

The Inhibition Effects of Sodium Nitroprusside on the Survival of Differentiated Neural Stem Cells through the p38 Pathway
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Methods and Materials
2.1. Isolation and Culture of Cerebral Cortex NSCs
2.2. Cell Viability Assay
2.3. Immunofluorescence Staining
2.4. Western Blotting Analysis
2.5. Statistical Analysis
3. Results
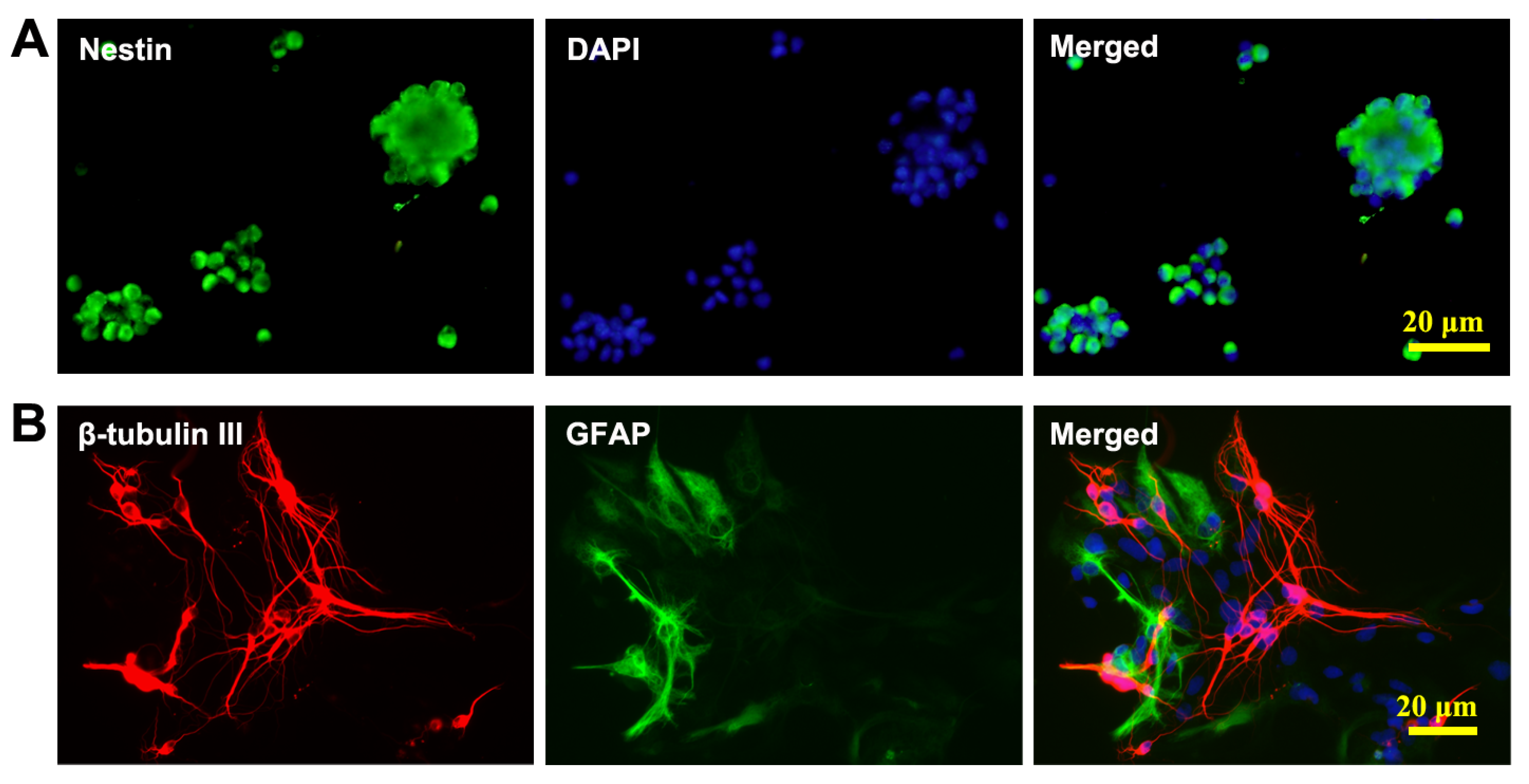
3.1. Culture and Identification of NSCs
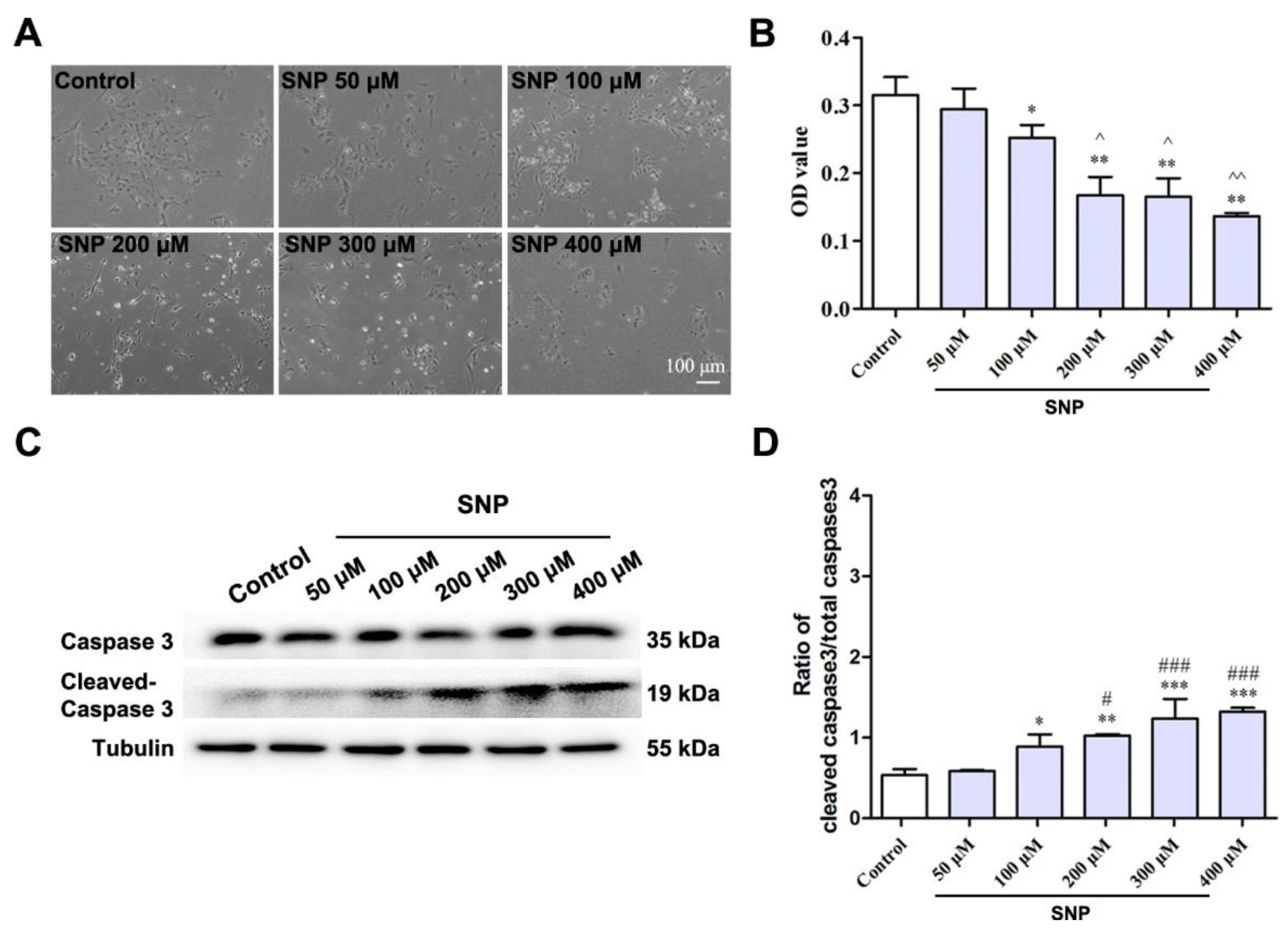
3.2. SNP Decreased the Survival of Differentiated NSCs
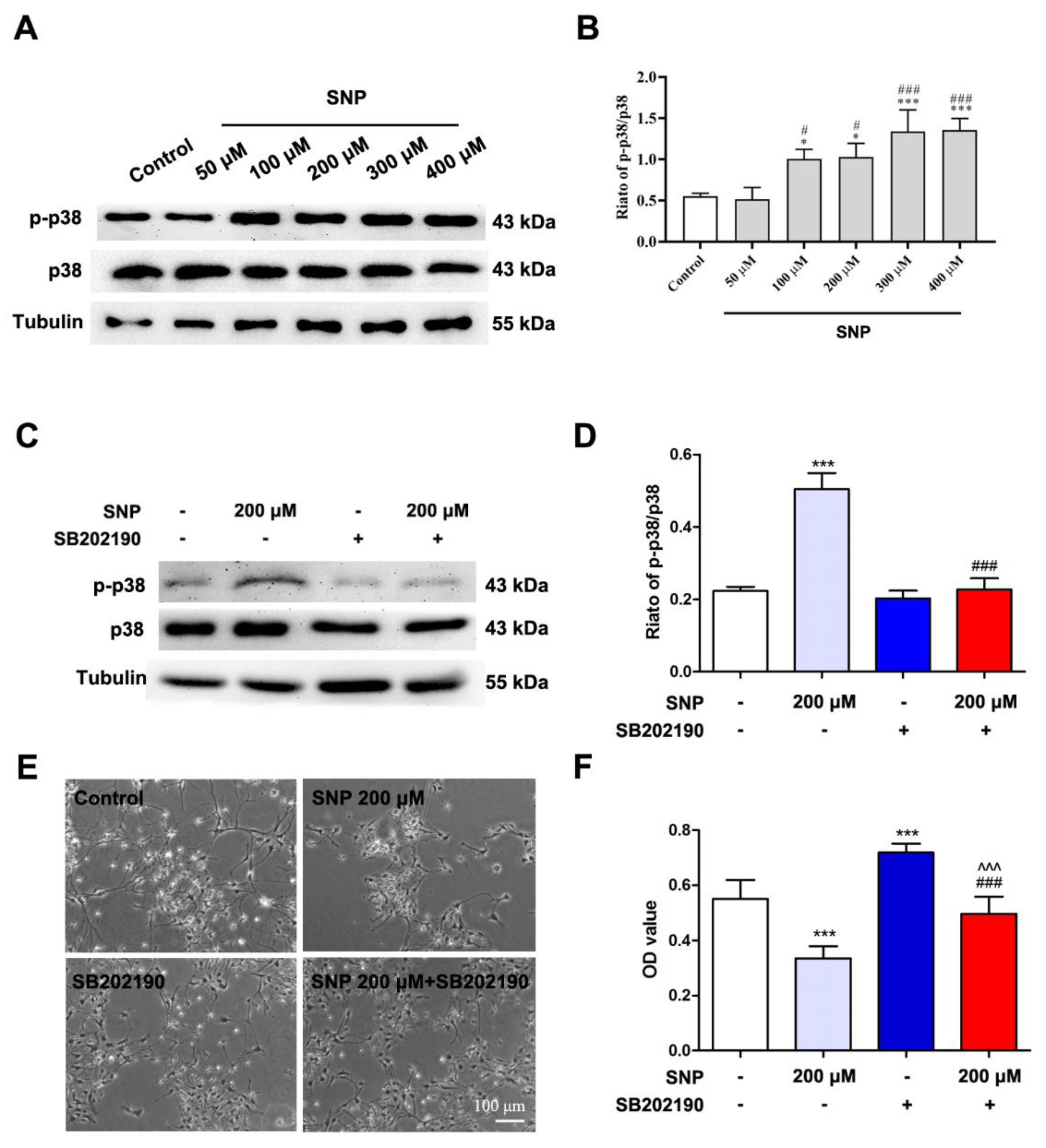
3.3. SNP Affected the Survival of Differentiated NSCs through p38
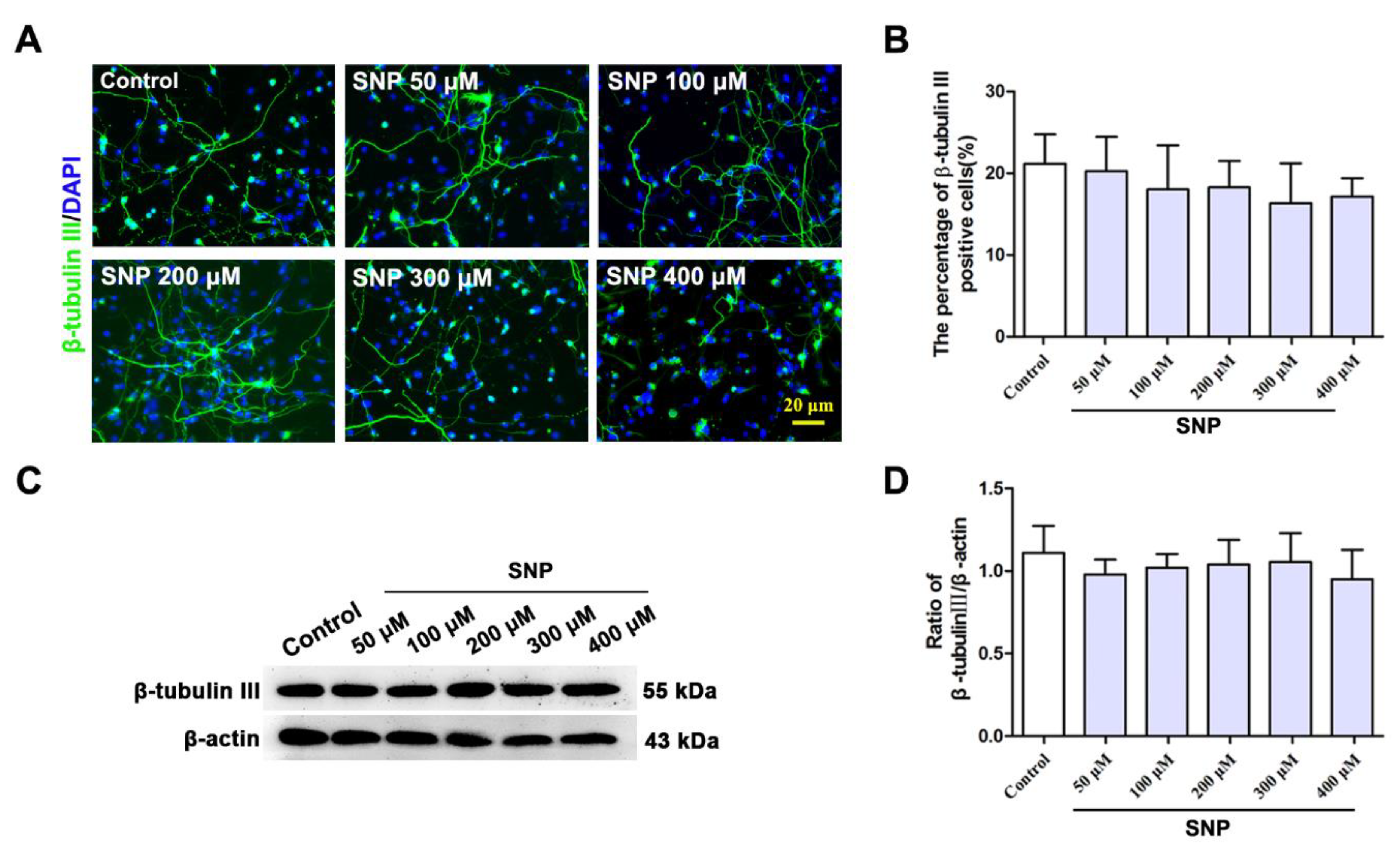
3.4. SNP Had No Effect on the Neuronal Differentiation of NSCs
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Carreira, B.P.; Morte, M.I.; Inacio, A.; Costa, G.; Rosmaninho-Salgado, J.; Agasse, F.; Carmo, A.; Couceiro, P.; Brundin, P.; Ambrosio, A.F.; et al. Nitric oxide stimulates the proliferation of neural stem cells bypassing the epidermal growth factor receptor. Stem Cells 2010, 28, 1219–1230. [Google Scholar] [CrossRef]
- Moreno-Lopez, B.; Romero-Grimaldi, C.; Noval, J.A.; Murillo-Carretero, M.; Matarredona, E.R.; Estrada, C. Nitric oxide is a physiological inhibitor of neurogenesis in the adult mouse subventricular zone and olfactory bulb. J. Neurosci. 2004, 24, 85–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, A.; Wang, S.; Cai, J.; Rao, M.S.; Mattson, M.P. Nitric oxide acts in a positive feedback loop with BDNF to regulate neural progenitor cell proliferation and differentiation in the mammalian brain. Dev. Biol. 2003, 258, 319–333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Contestabile, A.; Ciani, E. Role of nitric oxide in the regulation of neuronal proliferation, survival and differentiation. Neurochem. Int. 2004, 45, 903–914. [Google Scholar] [CrossRef] [PubMed]
