
Articulation-Function-Associated Cortical Developmental Changes in Patients with Cleft Lip and Palate
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. MRI Acquisition and Preprocessing
2.3. Volumetric and Cortical Reconstruction for T1w Imaging Data
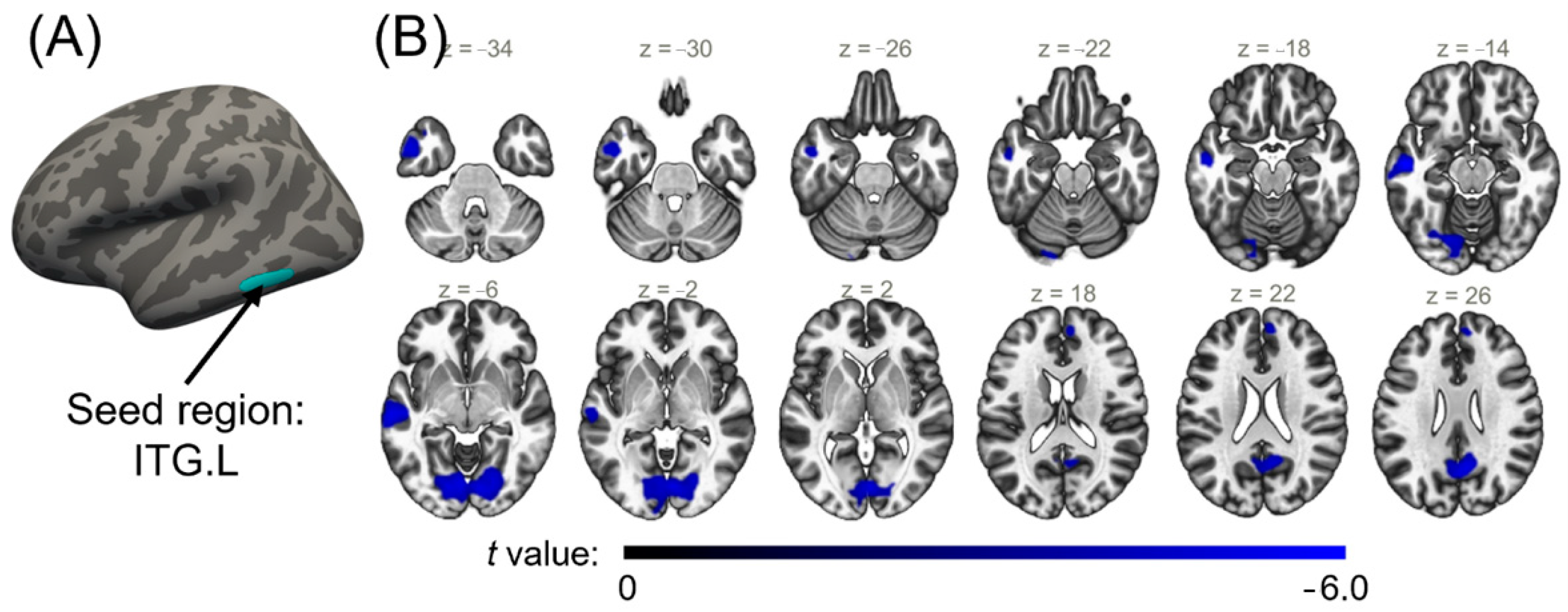
2.4. Seed-Based Functional Connectivity Analysis for Cortical Regions with a Significant Interaction Effect
2.5. Statistical Analysis
3. Results
3.1. Demographics
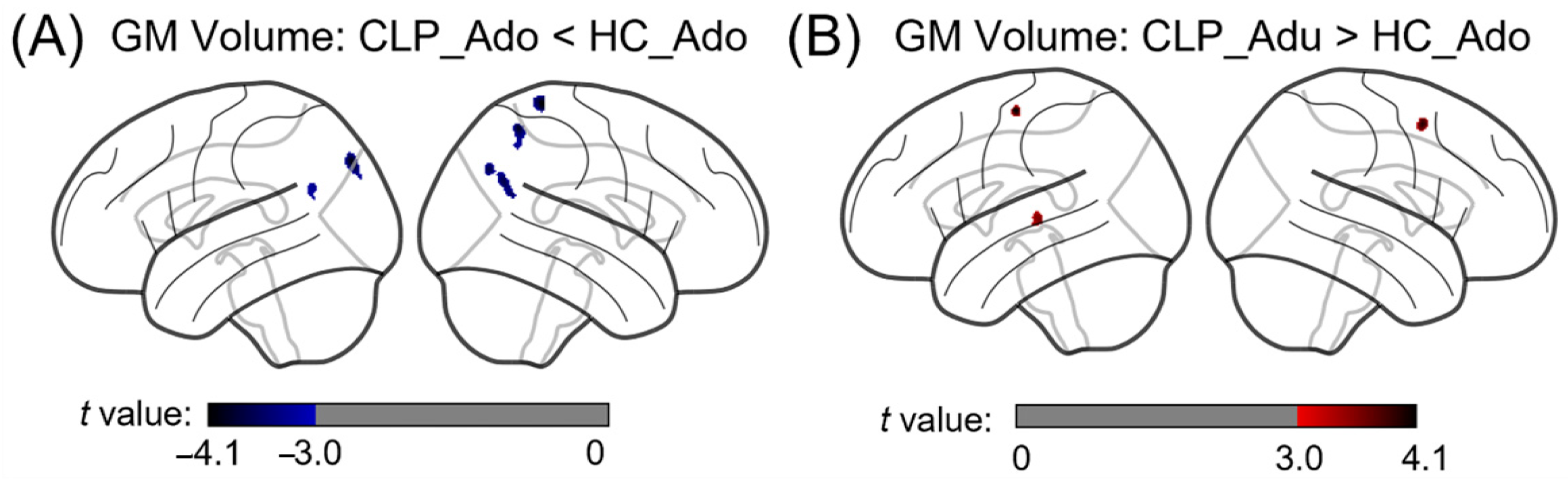
3.2. The Differences in Gray Matter Volume between Cleft Lip with Palate (CLP) Participants and Healthy Controls across Development
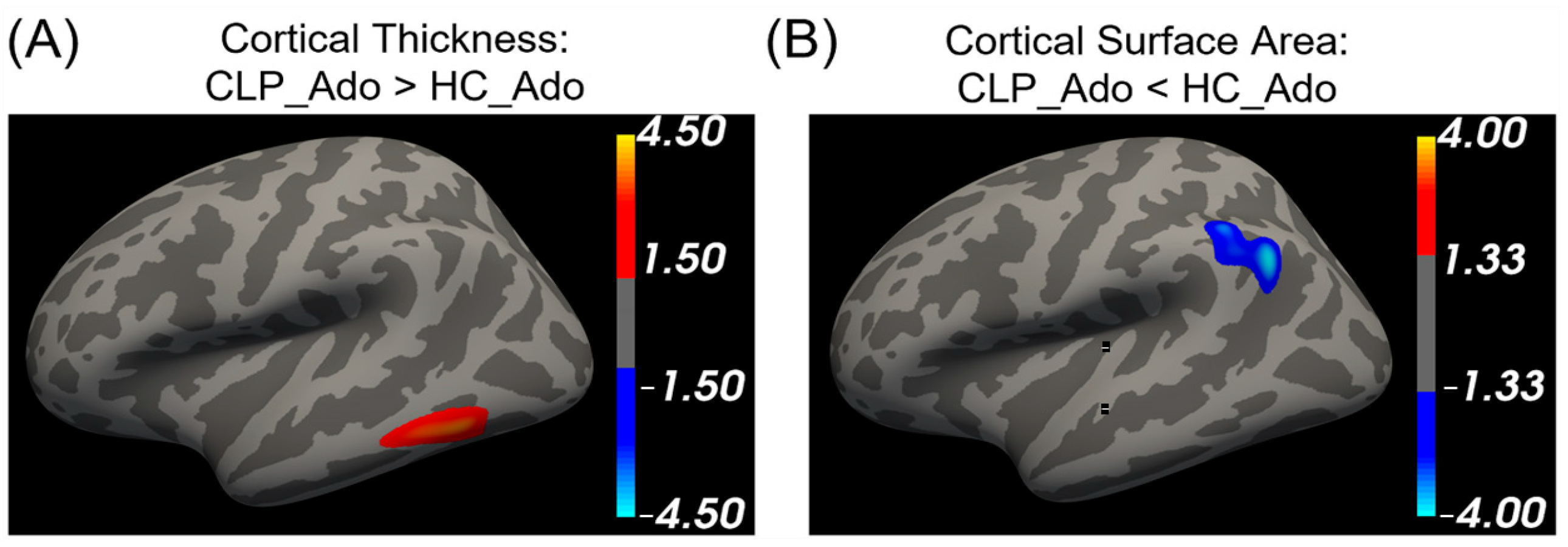
3.3. The Differences in Cortical Thickness and Cortical Surface between CLP and Healthy Controls across Development
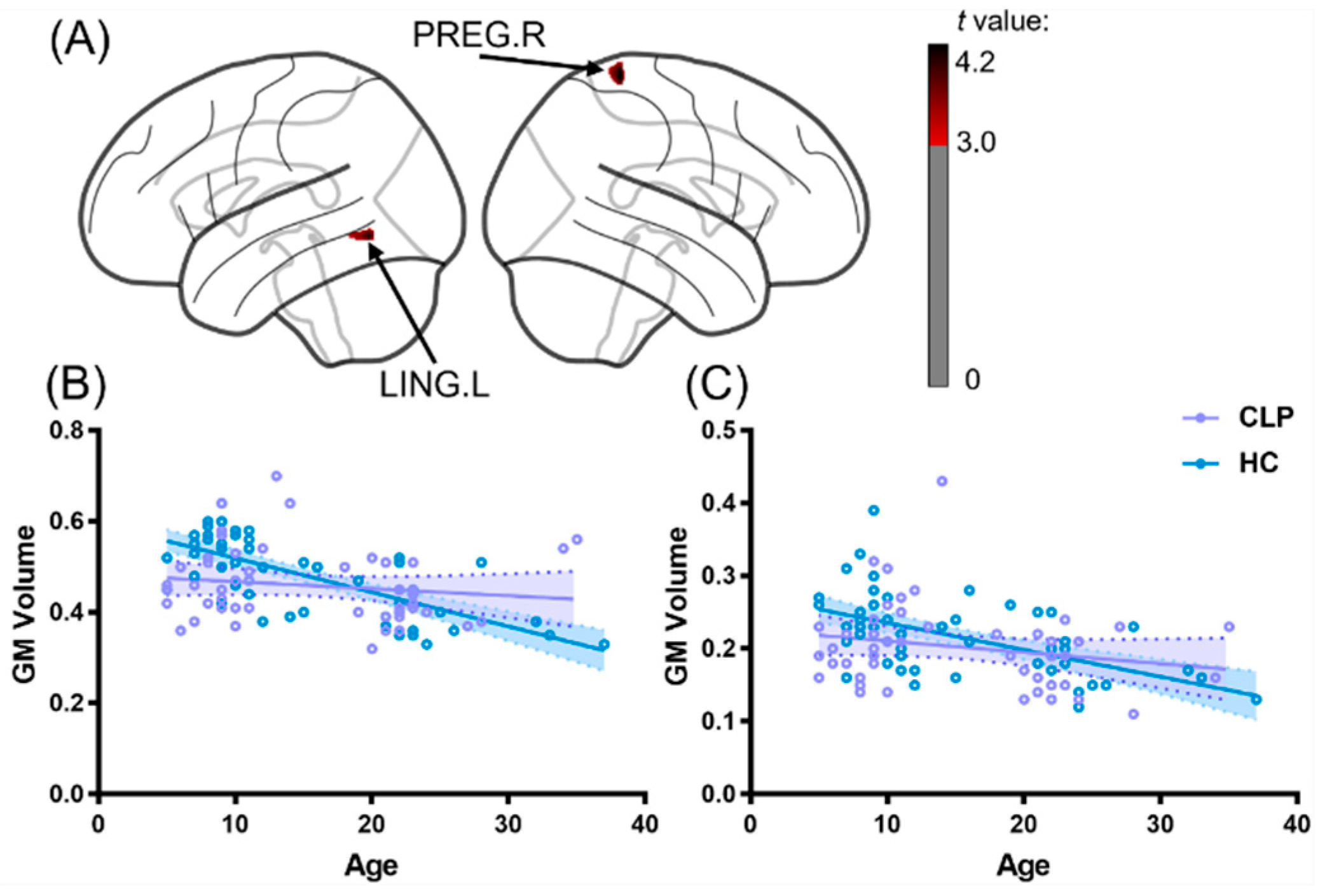
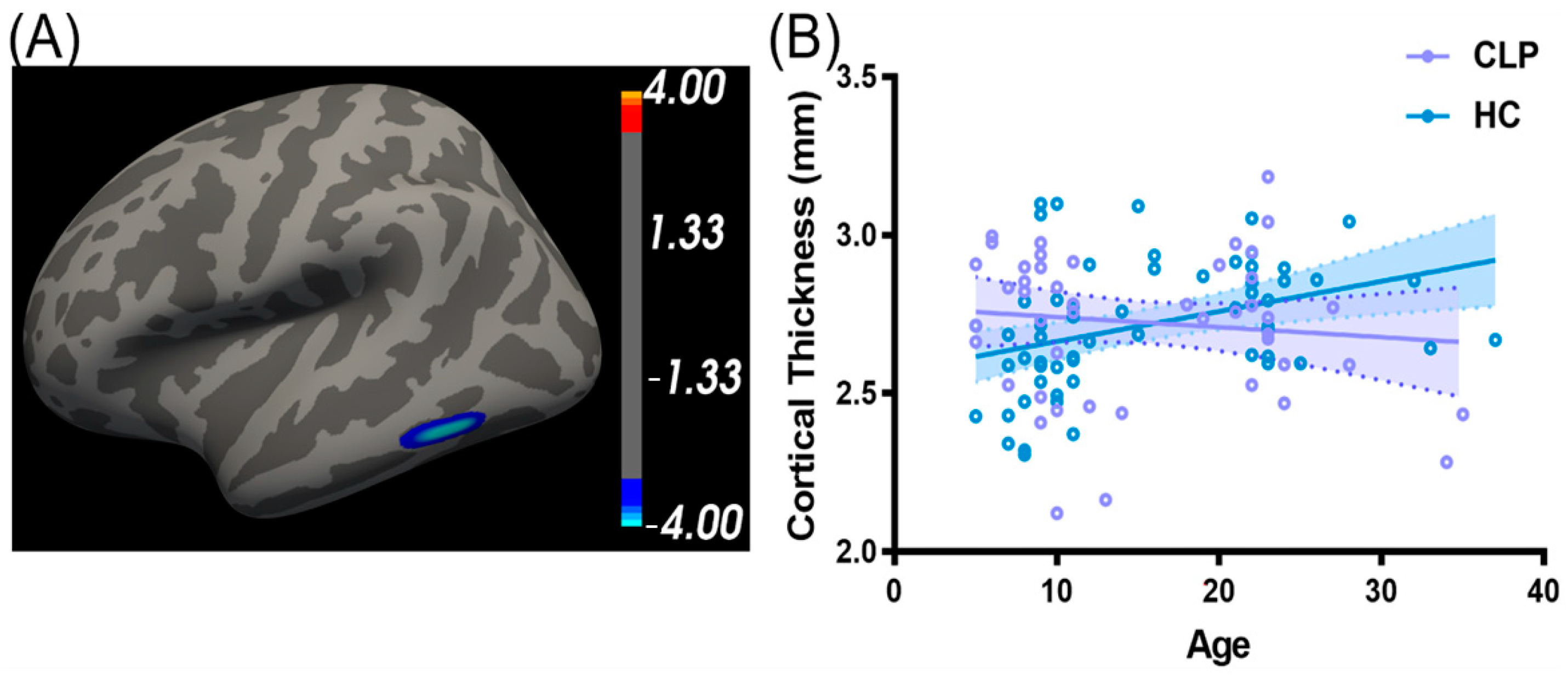
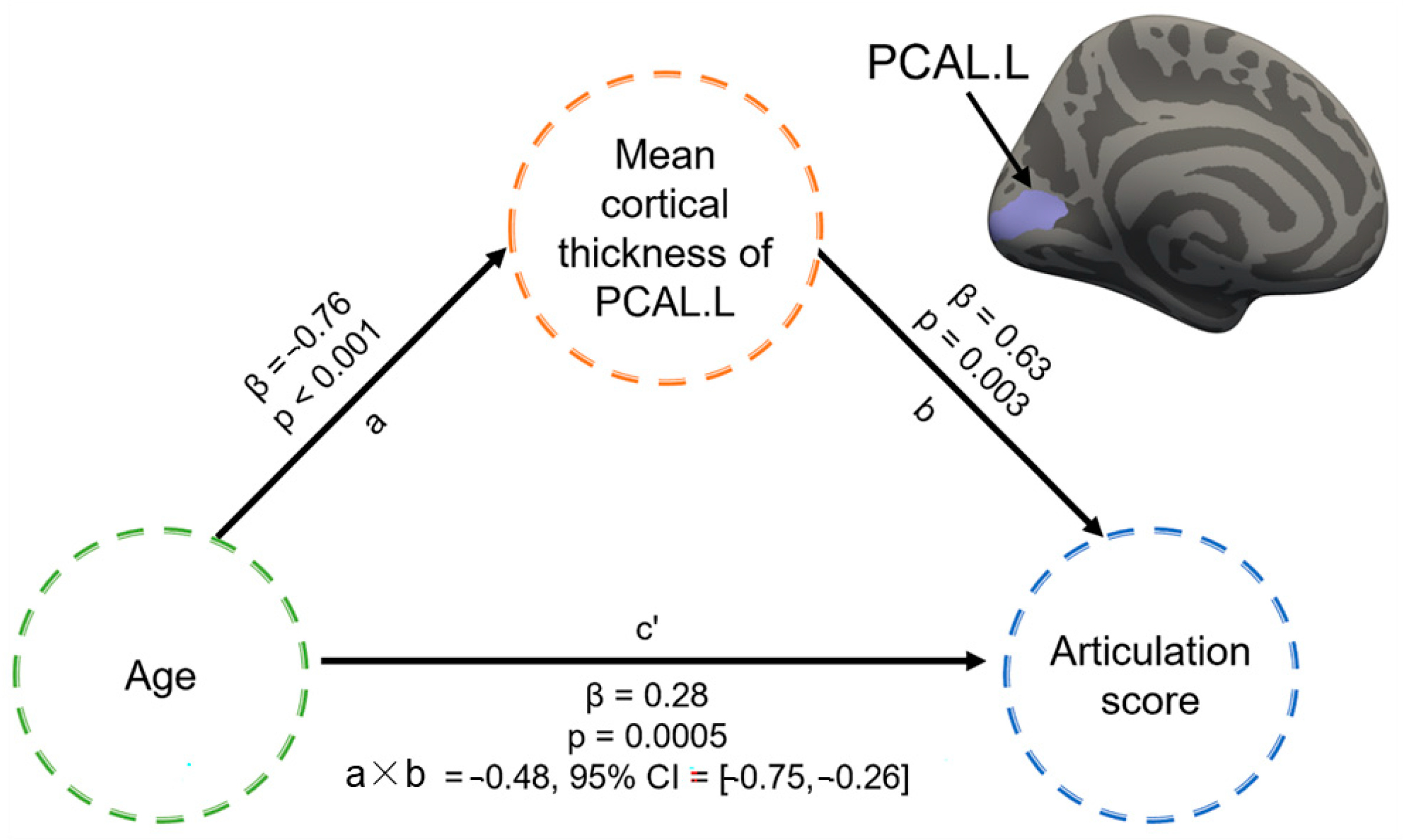
3.4. Mediation Effect of Cortical Structure between Age and Articulation Function in CLP
4. Discussion
4.1. Specific Structural Changes in CLP between Different Age Durations, Adolescence vs. Adult
4.2. Cortical Structural Changes in CLP Participants during Development
4.3. Mediation Effect of the Left Pericalcarine Cortex between Age and Speech Performance of CLP
5. Limitations
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Goh, B.S.; Tang, C.L.; Hashim, N.D.; Annamalay, T.; Abd Rahman, F.N. Hearing status and behavioural patterns among school aged children with cleft lip and/or palate. Int. J. Pediatr. Otorhinolaryngol. 2019, 118, 1–5. [Google Scholar] [CrossRef]
- Genisca, A.E.; Frias, J.L.; Broussard, C.S.; Honein, M.A.; Lammer, E.J.; Moore, C.A.; Shaw, G.M.; Murray, J.C.; Yang, W.; Rasmussen, S.A.; et al. Orofacial clefts in the National Birth Defects Prevention Study, 1997–2004. Am. J. Med. Genet. A 2009, 149A, 1149–1158. [Google Scholar] [CrossRef] [Green Version]
- Fitzsimons, D.A. International confederation for cleft lip and palate and related craniofacial anomalies task force report: Speech assessment. Cleft Palate Craniofac. J. 2014, 51, e138–e145. [Google Scholar] [CrossRef]
- Richman, L.C.; McCoy, T.E.; Conrad, A.L.; Nopoulos, P.C. Neuropsychological, behavioral, and academic sequelae of cleft: Early developmental, school age, and adolescent/young adult outcomes. Cleft Palate Craniofac. J. 2012, 49, 387–396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, S. Early Language and Speech Intervention Program for Chinese Mandarin Toddlers with Cleft Palate. Chin. Sci. J. Hear. Speech Rehabil. 2019, 17, 4. [Google Scholar]
