
Identification and Characterization of TMEM119-Positive Cells in the Postnatal and Adult Murine Cochlea


Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Tissue Harvesting
2.2. The Noise Trauma Model
2.3. Tissue Processing for Cryosections
2.4. Immunolabeling and Confocal Microscopy
2.5. Protein Isolation and Western Blot Analysis
2.6. RNA Isolation and Semiquantitative Real-Time Reverse-Transcription–Polymerase Chain Reaction (RT-PCR)
2.7. Statistical Analysis
3. Results
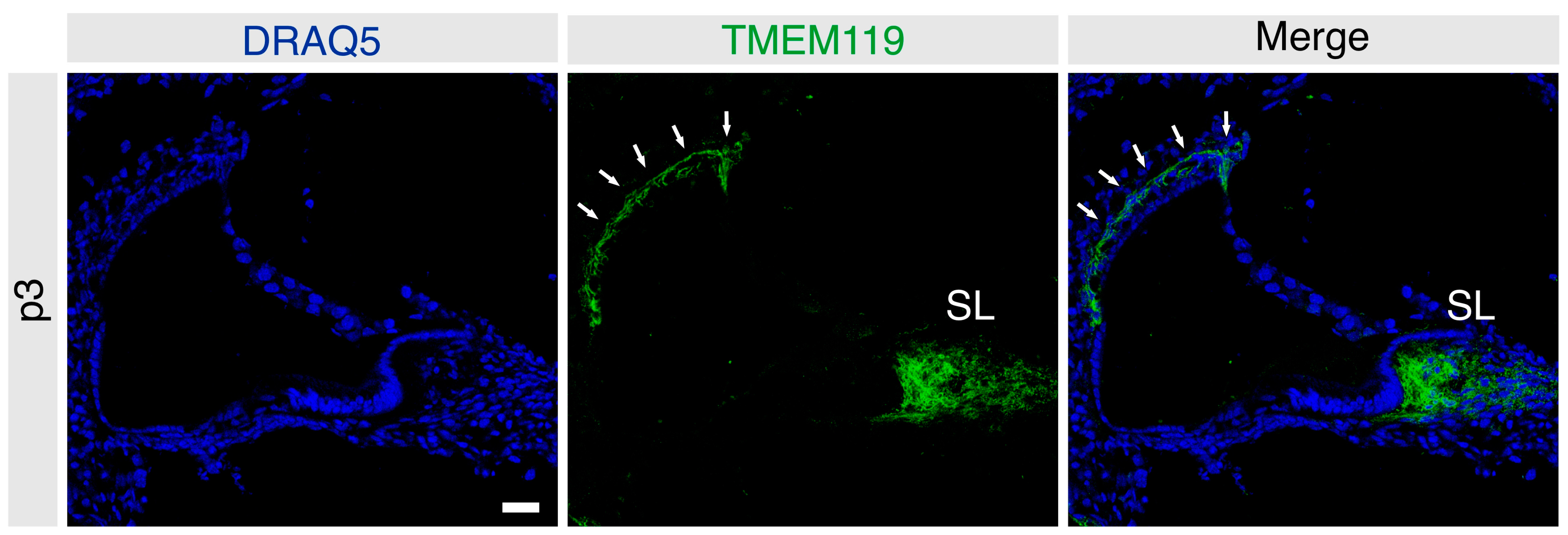
3.1. Localization of TMEM119 Protein in the Immature Postnatal Murine Cochlea
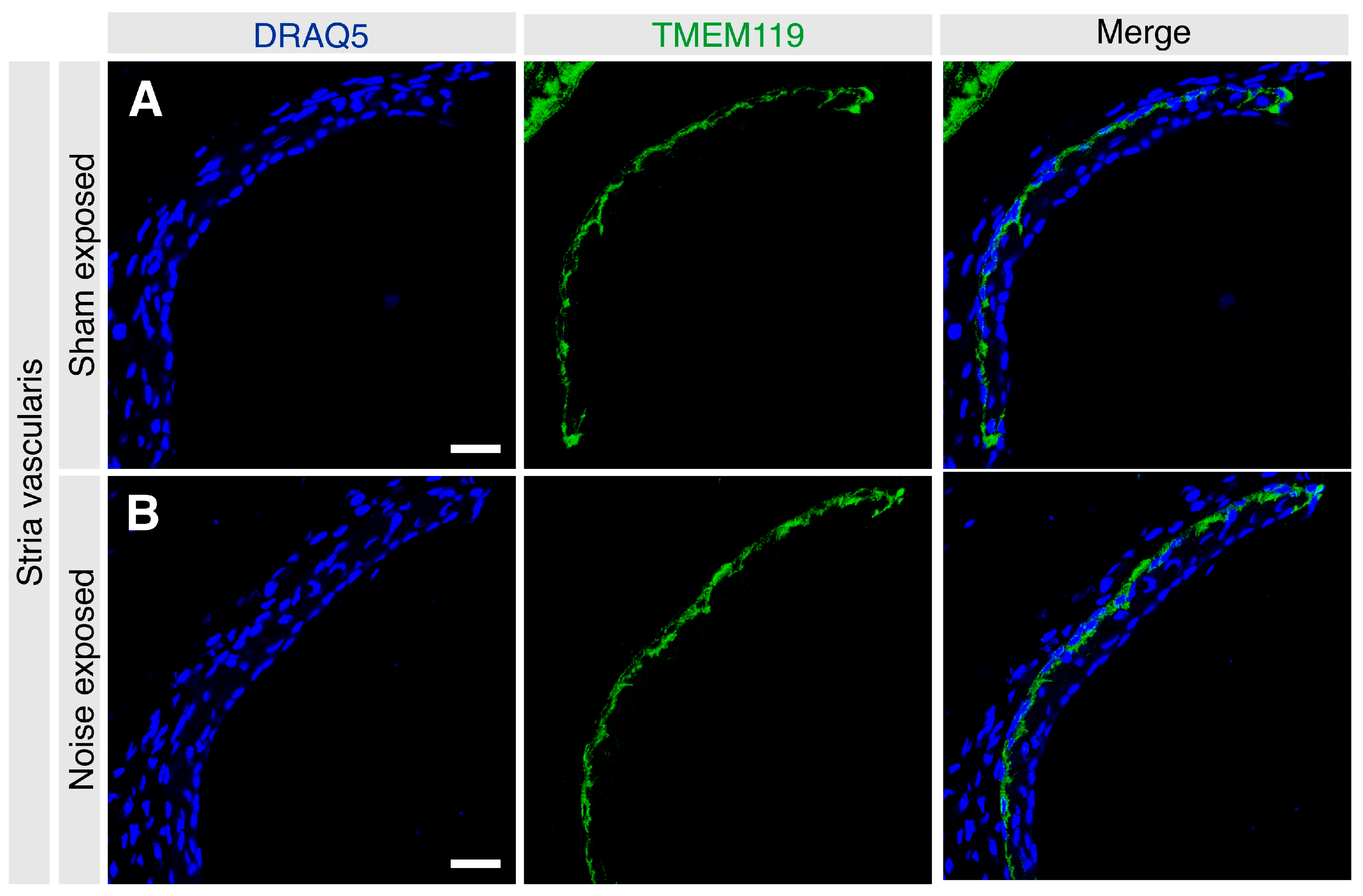
3.2. Localization of TMEM119 Protein in the Adult Murine Cochlea under Basal and Noise-Exposed Conditions
3.3. Quantitative Comparison of the TMEM119 Protein Levels in the Postnatal Cochleae and the Brain
3.4. TMEM119 Gene Expression in the Cochlear Tissues
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Perry, V.H.; Hume, D.A.; Gordon, S. Immunohistochemical localization of macrophages and microglia in the adult and developing mouse brain. Neuroscience 1985, 15, 313–326. [Google Scholar] [CrossRef]
- De Groot, C.J.; Huppes, W.; Sminia, T.; Kraal, G.; Dijkstra, C.D. Determination of the origin and nature of brain macrophages and microglial cells in mouse central nervous system, using non-radioactive in situ hybridization and immunoperoxidase techniques. Glia 1992, 6, 301–309. [Google Scholar] [CrossRef]
- Conde, J.R.; Streit, W.J. Microglia in the aging brain. J. Neuropathol. Exp. Neurol. 2006, 65, 199–203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tremblay, M.E.; Stevens, B.; Sierra, A.; Wake, H.; Bessis, A.; Nimmerjahn, A. The role of microglia in the healthy brain. J. Neurosci. 2011, 31, 16064–16069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ekdahl, C.T.; Kokaia, Z.; Lindvall, O. Brain inflammation and adult neurogenesis: The dual role of microglia. Neuroscience 2009, 158, 1021–1029. [Google Scholar] [CrossRef]
- Salter, M.W.; Beggs, S. Sublime microglia: Expanding roles for the guardians of the CNS. Cell 2014, 158, 15–24. [Google Scholar] [CrossRef] [Green Version]
- Lenz, K.M.; Nelson, L.H. Microglia and beyond: Innate immune cells as regulators of brain development and behavioral function. Front. Immunol. 2018, 9, 698. [Google Scholar] [CrossRef] [Green Version]
- Wieghofer, P.; Knobeloch, K.P.; Prinz, M. Genetic targeting of microglia. Glia 2015, 63, 1–22. [Google Scholar] [CrossRef]