- Scholz, N.L.; Chang, E.S.; Graubard, K.; Truman, J.W. The NO/cGMP pathway and the development of neural networks in postembryonic lobsters. J. Neurobiol. 1998, 34, 208–226. [Google Scholar] [CrossRef]
- Hindley, S.; Juurlink, B.H.; Gysbers, J.W.; Middlemiss, P.J.; Herman, M.A.; Rathbone, M.P. Nitric oxide donors enhance neurotrophin-induced neurite outgrowth through a cGMP-dependent mechanism. J. Neurosci. Res. 1997, 47, 427–439. [Google Scholar] [CrossRef]
- Enikolopov, G.; Banerji, J.; Kuzin, B. Nitric oxide and Drosophila development. Cell Death Differ. 1999, 6, 956–963. [Google Scholar] [CrossRef] [Green Version]
- Lin, I.C.; Smartt, J.M., Jr.; Nah, H.D.; Ischiropoulos, H.; Kirschner, R.E. Nitric oxide stimulates proliferation and differentiation of fetal calvarial osteoblasts and dural cells. Plast. Reconstr. Surg. 2008, 121, 1554–1566. [Google Scholar] [CrossRef]
- Arnhold, S.; Fassbender, A.; Klinz, F.J.; Kruttwig, K.; Lohnig, B.; Andressen, C.; Addicks, K. NOS-II is involved in early differentiation of murine cortical, retinal and ES cell-derived neurons-an immunocytochemical and functional approach. Int. J. Dev. Neurosci. 2002, 20, 83–92. [Google Scholar] [CrossRef]
- Champlin, D.T.; Truman, J.W. Ecdysteroid coordinates optic lobe neurogenesis via a nitric oxide signaling pathway. Development 2000, 127, 3543–3551. [Google Scholar] [CrossRef]
- Sulz, L.; Bacigalupo, J. Role of nitric oxide during neurogenesis in the olfactory epithelium. Biol. Res. 2006, 39, 589–599. [Google Scholar] [CrossRef] [PubMed]
