

Moderate-Intensity Intermittent Training Alters the DNA Methylation Pattern of PDE4D Gene in Hippocampus to Improve the Ability of Spatial Learning and Memory in Aging Rats Reduced by D-Galactose
 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Study Design
2.2. Establishment of the Aging Model
2.3. Exercise Protocols
2.4. Morris Water Maze Behavioral Test
2.5. Tissue Preparation
2.6. Transmission Electron Microscopy Assay
2.7. ELISA
2.8. Gene expression Analysis
2.9. Western Blot Analysis
2.10. DNA Methylation Analysis
2.11. Statistical Analysis
3. Results
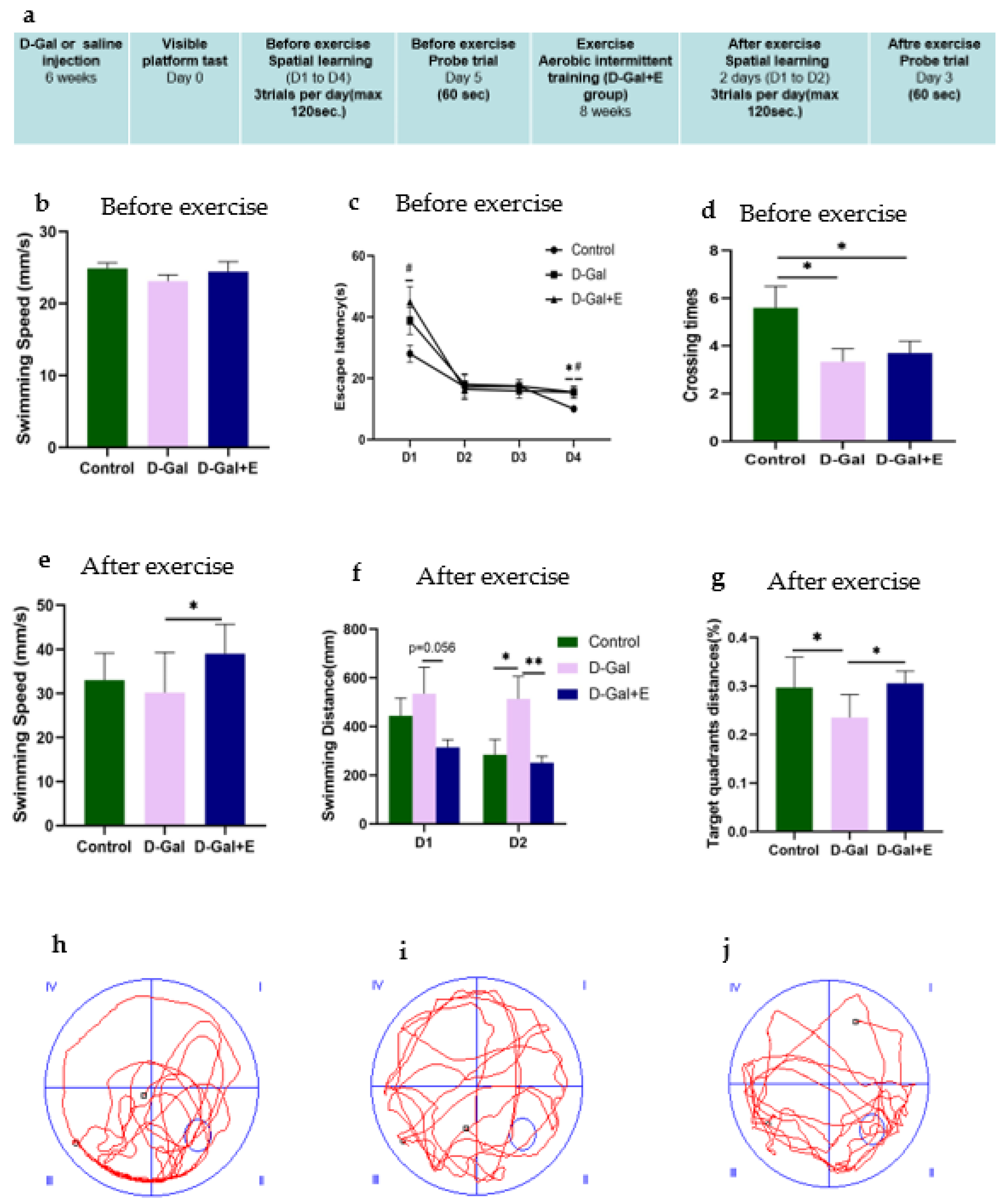
3.1. Eight Weeks MIIT Improves Spatial Learning and Memory in D-Gal-Induced Aging Rats
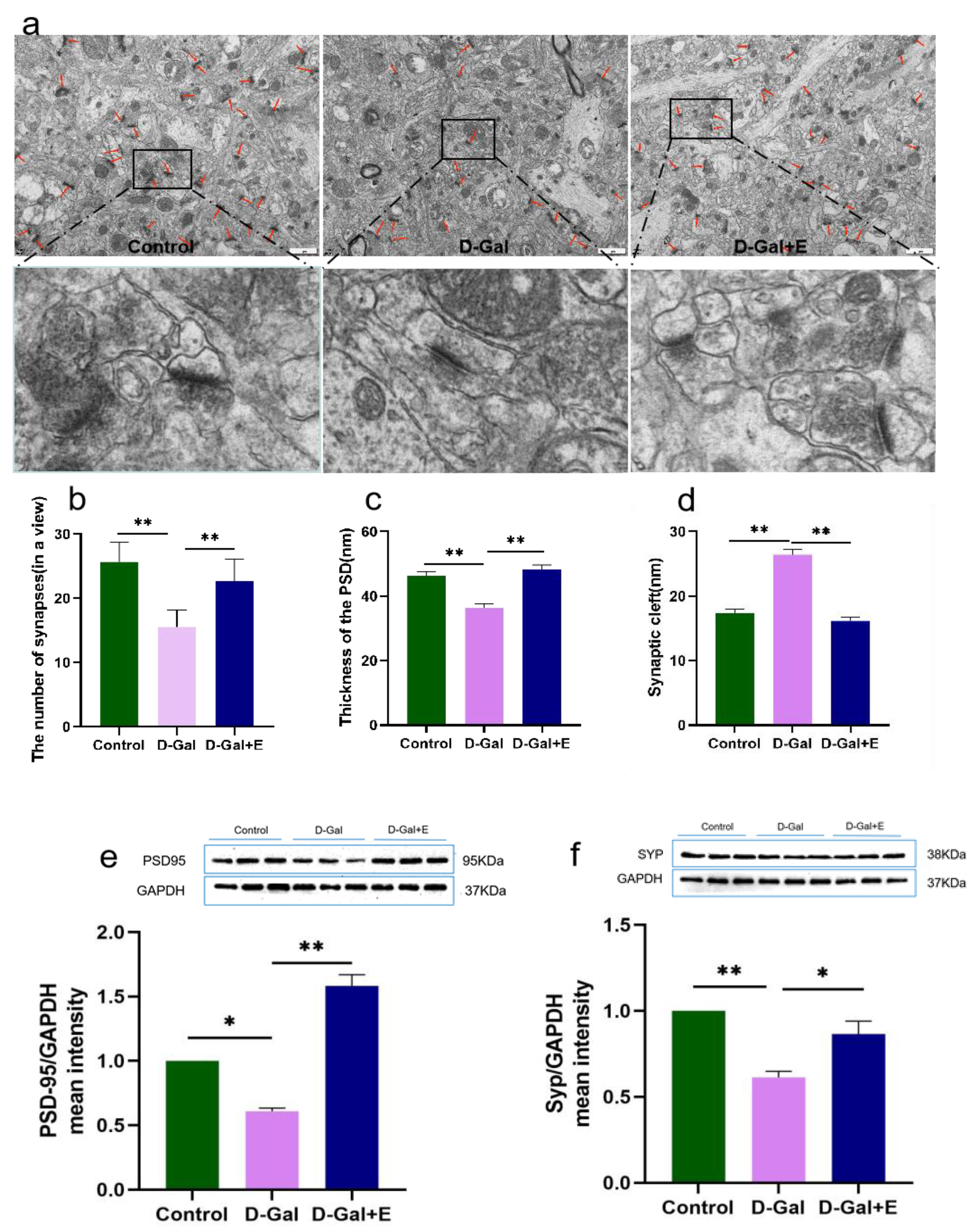
3.2. Eight Weeks MIIT Improves the Ultrastructure of Hippocampus Synapses and Up-Regulates the Expression of Synaptic Associated Proteins in Aging Rats
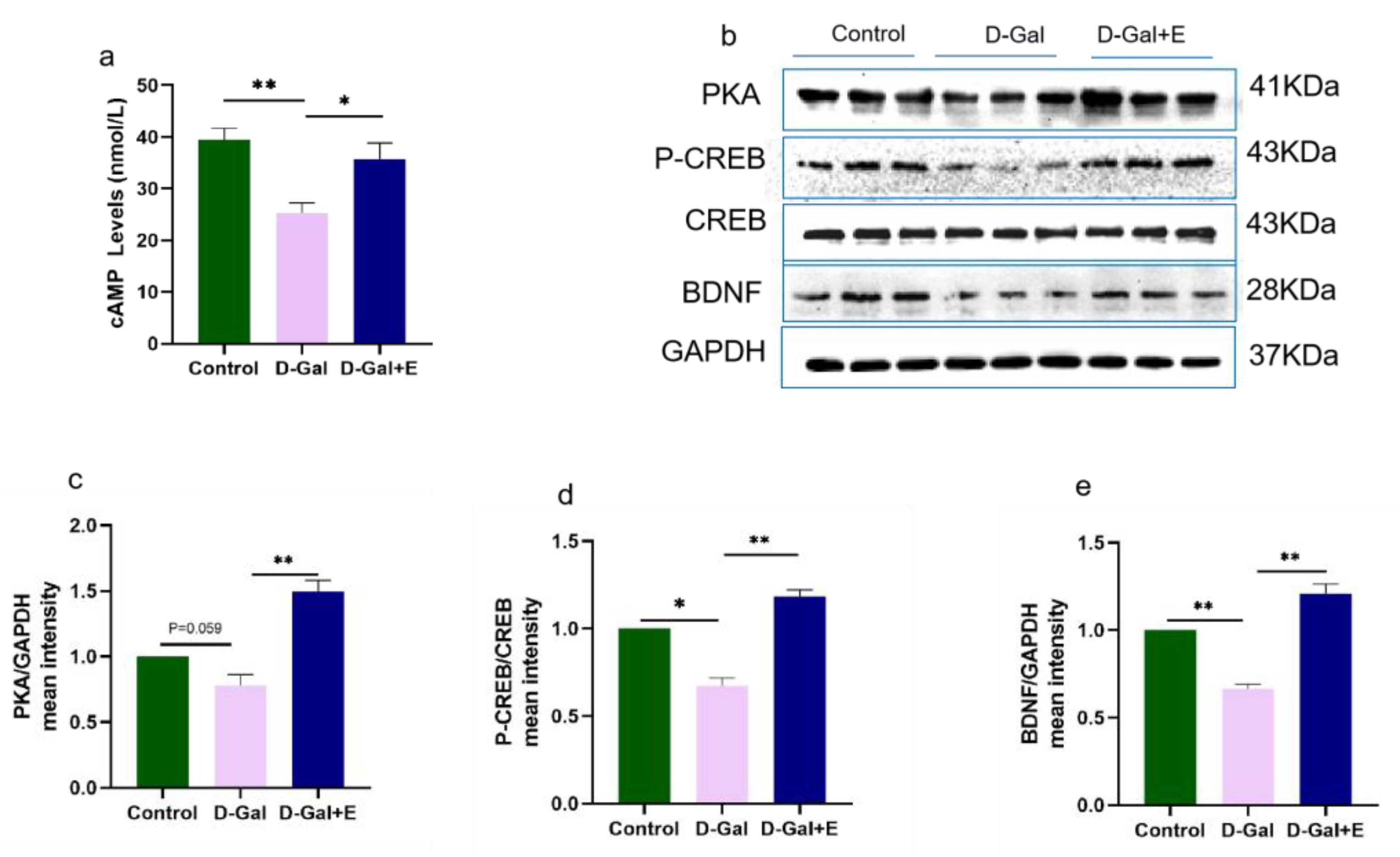
3.3. MIIT Promotes cAMP/PKA/CREB/BDNF Signaling Pathway up-Regulation in Aging Rats
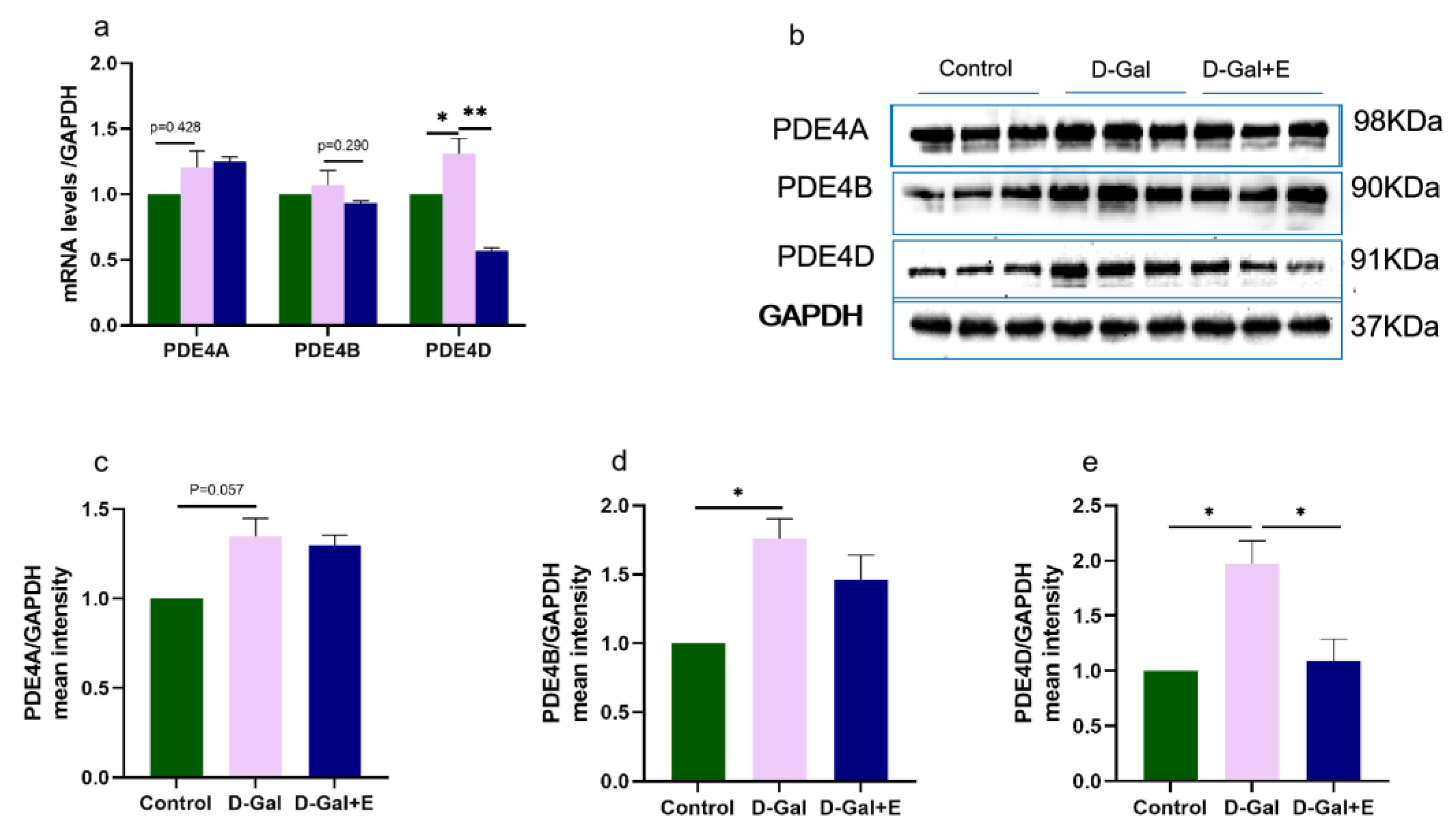
3.4. MIIT Effectively Reduced PDE4D Expression but Not PDE4A and PDE4B
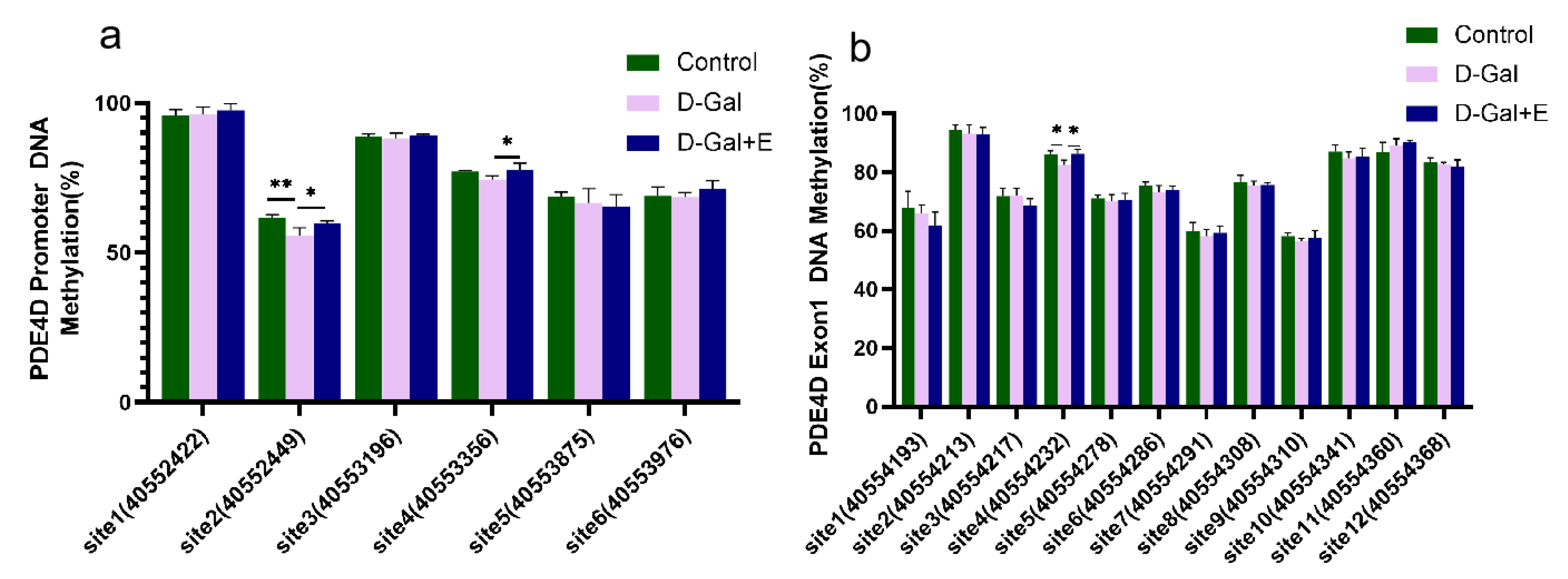
3.5. MIIT Regulates Methylation Levels in PDE4D Promoter and Exon1 Regions