- Bennett, M.L.; Bennett, F.C.; Liddelow, S.A.; Ajami, B.; Zamanian, J.L.; Fernhoff, N.B.; Mulinyawe, S.B.; Bohlen, C.J.; Adil, A.; Tucker, A.; et al. New tools for studying microglia in the mouse and human CNS. Proc. Natl. Acad. Sci. USA 2016, 113, E1738–E1746. [Google Scholar] [CrossRef] [Green Version]
- Buttgereit, A.; Lelios, I.; Yu, X.; Vrohlings, M.; Krakoski, N.R.; Gautier, E.L.; Nishinakamura, R.; Becher, B.; Greter, M. Sall1 is a transcriptional regulator defining microglia identity and function. Nat. Immunol. 2016, 17, 1397–1406. [Google Scholar] [CrossRef]
- Neidert, N.; Von Ehr, A.; Zöller, T.; Spittau, B. Microglia-specific expression of Olfml3 is directly regulated by transforming growth factor β1-induced Smad2 signaling. Front. Immunol. 2018, 9, 1728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Werner, Y.; Mass, E.; Kumar, P.A.; Ulas, T.; Händler, K.; Horne, A.; Klee, K.; Lupp, A.; Schütz, D.; Saaber, F.; et al. Cxcr4 distinguishes HSC-derived monocytes from microglia and reveals monocyte immune responses to experimental stroke. Nat. Neurosci. 2020, 23, 351–362. [Google Scholar] [CrossRef] [PubMed]
- McKinsey, G.L.; Lizama, C.O.; Keown-Lang, A.E.; Niu, A.; Santander, N.; Larpthaveesarp, A.; Chee, E.; Gonzalez, F.F.; Arnold, T.D. A new genetic strategy for targeting microglia in development and disease. Elife 2020, 9, e54590. [Google Scholar] [CrossRef]
- Kaiser, T.; Feng, G. Tmem119-EGFP and Tmem119-CreERT2 transgenic mice for labeling and manipulating microglia. eNeuro 2019, 6, ENEURO.0448-0418.2019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masuda, T.; Amann, L.; Sankowski, R.; Staszewski, O.; Lenz, M.; Errico, P.D.; Snaidero, N.; Jordão, M.J.C.; Böttcher, C.; Kierdorf, K.; et al. Novel Hexb-based tools for studying microglia in the CNS. Nat. Immunol. 2020, 21, 802–815. [Google Scholar] [CrossRef]
- Brandenburg, S.; Blank, A.; Bungert, A.D.; Vajkoczy, P. Distinction of microglia and macrophages in glioblastoma: Close relatives, different tasks? Int. J. Mol. Sci. 2020, 22, 194. [Google Scholar] [CrossRef]
- Wojtera, M.; Sobow, T.; Kloszewska, I.; Liberski, P.P.; Brown, D.R.; Sikorska, B. Expression of immunohistochemical markers on microglia in Creutzfeldt-Jakob disease and Alzheimer’s disease: Morphometric study and review of the literature. Folia Neuropathol. 2012, 50, 74–84. [Google Scholar]
