uncertainty in human decision-making. Science 2005, 310, 1680–1683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramayya, A.G.; Pedisich, I.; Kahana, M.J. Expectation modulates neural representations of valence throughout the human brain. Neuroimage 2015, 115, 214–223. [Google Scholar] [CrossRef] [Green Version]
- Weber, M.J.; Messing, S.B.; Rao, H.; Detre, J.A.; Thompson Schill, S.L. Prefrontal transcranial direct current stimulation alters activation and connectivity in cortical and subcortical reward systems: A tDCS-fMRI study. Human Brain Mapp. 2014, 35, 3673–3686. [Google Scholar] [CrossRef] [Green Version]
- Benussi, A.; Alberici, A.; Cantoni, V.; Manenti, R.; Brambilla, M.; Dell’Era, V.; Gazzina, S.; Manes, M.; Cristillo, V.; Padovani, A.; et al. Modulating risky decision-making in Parkinson’s disease by transcranial direct current stimulation. Eur. J. Neurol. 2017, 24, 751–754. [Google Scholar] [CrossRef]
- Ashok, A.H.; Marques, T.R.; Jauhar, S.; Nour, M.M.; Goodwin, G.M.; Young, A.H.; Howes, O.D. The dopamine hypothesis of bipolar affective disorder: The state of the art and implications for treatment. Mol. Psychiatry 2017, 22, 666–679. [Google Scholar] [CrossRef] [PubMed]
- Bunai, T.; Hirosawa, T.; Kikuchi, M.; Fukai, M.; Yokokura, M.; Ito, S.; Takata, Y.; Terada, T.; Ouchi, Y. tDCS-induced modulation of GABA concentration and dopamine release in the human brain: A combination study of magnetic resonance spectroscopy and positron emission tomography. Brain Stimul. 2021, 14, 154–160. [Google Scholar] [CrossRef] [PubMed]
- Mendes, A.J.; Pacheco-Barrios, K.; Lema, A.; Goncalves, O.F.; Fregni, F.; Leite, J.; Carvalho, S. Modulation of the cognitive event-related potential P3 by transcranial direct current stimulation: Systematic review and meta-analysis. Neurosci. Biobehav. Rev. 2022, 132, 894–907. [Google Scholar] [CrossRef]
- Wu, Y.; Zhou, X. The P300 and reward valence, magnitude, and expectancy in outcome evaluation. Brain Res. 2009, 1286, 114–122. [Google Scholar] [CrossRef]
- Glazer, J.E.; Kelley, N.J.; Pornpattananangkul, N.; Mittal, V.A.; Nusslock, R. Beyond the FRN: Broadening the time-course of EEG and ERP components implicated in reward processing. Int. J. Psychophysiol. 2018, 132, 184–202. [Google Scholar] [CrossRef]
- Delorme, A.; Makeig, S. EEGLAB: An open source toolbox for analysis of single-trial EEG dynamics including independent component analysis. J. Neurosci. Meth. 2004, 134, 9–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Possti, D.; Fahoum, F.; Sosnik, R.; Giladi, N.; Hausdorff, J.M.; Mirelman, A.; Maidan, I. Changes in the EEG spectral power during dual-task walking with aging and Parkinson’s disease: Initial findings using Event-Related Spectral Perturbation analysis. J. Neurol. 2021, 268, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Gao, J.; Zhu, C. Abnormal negative feedback processing in individuals with autistic traits in the Iowa gambling task: Evidence from behavior and event-related potentials. Int. J. Psychophysiol. 2021, 165, 36–46. [Google Scholar] [CrossRef]