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Adcock, M.; Fankhauser, M.; Post, J.; Lutz, K.; Zizlsperger, L.; Luft, A.R.; Guimarães, V.; Schättin, A.; De Bruin, E.D. Effects of an In-home Multicomponent Exergame Training on Physical Functions, Cognition, and Brain Volume of Older Adults: A Randomized Controlled Trial. Front. Med. 2019, 6, 321. [Google Scholar] [CrossRef] [PubMed]
- Peters, R. Ageing and the brain. Postgrad. Med. J. 2006, 82, 84–88. [Google Scholar] [CrossRef] [PubMed]
- Fotuhi, M.; Do, D.; Jack, C. Modifiable factors that alter the size of the hippocampus with ageing. Nat. Rev. Neurol. 2012, 8, 189–202. [Google Scholar] [CrossRef] [PubMed]
- Geinisman, Y.; Detoledo-Morrell, L.; Morrell, F.; Heller, R.E. Hippocampal markers of age-related memory dysfunction: Behavioral, electrophysiological and morphological perspectives. Prog. Neurobiol. 1995, 45, 223–252. [Google Scholar] [CrossRef] [PubMed]
- Ho, V.M.; Lee, J.A.; Martin, K.C. The cell biology of synaptic plasticity. Science 2011, 334, 623–628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhat, A.; Ray, B.; Mahalakshmi, A.M.; Tuladhar, S.; Nandakumar, D.; Srinivasan, M.; Essa, M.M.; Chidambaram, S.B.; Guillemin, G.J.; Sakharkar, M.K. Phosphodiesterase-4 enzyme as a therapeutic target in neurological disorders. Pharmacol. Res. 2020, 160, 105078. [Google Scholar] [CrossRef]