- Korzhevskii, D.E.; Kirik, O.V. Brain microglia and microglial markers. Neurosci. Behav. Physiol. 2016, 46, 284–290. [Google Scholar] [CrossRef]
- Hopperton, K.E.; Mohammad, D.; Trepanier, M.O.; Giuliano, V.; Bazinet, R.P. Markers of microglia in post-mortem brain samples from patients with Alzheimer’s disease: A systematic review. Mol. Psychiatry 2018, 23, 177–198. [Google Scholar] [CrossRef]
- Jurga, A.M.; Paleczna, M.; Kuter, K.Z. Overview of general and discriminating markers of differential microglia phenotypes. Front. Cell. Neurosci. 2020, 14, 198. [Google Scholar] [CrossRef]
- Furman, N.T.; Gottlieb, A.; Prabhakara, K.S.; Bedi, S.; Caplan, H.W.; Ruppert, K.A.; Srivastava, A.K.; Olson, S.D.; Cox, C.S. High-resolution and differential analysis of rat microglial markers in traumatic brain injury: Conventional flow cytometric and bioinformatics analysis. Sci. Rep. 2020, 10, 11991. [Google Scholar] [CrossRef]
- Ito, D.; Imai, Y.; Ohsawa, K.; Nakajima, K.; Fukuuchi, Y.; Kohsaka, S. Microglia-specific localisation of a novel calcium binding protein, Iba1. Brain Res. Mol. Brain Res. 1998, 57, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Hirasawa, T.; Ohsawa, K.; Imai, Y.; Ondo, Y.; Akazawa, C.; Uchino, S.; Kohsaka, S. Visualization of microglia in living tissues using Iba1-EGFP transgenic mice. J. Neurosci. Res. 2005, 81, 357–362. [Google Scholar] [CrossRef] [PubMed]
- Burgess, M.; Wicks, K.; Gardasevic, M.; Mace, K.A. Cx3CR1 expression identifies distinct macrophage populations that contribute differentially to inflammation and repair. Immunohorizons 2019, 3, 262–273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, B.A.; Beamer, M.; Ahmed, S. Fractalkine/CX3CL1: A potential new target for inflammatory diseases. Mol. Interv. 2010, 10, 263–270. [Google Scholar] [CrossRef] [Green Version]
- Satoh, J.; Kino, Y.; Asahina, N.; Takitani, M.; Miyoshi, J.; Ishida, T.; Saito, Y. TMEM119 marks a subset of microglia in the human brain. Neuropathology 2016, 36, 39–49. [Google Scholar] [CrossRef]
- Haynes, S.E.; Hollopeter, G.; Yang, G.; Kurpius, D.; Dailey, M.E.; Gan, W.B.; Julius, D. The P2Y12 receptor regulates microglial activation by extracellular nucleotides. Nat. Neurosci. 2006, 9, 1512–1519. [Google Scholar] [CrossRef]