- Lindeborg, M.M.; Shakya, P.; Rai, S.M.; Shaye, D.A. Optimizing speech outcomes for cleft palate. Curr. Opin. Otolaryngol. Head Neck Surg. 2020, 28, 206–211. [Google Scholar] [CrossRef]
- Nopoulos, P.; Berg, S.; Canady, J.; Richman, L.; Van Demark, D.; Andreasen, N.C. Abnormal brain morphology in patients with isolated cleft lip, cleft palate, or both: A preliminary analysis. Cleft Palate Craniofac. J. 2000, 37, 441–446. [Google Scholar] [CrossRef]
- Nopoulos, P.; Berg, S.; Canady, J.; Richman, L.; Van Demark, D.; Andreasen, N.C. Structural brain abnormalities in adult males with clefts of the lip and/or palate. Genet. Med. 2002, 4, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Nopoulos, P.; Berg, S.; VanDemark, D.; Richman, L.; Canady, J.; Andreasen, N.C. Increased incidence of a midline brain anomaly in patients with nonsyndromic clefts of the lip and/or palate. J. Neuroimaging 2001, 11, 418–424. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Zhang, W.; Li, C.; Wang, M.; Wang, S.; Chen, R.; Zhang, X. Articulation rehabilitation induces cortical plasticity in adults with non-syndromic cleft lip and palate. Aging 2020, 12, 13147–13159. [Google Scholar] [CrossRef]
- Shriver, A.S.; Canady, J.; Richman, L.; Andreasen, N.C.; Nopoulos, P. Structure and function of the superior temporal plane in adult males with cleft lip and palate: Pathologic enlargement with no relationship to childhood hearing deficits. J. Child Psychol. Psychiatry 2006, 47, 994–1002. [Google Scholar] [CrossRef] [PubMed]
- Weinberg, S.M.; Andreasen, N.C.; Nopoulos, P. Three-dimensional morphometric analysis of brain shape in nonsyndromic orofacial clefting. J. Anat. 2009, 214, 926–936. [Google Scholar] [CrossRef]
- Nopoulos, P.; Choe, I.; Berg, S.; Van Demark, D.; Canady, J.; Richman, L. Ventral frontal cortex morphology in adult males with isolated orofacial clefts: Relationship to abnormalities in social function. Cleft Palate Craniofac. J. 2005, 42, 138–144. [Google Scholar] [CrossRef] [PubMed]
- Xie, N.; Yang, F.; Gan, Y.; McPherson, B.; Xiang, K.; Shu, H.; Lin, F. Structural MRI of the Congnition associated Brain Regions in Children with Cleft Lip and Palate. Chin. J. CT MRI 2012, 10, 4. [Google Scholar]