- Blokland, A.; Van Duinen, M.A.; Sambeth, A.; Heckman, P.R.; Tsai, M.; Lahu, G.; Uz, T.; Prickaerts, J. Acute treatment with the PDE4 inhibitor roflumilast improves verbal word memory in healthy old individuals: A double-blind placebo-controlled study. Neurobiol. Aging 2019, 77, 37–43. [Google Scholar] [CrossRef]
- Vogel, E.W., III; Morales, F.N.; Meaney, D.F.; Bass, C.R.; Morrison, B., III. Phosphodiesterase-4 inhibition restored hippocampal long term potentiation after primary blast. Exp. Neurol. 2017, 293, 91–100. [Google Scholar] [CrossRef] [Green Version]
- Peters, M.; Bletsch, M.; Stanley, J.; Wheeler, D.; Scott, R.; Tully, T. The PDE4 inhibitor HT-0712 improves hippocampus-dependent memory in aged mice. Neuropsychopharmacology 2014, 39, 2938–2948. [Google Scholar] [CrossRef] [Green Version]
- Wiescholleck, V.; Manahan-Vaughan, D. PDE4 inhibition enhances hippocampal synaptic plasticity in vivo and rescues MK801-induced impairment of long-term potentiation and object recognition memory in an animal model of psychosis. Transl. Psychiatry 2012, 2, e89. [Google Scholar] [CrossRef] [Green Version]