- Walker, D.G.; Tang, T.M.; Mendsaikhan, A.; Tooyama, I.; Serrano, G.E.; Sue, L.I.; Beach, T.G.; Lue, L.F. Patterns of expression of purinergic receptor P2RY12, a putative marker for non-activated microglia, in aged and Alzheimer’s disease brains. Int. J. Mol. Sci. 2020, 21, 678. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.; Molnar, M.; Garnham, C.; Benav, H.; Rask-Andersen, H. Macrophages in the human cochlea: Saviors or predators-a study using super-resolution immunohistochemistry. Front. Immunol. 2018, 9, 223. [Google Scholar] [CrossRef] [Green Version]
- Kishimoto, I.; Okano, T.; Nishimura, K.; Motohashi, T.; Omori, K. Early development of resident macrophages in the mouse cochlea depends on yolk sac hematopoiesis. Front. Neurol. 2019, 10, 1115. [Google Scholar] [CrossRef]
- O’Malley, J.T.; Nadol, J.B.; McKenna, M.J. Anti CD163+, Iba1+, and CD68+ cells in the adult human inner ear: Normal distribution of an unappreciated class of macrophages/microglia and implications for inflammatory otopathology in humans. Otol. Neurotol. 2016, 37, 99–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okano, T.; Nakagawa, T.; Kita, T.; Kada, S.; Yoshimoto, M.; Nakahata, T.; Ito, J. Bone marrow-derived cells expressing Iba1 are constitutively present as resident tissue macrophages in the mouse cochlea. J. Neurosci. Res. 2008, 86, 1758–1767. [Google Scholar] [CrossRef]
- Okayasu, T.; Quesnel, A.M.; O’Malley, J.T.; Kamakura, T.; Nadol, J.B. The distribution and prevalence of macrophages in the cochlea following cochlear implantation in the human: An immunohistochemical study using anti-Iba1 antibody. Otol. Neurotol. 2019, 41, e304–e316. [Google Scholar] [CrossRef]
- Kaur, T.; Zamani, D.; Tong, L.; Rubel, E.W.; Ohlemiller, K.K.; Hirose, K.; Warchol, M.E. Fractalkine signaling regulates macrophage recruitment into the cochlea and promotes the survival of spiral ganglion neurons after selective hair cell lesion. J. Neurosci. 2015, 35, 15050–15061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, S.; Yu, H.; Yu, H.; Honglin, M.; Ni, W.; Zhang, Y.; Guo, L.; He, Y.; Xue, Z.; Ni, Y.; et al. Inhibition of the activation and recruitment of microglia-like cells protects against neomycin-induced ototoxicity. Mol. Neurobiol. 2015, 51, 252–267. [Google Scholar] [CrossRef] [PubMed]
- Frye, M.D.; Yang, W.; Zhang, C.; Xiong, B.; Hu, B.H. Dynamic activation of basilar membrane macrophages in response to chronic sensory cell degeneration in aging mouse cochleae. Hear. Res. 2017, 344, 125–134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, L.N.; Xing, Y.; Noble, K.V.; Barth, J.L.; Panganiban, C.H.; Smythe, N.M.; Bridges, M.C.; Zhu, J.; Lang, H. Macrophage-mediated glial cell elimination in the postnatal mouse cochlea. Front. Mol. Neurosci. 2017, 10, 407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, Y.; Zhang, C.; Frye, M.; Yang, W.; Ding, D.; Sharma, A.; Guo, W.; Hu, B.H. Differential fates of tissue macrophages in the cochlea during postnatal development. Hear. Res. 2018, 365, 110–126. [Google Scholar] [CrossRef]