- Ullman, M.T.; Earle, F.S.; Walenski, M.; Janacsek, K. The Neurocognition of Developmental Disorders of Language. Annu. Rev. Psychol. 2020, 71, 389–417. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.; Zhang, W.; Wang, M.; Wang, S.; Li, Z.; Zhao, C.; Lin, M.; Si, Q.; Li, X.; Liang, Y.; et al. Reading-related Brain Function Restored to Normal After Articulation Training in Patients with Cleft Lip and Palate: An fMRI Study. Neurosci. Bull. 2022, 38, 1215–1228. [Google Scholar] [CrossRef]
- Rao, B.; Cheng, H.; Fan, Y.; Zhang, W.; Chen, R.; Peng, Y. Topological properties of the resting-state functional network in nonsyndromic cleft lip and palate children after speech rehabilitation. J. Integr. Neurosci. 2020, 19, 285–293. [Google Scholar] [PubMed]
- Guye, M.; Bettus, G.; Bartolomei, F.; Cozzone, P.J. Graph theoretical analysis of structural and functional connectivity MRI in normal and pathological brain networks. MAGMA 2010, 23, 409–421. [Google Scholar] [CrossRef]
- Martins, M.; Reis, A.M.; Castro, S.L.; Gaser, C. Gray matter correlates of reading fluency deficits: SES matters, IQ does not. Brain Struct. Funct. 2021, 226, 2585–2601. [Google Scholar] [CrossRef]
- Weber, S.; Hausmann, M.; Kane, P.; Weis, S. The relationship between language ability and brain activity across language processes and modalities. Neuropsychologia 2020, 146, 107536. [Google Scholar] [CrossRef]
- Olulade, O.A.; Seydell-Greenwald, A.; Chambers, C.E.; Turkeltaub, P.E.; Dromerick, A.W.; Berl, M.M.; Gaillard, W.D.; Newport, E.L. The neural basis of language development: Changes in lateralization over age. Proc. Natl. Acad. Sci. USA 2020, 117, 23477–23483. [Google Scholar] [CrossRef]
- Qi, T.; Schaadt, G.; Cafiero, R.; Brauer, J.; Skeide, M.A.; Friederici, A.D. The emergence of long-range language network structural covariance and language abilities. Neuroimage 2019, 191, 36–48. [Google Scholar] [CrossRef]
- Samara, A.; Smith, K.; Brown, H.; Wonnacott, E. Acquiring variation in an artificial language: Children and adults are sensitive to socially conditioned linguistic variation. Cogn. Psychol. 2017, 94, 85–114. [Google Scholar] [CrossRef]
- Gaillard, W.D.; Sachs, B.C.; Whitnah, J.R.; Ahmad, Z.; Balsamo, L.M.; Petrella, J.R.; Braniecki, S.H.; McKinney, C.M.; Hunter, K.; Xu, B.; et al. Developmental aspects of language processing: fMRI of verbal fluency in children and adults. Hum. Brain Mapp. 2003, 18, 176–185. [Google Scholar] [CrossRef]
- Zhang, W.J.; Zhao, C.; Li, C.L.; Yang, X.T.; Chen, R.J. Gray matter morphological changes in children with cleft palate speech disorder. Zhonghua Kou Qiang Yi Xue Za Zhi 2022, 57, 899–906. [Google Scholar] [PubMed]