- Titus, D.; Sakurai, A.; Kang, Y.; Furones, C.; Jergova, S.; Santos, R.; Sick, T.J.; Atkins, C.M. Phosphodiesterase inhibition rescues chronic cognitive deficits induced by traumatic brain injury. J. Neurosci. 2013, 33, 5216–5226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Y.; Zhu, N.; Xu, W.; Ye, H.; Liu, K.; Wu, F.; Zhang, M.; Ding, Y.; Zhang, C.; Zhang, H.; et al. Inhibition of Phosphodiesterase-4 Reverses Aβ-Induced Memory Impairment by Regulation of HPA Axis Related cAMP Signaling. Front. Aging Neurosci. 2018, 10, 204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolsius, Y.G.; Heckman, P.R.; Paraciani, C.; Wilhelm, S.; Raven, F.; Meijer, E.L.; Kas, M.J.; Ramirez, S.; Meerlo, P.; Havekes, R. Recovering object-location memories after sleep deprivation-induced amnesia. Curr. Biol. 2023, 33, 298–308.e5. [Google Scholar] [CrossRef] [PubMed]
- Peng, T.; Qi, B.; He, J.; Ke, H.; Shi, J. Advances in the Development of Phosphodiesterase-4 Inhibitors. J. Med. Chem. 2020, 63, 10594–10617. [Google Scholar] [CrossRef]
- Wang, H.; Zhang, F.F.; Xu, Y.; Fu, H.R.; Wang, X.D.; Wang, L.; Chen, W.; Xu, X.Y.; Gao, Y.F.; Zhang, J.G.; et al. The Phosphodiesterase-4 Inhibitor Roflumilast, a Potential Treatment for the Comorbidity of Memory Loss and Depression in Alzheimer’s Disease: A Preclinical Study in APP/PS1 Transgenic Mice. Int. J. Neuropsychopharmacol. 2020, 23, 700–711. [Google Scholar] [CrossRef]