- Carreira, B.P.; Morte, M.I.; Santos, A.I.; Lourenco, A.S.; Ambrosio, A.F.; Carvalho, C.M.; Araujo, I.M. Nitric oxide from inflammatory origin impairs neural stem cell proliferation by inhibiting epidermal growth factor receptor signaling. Front. Cell. Neurosci. 2014, 8, 343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, L.; Zhang, Y.; Chen, Y.; Zhu, J.; Yang, Y.; Zhang, H.L. Role of Microglia in Neurological Disorders and Their Potentials as a Therapeutic Target. Mol. Neurobiol. 2017, 54, 7567–7584. [Google Scholar] [CrossRef] [PubMed]
- Zhong, H.J.; Liu, L.J.; Chong, C.M.; Lu, L.; Wang, M.; Chan, D.S.; Chan, P.W.; Lee, S.M.; Ma, D.L.; Leung, C.H. Discovery of a natural product-like iNOS inhibitor by molecular docking with potential neuroprotective effects in vivo. PLoS ONE 2014, 9, e92905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grossi, L.; D’Angelo, S. Sodium nitroprusside: Mechanism of NO release mediated by sulfhydryl-containing molecules. J. Med. Chem. 2005, 48, 2622–2626. [Google Scholar] [CrossRef]
- Zoupa, E.; Pitsikas, N. The Nitric Oxide (NO) Donor Sodium Nitroprusside (SNP) and Its Potential for the Schizophrenia Therapy: Lights and Shadows. Molecules 2021, 26, 3196. [Google Scholar] [CrossRef]
- Jiao, Q.; Li, X.; An, J.; Zhang, Z.; Chen, X.; Tan, J.; Zhang, P.; Lu, H.; Liu, Y. Cell-Cell Connection Enhances Proliferation and Neuronal Differentiation of Rat Embryonic Neural Stem/Progenitor Cells. Front. Cell. Neurosci. 2017, 11, 200. [Google Scholar] [CrossRef] [Green Version]
- Gong, B.; Jiao, L.; Du, X.; Li, Y.; Bi, M.; Jiao, Q.; Jiang, H. Ghrelin promotes midbrain neural stem cells differentiation to dopaminergic neurons through Wnt/beta-catenin pathway. J. Cell. Physiol. 2020, 235, 8558–8570. [Google Scholar] [CrossRef]
- Shen, Q.; Wang, Y.; Kokovay, E.; Lin, G.; Chuang, S.M.; Goderie, S.K.; Roysam, B.; Temple, S. Adult SVZ stem cells lie in a vascular niche: A quantitative analysis of niche cell-cell interactions. Cell Stem Cell 2008, 3, 289–300. [Google Scholar] [CrossRef] [Green Version]