- Nandita, V.; Nicholas, B.A.; George, Y.; Meg, D.; Murat, Y.; Julian, G.S.; Sarah, W. Brain development during adolescence: A mixed-longitudinal investigation of cortical thickness, surface area, and volume. Hum. Brain Mapp. 2016, 37, 2027–2038. [Google Scholar]
- Sunhae, S.; Berna, G.; Eveline, A.C.; Luke, J.C. Medial prefrontal cortical thinning mediates shifts in other-regarding preferences during adolescence. Sci. Rep. 2017, 7, 8510. [Google Scholar]
- Goto, M.; Abe, O.; Aoki, S.; Hayashi, N.; Miyati, T.; Takao, H.; Iwatsubo, T.; Yamashita, F.; Matsuda, H.; Mori, H.; et al. Diffeomorphic Anatomical Registration Through Exponentiated Lie Algebra provides reduced effect of scanner for cortex volumetry with atlas-based method in healthy subjects. Neuroradiology 2013, 55, 869–875. [Google Scholar] [CrossRef] [PubMed]
- Nieto-Castanon, A. Brain-wide connectome inferences using functional connectivity MultiVariate Pattern Analyses (fc-MVPA). PLoS Comput. Biol. 2022, 18, e1010634. [Google Scholar] [CrossRef]
- Wang, Y.; Guan, H.; Ma, L.; Luo, J.; Chu, C.; Hu, M.; Zhao, G.; Men, W.; Tan, S.; Gao, J.H.; et al. Learning to read may help promote attention by increasing the volume of the left middle frontal gyrus and enhancing its connectivity to the ventral attention network. Cereb. Cortex 2023, 33, 2260–2272. [Google Scholar] [CrossRef]
- Kim, H.; Wang, K.; Cutting, L.E.; Willcutt, E.G.; Petrill, S.A.; Leopold, D.R.; Reineberg, A.E.; Thompson, L.A.; Banich, M.T. The Angular Gyrus as a Hub for Modulation of Language-related Cortex by Distinct Prefrontal Executive Control Regions. J. Cogn. Neurosci. 2022, 34, 2275–2296. [Google Scholar] [CrossRef]
- Zhang, W.; Xiang, M.; Wang, S. The role of left angular gyrus in the representation of linguistic composition relations. Hum. Brain Mapp. 2022, 43, 2204–2217. [Google Scholar] [CrossRef]
- Alyahya, R.S.W.; Halai, A.D.; Conroy, P.; Lambon Ralph, M.A. A unified model of post-stroke language deficits including discourse production and their neural correlates. Brain 2020, 143, 1541–1554. [Google Scholar] [CrossRef] [PubMed]
- Dong, J.W.; Brennan, N.M.; Izzo, G.; Peck, K.K.; Holodny, A.I. fMRI activation in the middle frontal gyrus as an indicator of hemispheric dominance for language in brain tumor patients: A comparison with Broca’s area. Neuroradiology 2016, 58, 513–520. [Google Scholar] [CrossRef] [Green Version]