- He, W.; Yu, J.; Sun, Y.; Kong, W. Macrophages in noise-exposed cochlea: Changes, regulation and the potential role. Aging Dis. 2020, 11, 191–199. [Google Scholar] [CrossRef] [Green Version]
- Bae, S.H.; Kwak, S.H.; Yoo, J.E.; Kim, K.M.; Hyun, Y.M.; Choi, J.Y.; Jung, J. 3D distribution of cochlear macrophages in the lateral wall of cleared cochlea. Clin. Exp. Otorhinolaryngol. 2020, 14, 179–184. [Google Scholar] [CrossRef]
- Hirose, K.; Discolo, C.M.; Keasler, J.R.; Ransohoff, R. Mononuclear phagocytes migrate into the murine cochlea after acoustic trauma. J. Comp. Neurol. 2005, 489, 180–194. [Google Scholar] [CrossRef]
- Yang, W.; Vethanayagam, R.R.; Dong, Y.; Cai, Q.; Hu, B.H. Activation of the antigen presentation function of mononuclear phagocyte populations associated with the basilar membrane of the cochlea after acoustic overstimulation. Neuroscience 2015, 303, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Frye, M.D.; Zhang, C.; Hu, B.H. Lower level noise exposure that produces only TTS modulates the immune homeostasis of cochlear macrophages. J. Neuroimmunol. 2018, 323, 152–166. [Google Scholar] [CrossRef]
- Frye, M.D.; Ryan, A.F.; Kurabi, A. Inflammation associated with noise-induced hearing loss. J. Acoust. Soc. Am. 2019, 146, 4020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujioka, M.; Kanzaki, S.; Okano, H.J.; Masuda, M.; Ogawa, K.; Okano, H. Proinflammatory cytokines expression in noise-induced damaged cochlea. J. Neurosci. Res. 2006, 83, 575–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, W.; Nordström, C.K.; Danckwardt-Lillieström, N.; Rask-Andersen, H. Human inner ear immune activity: A super-resolution immunohistochemistry study. Front. Neurol. 2019, 10, 728. [Google Scholar] [CrossRef] [Green Version]
- Chen, P.; Chai, Y.; Liu, H.; Li, G.; Wang, L.; Yang, T.; Wu, H. Postnatal development of microglia-like cells in mouse cochlea. Neural Plast. 2018, 2018, 1970150. [Google Scholar] [CrossRef] [PubMed]