- Avvaru, S.; Peled, N.; Provenza, N.R.; Widge, A.S.; Parhi, K.K. Region-Level Functional and Effective Network Analysis of Human Brain During Cognitive Task Engagement. IEEE Trans. Neural Syst. Rehabil. Eng. 2021, 29, 1651–1660. [Google Scholar] [CrossRef]
- van der Molen, M.; Dekkers, L.; Westenberg, P.M.; van der Veen, F.M.; van der Molen, M.W. Why don’t you like me? Midfrontal theta power in response to unexpected peer rejection feedback. Neuroimage 2017, 146, 474–483. [Google Scholar] [CrossRef] [Green Version]
- van de Vijver, I.; van Driel, J.; Hillebrand, A.; Cohen, M.X. Interactions between frontal and posterior oscillatory dynamics support adjustment of stimulus processing during reinforcement learning. Neuroimage 2018, 181, 170–181. [Google Scholar] [CrossRef] [PubMed]
- van den Bos, R.; Homberg, J.; de Visser, L. A critical review of sex differences in decision-making tasks: Focus on the Iowa Gambling Task. Behav. Brain Res. 2013, 238, 95–108. [Google Scholar] [CrossRef] [PubMed]
- León, J.J.; Sánchez-Kuhn, A.; Fernández-Martín, P.; Páez-Pérez, M.A.; Thomas, C.; Datta, A.; Sánchez-Santed, F.; Flores, P. Transcranial direct current stimulation improves risky decision making in women but not in men: A sham-controlled study. Behav. Brain Res. 2020, 382, 112485. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Active Group (n = 16) Mean (SD) | Sham Group (n = 17) Mean (SD) | Baseline Comparison | ||||
|---|---|---|---|---|---|---|
| Pre | Post | Pre | Post | χ2/z/t | p | |
| Age (years) | 23.31 (3.26) | - | 21.88 (2.26) | - | −1.300 b | 0.194 |
| Education(years) | 15.81 (1.94) | - | 15.76 (1.86) | - | −0.112 b | 0.911 |
| MoCA | 28.56 (0.96) | 28.94 (1.06) | 28.76 (1.03) | 28.82 (1.19) | −0.601 b | 0.548 |
| Sex(male/female) | 7/9 | - | 10/7 | - | 0.750 a | 0.387 |
| Total netscores | 69.63 (92.13) | 153.38 (72.64) | 63.88 (58.52) | 73.41 (66.31) | 0.215 c | 0.831 |
| Money accounts | 493.75 (1320.09) | 1796.88 (1145.78) | 570.59 (934.25) | 544.12 (933.05) | −0.194 c | 0.847 |
| Ratio of positive feedback | 0.68 (0.18) | 0.81 (0.11) | 0.70 (0.20) | 0.74 (0.17) | −0.259 c | 0.798 |
| Ratio of negative feedback | 0.55 (0.23) | 0.73 (0.21) | 0.64 (0.18) | 0.64 (0.21) | −1.228 c | 0.229 |
| Active Group (n = 16, 7 Males) Mean (SD) | Sham Group (n = 17, 10 Males) Mean (SD) | Factor Time | Effect Size | Group by Time Interaction | Effect Size | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Pre | Post | Pre | Post | F | p a | F | p a | |||
| P300 amplitude | ||||||||||
| FCZ100 | 10.08 (5.90) | 14.04 (7.15) | 8.90 (4.65) | 9.19 (4.80) | 6.581 | 0.015 * | 0.175 | 4.925 | 0.034 * | 0.137 |
| FCZ101 | 9.51 (4.37) | 11.12 (5.17) | 8.07 (5.07) | 9.32 (4.57) | 3.355 | 0.077 | 0.098 | 0.052 | 0.822 | 0.002 |
| FCZ50 | 8.80 (5.91) | 10.51 (8.06) | 6.76 (5.14) | 8.92 (5.01) | 6.050 | 0.020 * | 0.163 | 0.082 | 0.776 | 0.003 |
| FCZ51 | 6.73 (3.68) | 7.46 (4.46) | 6.01 (4.04) | 6.40 (4.18) | 0.829 | 0.370 | 0.026 | 0.073 | 0.789 | 0.002 |
| CZ100 | 9.84 (5.25) | 13.36 (6.67) | 8.36 (3.84) | 8.43 (4.03) | 5.910 | 0.021 * | 0.160 | 5.460 | 0.026 * | 0.150 |
| CZ101 | 9.98 (4.36) | 11.17 (4.85) | 8.59 (4.92) | 9.49 (4.83) | 1.756 | 0.195 | 0.054 | 0.035 | 0.854 | 0.001 |
| CZ50 | 8.37 (5.67) | 9.82 (8.03) | 6.45 (4.59) | 8.20 (4.58) | 4.224 | 0.048 * | 0.120 | 0.036 | 0.850 | 0.001 |