- Brown, S.; Martinez, M.J.; Parsons, L.M. Music and language side by side in the brain: A PET study of the generation of melodies and sentences. Eur. J. Neurosci. 2006, 23, 2791–2803. [Google Scholar] [CrossRef]
- Wang, S.; Zhu, Z.; Zhang, J.X.; Wang, Z.; Xiao, Z.; Xiang, H.; Chen, H.C. Broca’s area plays a role in syntactic processing during Chinese reading comprehension. Neuropsychologia 2008, 46, 1371–1378. [Google Scholar] [CrossRef]
- Abrahams, S.; Goldstein, L.H.; Simmons, A.; Brammer, M.J.; Williams, S.C.; Giampietro, V.P.; Andrew, C.M.; Leigh, P.N. Functional magnetic resonance imaging of verbal fluency and confrontation naming using compressed image acquisition to permit overt responses. Hum. Brain Mapp. 2003, 20, 29–40. [Google Scholar] [CrossRef]
- Leung, H.C.; Gore, J.C.; Goldman-Rakic, P.S. Sustained mnemonic response in the human middle frontal gyrus during on-line storage of spatial memoranda. J. Cogn. Neurosci. 2002, 14, 659–671. [Google Scholar] [CrossRef]
- Kristinsson, S.; Thors, H.; Yourganov, G.; Magnusdottir, S.; Hjaltason, H.; Stark, B.C.; Basilakos, A.; den Ouden, D.B.; Bonilha, L.; Rorden, C.; et al. Brain Damage Associated with Impaired Sentence Processing in Acute Aphasia. J. Cogn. Neurosci. 2020, 32, 256–271. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Deng, J.; Zhang, Z.; Zhang, Z.Y.; Sun, Y.G.; Yang, T.; Yao, H. Orbitofrontal control of visual cortex gain promotes visual associative learning. Nat. Commun. 2020, 11, 2784. [Google Scholar] [CrossRef] [PubMed]
- Lancaster, H.S.; Lien, K.M.; Chow, J.C.; Frey, J.R.; Scherer, N.J.; Kaiser, A.P. Early Speech and Language Development in Children With Nonsyndromic Cleft Lip and/or Palate: A Meta-Analysis. J. Speech Lang. Hear. Res. 2020, 63, 14–31. [Google Scholar] [CrossRef] [Green Version]
- Grabski, K.; Sato, M. Adaptive phonemic coding in the listening and speaking brain. Neuropsychologia 2020, 136, 107267. [Google Scholar] [CrossRef] [PubMed]
- Silva, A.B.; Liu, J.R.; Zhao, L.; Levy, D.F.; Scott, T.L.; Chang, E.F. A Neurosurgical Functional Dissection of the Middle Precentral Gyrus during Speech Production. J. Neurosci. 2022, 42, 8416–8426. [Google Scholar] [CrossRef]
- Japee, S.; Holiday, K.; Satyshur, M.D.; Mukai, I.; Ungerleider, L.G. A role of right middle frontal gyrus in reorienting of attention: A case study. Front. Syst. Neurosci. 2015, 9, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adamson, C.L.; Anderson, V.A.; Nopoulos, P.; Seal, M.L.; Da Costa, A.C. Regional brain morphometric characteristics of nonsyndromic cleft lip and palate. Dev. Neurosci. 2014, 36, 490–498. [Google Scholar] [CrossRef]