- Paes, D.; Schepers, M.; Rombaut, B.; Hove, D.V.D.; Vanmierlo, T.; Prickaerts, J. The Molecular Biology of Phosphodiesterase 4 Enzymes as Pharmacological Targets: An Interplay of Isoforms, Conformational States, and Inhibitors. Pharmacol. Rev. 2021, 73, 1016–1049. [Google Scholar] [CrossRef]
- Liu, Z.; Liu, M.; Cao, Z.; Qiu, P.; Song, G. Phosphodiesterase-4 inhibitors: A review of current developments (2013–2021). Expert Opin. Ther. Pat. 2022, 32, 261–278. [Google Scholar] [CrossRef]
- Zhang, X.; Ll, J.; Gao, F. Exercise benefits cardiovascular health: From molecular mechanisms to clinical implications. Sci. Sin. Inf. 2022, 52, 1–16. (In Chinese) [Google Scholar] [CrossRef]
- Liang, J.; Wang, H.; Zeng, Y.; Qu, Y.; Liu, Q.; Zhao, F.; Duan, J.; Jiang, Y.; Li, S.; Ying, J.; et al. Physical exercise promotes brain remodeling by regulating epigenetics, neuroplasticity and neurotrophins. Rev. Neurosci. 2021, 32, 615–629. [Google Scholar] [CrossRef]
- Zhang, S.; Li, X.; Yuan, Q.; Wang, L. Effects of Aerobic Exercises on Synaptic Plasticity and Expression of PDE-4 in Hippocampus during the Aging Process of Rats. Chin. J. Sports Med. 2017, 36, 875–881. (In Chinese) [Google Scholar]