- Llorens-Bobadilla, E.; Martin-Villalba, A. Adult NSC diversity and plasticity: The role of the niche. Curr. Opin. Neurobiol. 2017, 42, 68–74. [Google Scholar] [CrossRef]
- Gray, W.P.; Cheung, A. Nitric oxide regulation of adult neurogenesis. Vitam. Horm. 2014, 96, 59–77. [Google Scholar] [CrossRef]
- Madhusoodanan, K.S.; Murad, F. NO-cGMP signaling and regenerative medicine involving stem cells. Neurochem. Res. 2007, 32, 681–694. [Google Scholar] [CrossRef] [PubMed]
- Tropea, M.R.; Gulisano, W.; Vacanti, V.; Arancio, O.; Puzzo, D.; Palmeri, A. Nitric oxide/cGMP/CREB pathway and amyloid-beta crosstalk: From physiology to Alzheimer’s disease. Free Radic. Biol. Med. 2022, 193, 657–668. [Google Scholar] [CrossRef] [PubMed]
- Brown, G.C. Nitric oxide and neuronal death. Nitric Oxide 2010, 23, 153–165. [Google Scholar] [CrossRef] [PubMed]
- Cokic, V.P.; Schechter, A.N. Effects of nitric oxide on red blood cell development and phenotype. Curr. Top. Dev. Biol. 2008, 82, 169–215. [Google Scholar] [CrossRef]
- Brenman, J.E.; Bredt, D.S. Synaptic signaling by nitric oxide. Curr. Opin. Neurobiol. 1997, 7, 374–378. [Google Scholar] [CrossRef]
- Wang, X.; Wang, H.; Li, H. Inhibition of iNOS ameliorates traumatic stress-induced deficits in synaptic plasticity and memory. Psychiatry Res. 2018, 268, 413–418. [Google Scholar] [CrossRef]
- Stewart, V.C.; Heslegrave, A.J.; Brown, G.C.; Clark, J.B.; Heales, S.J. Nitric oxide-dependent damage to neuronal mitochondria involves the NMDA receptor. Eur. J. Neurosci. 2002, 15, 458–464. [Google Scholar] [CrossRef]
- Li, N.; Sul, J.Y.; Haydon, P.G. A calcium-induced calcium influx factor, nitric oxide, modulates the refilling of calcium stores in astrocytes. J. Neurosci. 2003, 23, 10302–10310. [Google Scholar] [CrossRef] [Green Version]