- Bodoni, P.S.B.; Leoni, R.F.; do Vale, A.B.; da Silva, P.H.R.; Meira Junior, S.G.; Richieri Costa, A.; Tabaquim, M.L.M. Neuropsychological functioning and its relationship with brain anatomical measures of children and adolescents with non-syndromic cleft lip and palate. Child Neuropsychol. 2021, 27, 2–16. [Google Scholar] [CrossRef] [PubMed]
- Noven, M.; Schremm, A.; Horne, M.; Roll, M. Cortical thickness and surface area of left anterior temporal areas affects processing of phonological cues to morphosyntax. Brain Res. 2021, 1750, 147150. [Google Scholar] [CrossRef]
- Jiao, Y.; Lin, F.; Wu, J.; Li, H.; Fu, W.; Huo, R.; Cao, Y.; Wang, S.; Zhao, J. Plasticity in language cortex and white matter tracts after resection of dominant inferior parietal lobule arteriovenous malformations: A combined fMRI and DTI study. J. Neurosurg. 2020, 134, 953–960. [Google Scholar] [CrossRef]
- Ogura, A.; Watanabe, H.; Kawabata, K.; Ohdake, R.; Tanaka, Y.; Masuda, M.; Kato, T.; Imai, K.; Yokoi, T.; Hara, K.; et al. Semantic deficits in ALS related to right lingual/fusiform gyrus network involvement. EBioMedicine 2019, 47, 506–517. [Google Scholar] [CrossRef] [Green Version]
- Jung, J.; Kang, J.; Won, E.; Nam, K.; Lee, M.S.; Tae, W.S.; Ham, B.J. Impact of lingual gyrus volume on antidepressant response and neurocognitive functions in Major Depressive Disorder: A voxel-based morphometry study. J. Affect. Disord. 2014, 169, 179–187. [Google Scholar] [CrossRef]
- Deng, Y.; Chou, T.L.; Ding, G.S.; Peng, D.L.; Booth, J.R. The involvement of occipital and inferior frontal cortex in the phonological learning of Chinese characters. J. Cogn. Neurosci. 2011, 23, 1998–2012. [Google Scholar] [CrossRef] [Green Version]
- Kaestner, E.; Thesen, T.; Devinsky, O.; Doyle, W.; Carlson, C.; Halgren, E. An Intracranial Electrophysiology Study of Visual Language Encoding: The Contribution of the Precentral Gyrus to Silent Reading. J. Cogn. Neurosci. 2021, 33, 2197–2214. [Google Scholar] [CrossRef] [PubMed]
- Basilakos, A.; Smith, K.G.; Fillmore, P.; Fridriksson, J.; Fedorenko, E. Functional Characterization of the Human Speech Articulation Network. Cereb. Cortex 2018, 28, 1816–1830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hagoort, P. Nodes and networks in the neural architecture for language: Broca’s region and beyond. Curr. Opin. Neurobiol. 2014, 28, 136–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krishnan, S.; Leech, R.; Mercure, E.; Lloyd-Fox, S.; Dick, F. Convergent and Divergent fMRI Responses in Children and Adults to Increasing Language Production Demands. Cereb. Cortex 2015, 25, 3261–3277. [Google Scholar] [CrossRef] [Green Version]