- Möhrle, D.; Reimann, K.; Wolter, S.; Wolters, M.; Varakina, K.; Mergia, E.; Eichert, N.; Geisler, H.S.; Sandner, P.; Ruth, P.; et al. NO-sensitive guanylate cyclase isoforms NO-GC1 and NO-GC2 contribute to noise-induced inner hair cell synaptopathy. Mol. Pharmacol. 2017, 92, 375–388. [Google Scholar] [CrossRef] [Green Version]
- Bassiouni, M.; Stölzel, K.; Smorodchenko, A.; Olze, H.; Szczepek, A.J. Tackling the mouse-on-mouse problem in cochlear immunofluorescence: A simple double-blocking protocol for immunofluorescent labeling of murine cochlear sections with primary mouse antibodies. Curr. Protoc. Mouse Biol. 2020, 10, e84. [Google Scholar] [CrossRef]
- Mazurek, B.; Lou, X.; Olze, H.; Haupt, H.; Szczepek, A.J. In vitro protection of auditory hair cells by salicylate from the gentamicin-induced but not neomycin-induced cell loss. Neurosci. Lett. 2012, 506, 107–110. [Google Scholar] [CrossRef]
- Szczepek, A.J.; Dudnik, T.; Karayay, B.; Sergeeva, V.; Olze, H.; Smorodchenko, A. Mast cells in the auditory periphery of rodents. Brain Sci. 2020, 10, 697. [Google Scholar] [CrossRef] [PubMed]
- Noble, K.V.; Liu, T.; Matthews, L.J.; Schulte, B.A.; Lang, H. Age-related changes in immune cells of the human cochlea. Front. Neurol. 2019, 10, 895. [Google Scholar] [CrossRef] [Green Version]
- Wood, M.B.; Zuo, J. The contribution of immune infiltrates to ototoxicity and cochlear hair cell loss. Front. Cell. Neurosci. 2017, 11, 106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhave, S.A.; Oesterle, E.C.; Coltrera, M.D. Macrophage and microglia-like cells in the avian inner ear. J. Comp. Neurol. 1998, 398, 241–256. [Google Scholar] [CrossRef]
- Wang, Z.; Li, H. Microglia-like cells in rat organ of Corti following aminoglycoside ototoxicity. Neuroreport 2000, 11, 1389–1393. [Google Scholar] [CrossRef]
- Wang, Y.; Wei, W. Microglia-like or microglia: Results of the weak silver carbonate staining method of del Rio-Hortega. Biotech. Histochem. 2012, 87, 346–349. [Google Scholar] [CrossRef]
- Seigel, G.M.; Manohar, S.; Bai, Y.Y.; Ding, D.; Salvi, R. An immortalized microglial cell line (Mocha) derived from rat cochlea. Mol. Cell. Neurosci. 2017, 85, 202–210. [Google Scholar] [CrossRef]
- Janz, P.; Illing, R.B. A role for microglial cells in reshaping neuronal circuitry of the adult rat auditory brainstem after its sensory deafferentation. J. Neurosci. Res. 2014, 92, 432–445. [Google Scholar] [CrossRef] [PubMed]
- Rosskothen-Kuhl, N.; Hildebrandt, H.; Birkenhäger, R.; Illing, R.B. Astrocyte hypertrophy and microglia activation in the rat auditory midbrain is induced by electrical intracochlear stimulation. Front. Cell. Neurosci. 2018, 12, 43. [Google Scholar] [CrossRef] [Green Version]