- Phan Duy, A.; Pham, H.; Pansiot, J.; Gressens, P.; Charriaut-Marlangue, C.; Baud, O. Nitric Oxide Pathway and Proliferation of Neural Progenitors in the Neonatal Rat. Dev. Neurosci. 2015, 37, 417–427. [Google Scholar] [CrossRef]
- Boullerne, A.I.; Benjamins, J.A. Nitric oxide synthase expression and nitric oxide toxicity in oligodendrocytes. Antioxid. Redox Signal. 2006, 8, 967–980. [Google Scholar] [CrossRef] [PubMed]
- Kapralov, A.A.; Yang, Q.; Dar, H.H.; Tyurina, Y.Y.; Anthonymuthu, T.S.; Kim, R.; St Croix, C.M.; Mikulska-Ruminska, K.; Liu, B.; Shrivastava, I.H.; et al. Redox lipid reprogramming commands susceptibility of macrophages and microglia to ferroptotic death. Nat. Chem. Biol. 2020, 16, 278–290. [Google Scholar] [CrossRef] [PubMed]
- Stefano, G.B.; Kim, E.; Liu, Y.; Zhu, W.; Casares, F.; Mantione, K.; Jones, D.A.; Cadet, P. Nitric oxide modulates microglial activation. Med. Sci. Monit. 2004, 10, BR17–BR22. [Google Scholar] [PubMed]
- Dimmeler, S.; Zeiher, A.M. Nitric oxide-an endothelial cell survival factor. Cell Death Differ. 1999, 6, 964–968. [Google Scholar] [CrossRef] [Green Version]
- Lei, J.; Vodovotz, Y.; Tzeng, E.; Billiar, T.R. Nitric oxide, a protective molecule in the cardiovascular system. Nitric Oxide 2013, 35, 175–185. [Google Scholar] [CrossRef] [PubMed]
- Ziche, M.; Morbidelli, L.; Masini, E.; Amerini, S.; Granger, H.J.; Maggi, C.A.; Geppetti, P.; Ledda, F. Nitric oxide mediates angiogenesis in vivo and endothelial cell growth and migration in vitro promoted by substance P. J. Clin. Investig. 1994, 94, 2036–2044. [Google Scholar] [CrossRef]