- Hage, S.R.; Nieder, A. Dual Neural Network Model for the Evolution of Speech and Language. Trends Neurosci. 2016, 39, 813–829. [Google Scholar] [CrossRef]
- Bedny, M.; Pascual-Leone, A.; Dravida, S.; Saxe, R. A sensitive period for language in the visual cortex: Distinct patterns of plasticity in congenitally versus late blind adults. Brain Lang. 2012, 122, 162–170. [Google Scholar] [CrossRef]
- Watkins, K.E.; Coullon, G.S.L.; Bridge, H. Language and nonverbal auditory processing in the occipital cortex of individuals who are congenitally blind due to anophthalmia. Neuropsychologia 2022, 173, 108304. [Google Scholar] [CrossRef]
- Lopes, R.; Nunes, R.G.; Simoes, M.R.; Secca, M.F.; Leal, A. The Visual Word Form Area remains in the dominant hemisphere for language in late-onset left occipital lobe epilepsies: A postsurgery analysis of two cases. Epilepsy Behav. 2015, 46, 91–98. [Google Scholar] [CrossRef]
- Vonk, J.M.J.; Rizvi, B.; Lao, P.J.; Budge, M.; Manly, J.J.; Mayeux, R.; Brickman, A.M. Letter and Category Fluency Performance Correlates with Distinct Patterns of Cortical Thickness in Older Adults. Cereb. Cortex 2019, 29, 2694–2700. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CLP | HC | T/X2 | p | |
|---|---|---|---|---|
| (n = 49) | (n = 59) | |||
| Adolescents (n = 63) | CLP_Ado = 26 | HC_Ado = 37 | ||
| Age, years a | 8.88 (2.39) [5–14] | 9.89 (2.72) [5–16] | −1.52 | 0.133 |
| Gender (Female/Male)b | 8/18 (44.4%) | 10/27 (55.6%) | 0.11 | 0.746 |
| Articulation test score | 44.83 (25.73) [0–93] | -- | -- | -- |
| Adults (n = 45) | CLP_Adu = 23 | HC_Adu = 22 | ||
| Age, years (range) a | 23.35 (4.16) [18–35] | 24.41 (4.38) [19–37] | −0.83 | 0.409 |
| Gender (Female/Male) b | 6/17 (30.0%) | 14/8 (70%) | 6.42 | 0.011 |
| Articulation test score | 51.00 (23.57) [13–93] | -- | -- | -- |
| Total (n = 108) | CLP = 49 | HC = 59 | ||
| Age, years (range) a | 15.67 (8.01) [5–35] | 15.31 (7.85) [5–37] | 0.24 | 0.810 |
| Gender (Female/Male) b | 14/35 (36.8%) | 24/35 (63.2) | 1.72 | 0.190 |
| Articulation test score | 46.82 (23.92) [0–93] |
| No. Cluster | Cluster Size | Peak MNI Coordinates | Peak t Value | Anatomic Locations Based on AAL Atlas | |||