- Moreira, J.B.N.; Wohlwend, M.; Wisløff, U. Exercise and cardiac health: Physiological and molecular insights. Nat. Metab. 2020, 2, 829–839. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, M.; Mizuuchi, D.; Omura, K.; Lee, M.; Oharazawa, A.; Yook, J.S.; Inoue, K.; Soya, H. High-intensity Intermittent Training Enhances Spatial Memory and Hippocampal Neurogenesis Associated with BDNF Signaling in Rats. Cereb. Cortex 2021, 31, 4386–4397. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Fu, X.; Han, R.; Liu, X.; Zhao, X.; Wei, J. Neuroprotective Effect of HIIT against GFAP Hypertrophy through Mitochondrial Dynamics in APP/PS1 Mice. Oxid. Med. Cell. Longev. 2022, 2022, 1764589. [Google Scholar] [CrossRef] [PubMed]
- Aksoy, S.; Findikoglu, G.; Ardic, F.; Rota, S.; Dursunoglu, D. Effect of 10-Week Supervised Moderate-Intensity Intermittent vs. Continuous Aerobic Exercise Programs on Vascular Adhesion Molecules in Patients with Heart Failure. Am. J. Phys. Med. Rehabil. 2015, 94, 898–911. [Google Scholar]
- Alkahtani, S.A.; King, N.A.; Hills, A.P.; Byrne, N.M. Effect of interval training intensity on fat oxidation, blood lactate and the rate of perceived exertion in obese men. Springerplus 2013, 2, 532. [Google Scholar] [CrossRef] [Green Version]