- Wang, F.; Qin, K.; Wang, K.; Wang, H.; Liu, Q.; Qian, M.; Chen, S.; Sun, Y.; Hou, J.; Wei, Y.; et al. Nitric oxide improves regeneration and prevents calcification in bio-hybrid vascular grafts via regulation of vascular stem/progenitor cells. Cell Rep. 2022, 39, 110981. [Google Scholar] [CrossRef]
- Luo, C.X.; Jin, X.; Cao, C.C.; Zhu, M.M.; Wang, B.; Chang, L.; Zhou, Q.G.; Wu, H.Y.; Zhu, D.Y. Bidirectional regulation of neurogenesis by neuronal nitric oxide synthase derived from neurons and neural stem cells. Stem Cells 2010, 28, 2041–2052. [Google Scholar] [CrossRef]
- Reif, A.; Schmitt, A.; Fritzen, S.; Chourbaji, S.; Bartsch, C.; Urani, A.; Wycislo, M.; Mossner, R.; Sommer, C.; Gass, P.; et al. Differential effect of endothelial nitric oxide synthase (NOS-III) on the regulation of adult neurogenesis and behaviour. Eur. J. Neurosci. 2004, 20, 885–895. [Google Scholar] [CrossRef]
- Cheung, A.; Newland, P.L.; Zaben, M.; Attard, G.S.; Gray, W.P. Intracellular nitric oxide mediates neuroproliferative effect of neuropeptide y on postnatal hippocampal precursor cells. J. Biol. Chem. 2012, 287, 20187–20196. [Google Scholar] [CrossRef] [Green Version]
- Park, C.; Sohn, Y.; Shin, K.S.; Kim, J.; Ahn, H.; Huh, Y. The chronic inhibition of nitric oxide synthase enhances cell proliferation in the adult rat hippocampus. Neurosci. Lett. 2003, 339, 9–12. [Google Scholar] [CrossRef] [PubMed]
- Bai, J.; Ramos, R.L.; Ackman, J.B.; Thomas, A.M.; Lee, R.V.; LoTurco, J.J. RNAi reveals doublecortin is required for radial migration in rat neocortex. Nat. Neurosci. 2003, 6, 1277–1283. [Google Scholar] [CrossRef] [PubMed]
- Li, J.T.; Somasundaram, C.; Bian, K.; Xiong, W.; Mahmooduddin, F.; Nath, R.K.; Murad, F. Nitric oxide signaling and neural stem cell differentiation in peripheral nerve regeneration. Eplasty 2010, 10, e42. [Google Scholar]