|---|---|---|---|---|---|---|---|
| x | y | z | |||||
| Significant group differences a | |||||||
| Ado: CLP < HC | 1 | 107 | 31.25 | 48.75 | 1.76 | −3.31 | Left angular gyrus |
| 2 | 237 | 31.25 | 48.75 | 1.77 | −3.73 | Right angular gyrus | |
| 3 | 283 | 31.25 | 48.75 | 1.78 | −4.01 | Left middle occipital gyrus | |
| 4 | 136 | 31.25 | 48.75 | 1.79 | −3.69 | Right angular gyrus | |
| 5 | 228 | 31.25 | 48.75 | 1.8 | −3.92 | Right superior parietal gyrus | |
| 6 | 175 | 31.25 | 48.75 | 1.81 | −4.12 | Right postcentral gyrus | |
| Adu: CLP > HC | 1 | 125 | 31.25 | 48.75 | 1.82 | 3.53 | Left superior temporal gyrus |
| 2 | 113 | 31.25 | 48.75 | 1.83 | 3.94 | Right middle frontal gyrus | |
| 3 | 121 | 31.25 | 48.75 | 1.84 | 4.14 | Left precentral gyrus | |
| Interaction: Group × Age b | |||||||
| CLP > HC | 1 | 112 | −21.75 | −61.25 | −6.25 | 4.02 | Left lingual gyrus |
| 2 | 260 | 36.25 | −42.25 | 72.75 | 4.19 | Right precentral gyrus | |
| No. Cluster | Cluster Size (mm2) | Peak MNI305 Coordinates | Peak z Value | Anatomic Locations Based on AAL Atlas | |||
|---|---|---|---|---|---|---|---|
| x | y | z | |||||
| Significant group differences a | |||||||
| Ado:
CLP < HC (cortical thickness) | 1 | 644.5 | −54.2 | −57.6 | −12.2 | 3.68 | Left inferior temporal cortex |
| Ado: CLP > HC (surface area) | 1 | 845.36 | −50.5 | −59.4 | 28.9 | −3.89 | Left inferior parietal cortex |
| Interaction: Group × Age (cortical thickness) b | |||||||
| CLP < HC | 1 | 374.58 | −55.4 | −52.2 | −14.5 | −4.36 | Left inferior temporal cortex |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, W.; Zhao, C.; Sun, L.; Yang, X.; Yang, L.; Liang, Y.; Zhang, X.; Du, X.; Chen, R.; Li, C. Articulation-Function-Associated Cortical Developmental Changes in Patients with Cleft Lip and Palate. Brain Sci. 2023, 13, 550. https://doi.org/10.3390/brainsci13040550
Zhang W, Zhao C, Sun L, Yang X, Yang L, Liang Y, Zhang X, Du X, Chen R, Li C. Articulation-Function-Associated Cortical Developmental Changes in Patients with Cleft Lip and Palate. Brain Sciences. 2023; 13(4):550. https://doi.org/10.3390/brainsci13040550
Chicago/Turabian StyleZhang, Wenjing, Cui Zhao, Liwei Sun, Xintao Yang, Linrui Yang, Ying Liang, Xu Zhang, Xiaoxia Du, Renji Chen, and Chunlin Li. 2023. "Articulation-Function-Associated Cortical Developmental Changes in Patients with Cleft Lip and Palate" Brain Sciences 13, no. 4: 550. https://doi.org/10.3390/brainsci13040550