- Fuentes-Santamaría, V.; Alvarado, J.C.; Juiz, J.M. Long-term interaction between microglial cells and cochlear nucleus neurons after bilateral cochlear ablation. J. Comp. Neurol. 2012, 520, 2974–2990. [Google Scholar] [CrossRef]
- Fuentes-Santamaría, V.; Alvarado, J.C.; Gabaldón-Ull, M.C.; Juiz, J.M. Upregulation of insulin-like growth factor and interleukin 1β occurs in neurons but not in glial cells in the cochlear nucleus following cochlear ablation. J. Comp. Neurol. 2013, 521, 3478–3499. [Google Scholar] [CrossRef] [PubMed]
- Fuentes-Santamaría, V.; Alvarado, J.C.; Melgar-Rojas, P.; Gabaldón-Ull, M.C.; Miller, J.M.; Juiz, J.M. The role of glia in the peripheral and central auditory system following noise overexposure: Contribution of TNF-α and IL-1β to the pathogenesis of hearing loss. Front. Neuroanat. 2017, 11, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baizer, J.S.; Wong, K.M.; Manohar, S.; Hayes, S.H.; Ding, D.; Dingman, R.; Salvi, R.J. Effects of acoustic trauma on the auditory system of the rat: The role of microglia. Neuroscience 2015, 303, 299–311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, N.; März, S.; Plagemann, T.; Cao, J.; Schnittler, H.J.; Eter, N.; Heiduschka, P. Occurrence of transmembrane protein 119 in the retina is not restricted to the microglia: An immunohistochemical study. Transl. Vis. Sci. Technol. 2019, 8, 29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perl, K.; Ushakov, K.; Pozniak, Y.; Yizhar-Barnea, O.; Bhonker, Y.; Shivatzki, S.; Geiger, T.; Avraham, K.B.; Shamir, R. Reduced changes in protein compared to mRNA levels across non-proliferating tissues. BMC Genom. 2017, 18, 305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujioka, M.; Okano, H.; Ogawa, K. Inflammatory and immune responses in the cochlea: Potential therapeutic targets for sensorineural hearing loss. Front. Pharmacol. 2014, 5, 287. [Google Scholar] [CrossRef] [Green Version]
- Kitajiri, S.; Miyamoto, T.; Mineharu, A.; Sonoda, N.; Furuse, K.; Hata, M.; Sasaki, H.; Mori, Y.; Kubota, T.; Ito, J.; et al. Compartmentalization established by claudin-11-based tight junctions in stria vascularis is required for hearing through generation of endocochlear potential. J. Cell Sci. 2004, 117, 5087–5096. [Google Scholar] [CrossRef] [Green Version]