- Mokry, R.L.; O’Brien, B.S.; Adelman, J.W.; Rosas, S.; Schumacher, M.L.; Ebert, A.D.; Terhune, S.S. Nitric Oxide Attenuates Human Cytomegalovirus Infection yet Disrupts Neural Cell Differentiation and Tissue Organization. J. Virol. 2022, 96, e0012622. [Google Scholar] [CrossRef]
- Wang, J.; Liu, J.; Zhou, R.; Ding, X.; Zhang, Q.; Zhang, C.; Li, L. Zika virus infected primary microglia impairs NPCs proliferation and differentiation. Biochem. Biophys. Res. Commun. 2018, 497, 619–625. [Google Scholar] [CrossRef]
- Rabinovich, D.; Yaniv, S.P.; Alyagor, I.; Schuldiner, O. Nitric Oxide as a Switching Mechanism between Axon Degeneration and Regrowth during Developmental Remodeling. Cell 2016, 164, 170–182. [Google Scholar] [CrossRef] [Green Version]
- Sulz, L.; Astorga, G.; Bellette, B.; Iturriaga, R.; Mackay-Sim, A.; Bacigalupo, J. Nitric oxide regulates neurogenesis in adult olfactory epithelium in vitro. Nitric Oxide 2009, 20, 238–252. [Google Scholar] [CrossRef]
- Bredt, D.S.; Snyder, S.H. Transient nitric oxide synthase neurons in embryonic cerebral cortical plate, sensory ganglia, and olfactory epithelium. Neuron 1994, 13, 301–313. [Google Scholar] [CrossRef]
- Santacana, M.; Uttenthal, L.O.; Bentura, M.L.; Fernandez, A.P.; Serrano, J.; Martinez de Velasco, J.; Alonso, D.; Martinez-Murillo, R.; Rodrigo, J. Expression of neuronal nitric oxide synthase during embryonic development of the rat cerebral cortex. Brain Res. Dev. Brain Res. 1998, 111, 205–222. [Google Scholar] [CrossRef] [PubMed]
- Radi, R. Oxygen radicals, nitric oxide, and peroxynitrite: Redox pathways in molecular medicine. Proc. Natl. Acad. Sci. USA 2018, 115, 5839–5848. [Google Scholar] [CrossRef] [Green Version]