| CZ51 | 7.17 (3.78) | 7.61 (4.40) | 6.28 (3.99) | 6.28 (4.05) | 0.147 | 0.704 | 0.005 | 0.148 | 0.703 | 0.005 |
| CPZ100 | 8.97 (5.10) | 12.04 (5.94) | 8.02 (3.52) | 7.70 (3.49) | 3.636 | 0.066 | 0.105 | 5.525 | 0.025 * | 0.151 |
| CPZ101 | 9.34 (4.29) | 10.80 (4.82) | 8.74 (4.32) | 9.42 (4.87) | 2.035 | 0.164 | 0.062 | 0.265 | 0.610 | 0.008 |
| CPZ50 | 7.44 (5.95) | 9.02 (6.77) | 6.42 (4.26) | 7.11 (4.04) | 2.571 | 0.119 | 0.077 | 0.389 | 0.538 | 0.012 |
| CPZ51 | 6.95 (4.29) | 7.75 (4.26) | 6.60 (3.77) | 6.08 (3.76) | 0.065 | 0.801 | 0.002 | 1.422 | 0.242 | 0.044 |
| Active Group (n = 16, 7 Males) Mean (SD) | Sham Group (n = 17, 10 Males) Mean (SD) | Factor Time | Effect Size | Group by Time Interaction | Effect Size | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Pre | Post | Pre | Post | F | p a | F | p a | |||
| Theta oscillation power | ||||||||||
| FCZ100 | 3.08 (1.62) | 4.14 (1.58) | 2.74 (1.75) | 2.88 (1.75) | 6.797 | 0.014 * | 0.180 | 4.026 | 0.054 | 0.115 |
| FCZ101 | 1.18 (1.53) | 1.61 (2.28) | 1.17 (1.58) | 1.76 (1.41) | 2.638 | 0.114 | 0.078 | 0.060 | 0.808 | 0.002 |
| FCZ50 | 2.49 (2.09) | 3.43 (2.11) | 2.30 (1.65) | 2.30 (1.99) | 2.819 | 0.103 | 0.083 | 2.902 | 0.098 | 0.086 |
| FCZ51 | 1.08 (1.43) | 1.47 (1.13) | 0.59 (0.58) | 1.10 (1.37) | 5.956 | 0.021 * | 0.161 | 0.112 | 0.740 | 0.004 |
| CZ100 | 2.56 (1.37) | 3.70 (1.28) | 2.34 (1.56) | 2.57 (1.72) | 13.227 | 0.001 ** | 0.300 | 5.700 | 0.023 * | 0.155 |
| CZ101 | 1.49 (1.57) | 1.73 (2.48) | 1.51 (1.85) | 1.97 (1.46) | 1.093 | 0.304 | 0.034 | 0.103 | 0.750 | 0.003 |
| CZ50 | 2.13 (1.89) | 3.11 (2.02) | 1.96 (1.37) | 2.30 (1.99) | 5.290 | 0.028 * | 0.146 | 1.268 | 0.269 | 0.039 |
| CZ51 | 1.24 (1.48) | 1.59 (1.10) | 0.90 (0.82) | 1.12 (1.33) | 2.702 | 0.110 | 0.080 | 0.144 | 0.707 | 0.005 |
| CPZ100 | 1.96 (1.57) | 3.06 (1.29) | 1.93 (1.60) | 2.09 (1.49) | 7.054 | 0.012 * | 0.185 | 3.853 | 0.059 | 0.111 |
| CPZ101 | 1.67 (1.48) | 2.10 (2.29) | 1.56 (1.86) | 2.16 (1.54) | 2.792 | 0.105 | 0.083 | 0.071 | 0.792 | 0.002 |
| CPZ50 | 1.81 (1.70) | 2.62 (1.74) | 1.62 (1.24) | 1.74 (1.71) | 2.764 | 0.106 | 0.082 | 1.504 | 0.229 | 0.046 |
| CPZ51 | 1.13 (1.46) | 1.77 (1.11) | 1.09 (0.95) | 1.34 (1.29) | 7.974 | 0.008 ** | 0.205 | 1.495 | 0.231 | 0.046 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, Y.; Xiao, G.; Chen, Q.; Wang, Y.; Wang, L.; Xie, C.; Wang, K.; Chen, X. High-Definition Transcranial Direct Current Stimulation Improves Decision-Making Ability: A Study Based on EEG. Brain Sci. 2023, 13, 640. https://doi.org/10.3390/brainsci13040640
Zhou Y, Xiao G, Chen Q, Wang Y, Wang L, Xie C, Wang K, Chen X. High-Definition Transcranial Direct Current Stimulation Improves Decision-Making Ability: A Study Based on EEG. Brain Sciences. 2023; 13(4):640. https://doi.org/10.3390/brainsci13040640
Chicago/Turabian StyleZhou, Yuwei, Guixian Xiao, Qing Chen, Yuyang Wang, Lu Wang, Chengjuan Xie, Kai Wang, and Xingui Chen. 2023. "High-Definition Transcranial Direct Current Stimulation Improves Decision-Making Ability: A Study Based on EEG" Brain Sciences 13, no. 4: 640. https://doi.org/10.3390/brainsci13040640