- Gow, A.; Davies, C.; Southwood, C.M.; Frolenkov, G.; Chrustowski, M.; Ng, L.; Yamauchi, D.; Marcus, D.C.; Kachar, B. Deafness in Claudin 11-null mice reveals the critical contribution of basal cell tight junctions to stria vascularis function. J. Neurosci. 2004, 24, 7051–7062. [Google Scholar] [CrossRef] [Green Version]
- Zhang, N.; Cai, J.; Xu, L.; Wang, H.; Liu, W. Cisplatin-induced stria vascularis damage is associated with inflammation and fibrosis. Neural Plast. 2020, 2020, 8851525. [Google Scholar] [CrossRef]
- Breglio, A.M.; Rusheen, A.E.; Shide, E.D.; Fernandez, K.A.; Spielbauer, K.K.; McLachlin, K.M.; Hall, M.D.; Amable, L.; Cunningham, L.L. Cisplatin is retained in the cochlea indefinitely following chemotherapy. Nat. Commun. 2017, 8, 1654. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Ct Beta-Actin (Duplicate 1) | Ct Beta-Actin (Duplicate 2) | Average Ct (Beta-Actin) | Ct TMEM119 (Duplicate 1) | Ct TMEM119 (Duplicate 2) | Average Ct (TMEM119) | ΔCt (Average Ct TMEM119—Average Ct Beta-Actin) |
|---|---|---|---|---|---|---|---|
| cochlea 1 | 20.69 | 20.33 | 20.51 | 32.95 | 32.20 | 32.58 | 12.07 |
| cochlea 2 | 20.28 | 20.28 | 20.28 | 32.17 | 31.77 | 31.97 | 11.69 |
| cochlea 3 | 20.13 | 20.06 | 20.10 | 32.00 | 31.47 | 31.74 | 11.64 |
| brain 1 | 20.36 | 18.95 | 19.66 | 36.62 | 36.19 | 36.41 | 16.75 |
| brain 2 | 19.44 | 19.23 | 19.34 | 36.97 | 35.61 | 36.29 | 16.96 |
| brain 3 | 19.44 | 19.20 | 19.32 | 36.24 | 35.97 | 36.11 | 16.79 |
| spleen 1 | 19.46 | 18.98 | 19.22 | 34.46 | 32.88 | 33.67 | 14.45 |
| spleen 2 | 19.62 | 19.50 | 19.56 | 35.91 | 34.77 | 35.34 | 15.78 |
| spleen 3 | 19.51 | 19.36 | 19.44 | 36.37 | 35.23 | 35.80 | 16.37 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bassiouni, M.; Smorodchenko, A.; Olze, H.; Szczepek, A.J. Identification and Characterization of TMEM119-Positive Cells in the Postnatal and Adult Murine Cochlea. Brain Sci. 2023, 13, 516. https://doi.org/10.3390/brainsci13030516
Bassiouni M, Smorodchenko A, Olze H, Szczepek AJ. Identification and Characterization of TMEM119-Positive Cells in the Postnatal and Adult Murine Cochlea. Brain Sciences. 2023; 13(3):516. https://doi.org/10.3390/brainsci13030516
Chicago/Turabian StyleBassiouni, Mohamed, Alina Smorodchenko, Heidi Olze, and Agnieszka J. Szczepek. 2023. "Identification and Characterization of TMEM119-Positive Cells in the Postnatal and Adult Murine Cochlea" Brain Sciences 13, no. 3: 516. https://doi.org/10.3390/brainsci13030516