- Kim, T.; Kim, H.J.; Choi, W.; Lee, Y.M.; Pyo, J.H.; Lee, J.; Kim, J.; Kim, J.; Kim, J.H.; Kim, C.; et al. Deep brain stimulation by blood-brain-barrier-crossing piezoelectric nanoparticles generating current and nitric oxide under focused ultrasound. Nat. Biomed. Eng. 2022. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, Y.; Ikeuchi, T.; Hatanaka, H. Brain-derived neurotrophic factor accelerates nitric oxide donor-induced apoptosis of cultured cortical neurons. J. Neurochem. 2000, 75, 494–502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, C.; Wu, C.; Shu, P.; Liu, T.; Li, H.; Beuve, A. Soluble guanylyl cyclase mediates noncanonical nitric oxide signaling by nitrosothiol transfer under oxidative stress. Redox Biol. 2022, 55, 102425. [Google Scholar] [CrossRef] [PubMed]
- Duran, W.N.; Beuve, A.V.; Sanchez, F.A. Nitric oxide, S-nitrosation, and endothelial permeability. IUBMB Life 2013, 65, 819–826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parrish, M.C.; Chaim, I.A.; Nagel, Z.D.; Tannenbaum, S.R.; Samson, L.D.; Engelward, B.P. Nitric oxide induced S-nitrosation causes base excision repair imbalance. DNA Repair 2018, 68, 25–33. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiao, L.; Xu, T.; Du, X.; Chen, X.; Jiao, Q.; Jiang, H. The Inhibition Effects of Sodium Nitroprusside on the Survival of Differentiated Neural Stem Cells through the p38 Pathway. Brain Sci. 2023, 13, 438. https://doi.org/10.3390/brainsci13030438
Jiao L, Xu T, Du X, Chen X, Jiao Q, Jiang H. The Inhibition Effects of Sodium Nitroprusside on the Survival of Differentiated Neural Stem Cells through the p38 Pathway. Brain Sciences. 2023; 13(3):438. https://doi.org/10.3390/brainsci13030438
Chicago/Turabian StyleJiao, Lingling, Tongying Xu, Xixun Du, Xi Chen, Qian Jiao, and Hong Jiang. 2023. "The Inhibition Effects of Sodium Nitroprusside on the Survival of Differentiated Neural Stem Cells through the p38 Pathway" Brain Sciences 13, no. 3: 438. https://doi.org/10.3390/brainsci13030438