- Rakobowchuk, M.; Harris, E.; Taylor, A.; Baliga, V.; Cubbon, R.M.; Rossiter, H.; Birch, K. Heavy and moderate interval exercise training alters low-flow-mediated constriction but does not increase circulating progenitor cells in healthy humans. Exp. Physiol. 2012, 97, 375–385. [Google Scholar] [CrossRef]
- Racil, G.; Ben Ounis, O.; Hammouda, O.; Kallel, A.; Zouhal, H.; Chamari, K.; Amri, M. Effects of high vs. moderate exercise intensity during interval training on lipids and adiponectin levels in obese young females. Eur. J. Appl. Physiol. 2013, 113, 2531–2540. [Google Scholar]
- Jiménez-Pavón, D.; Lavie, C.J. High-intensity intermittent training versus moderate-intensity intermittent training: Is it a matter of intensity or intermittent efforts? Br. J. Sports Med. 2017, 51, 1319–1320. [Google Scholar] [CrossRef]
- Shen, Y.X.; Xu, S.Y.; Wei, W.; Sun, X.-X.; Yang, J.; Liu, L.-H.; Dong, C. Melatonin reduces memory changes and neural oxidative damage in mice treated with D-galactose. J. Pineal Res. 2002, 32, 173–178. [Google Scholar] [CrossRef]
- Tian, J.; Ishibashi, K.; Reiser, K.; Grebe, R.; Biswal, S.; Gehlbach, P.; Handa, J.T. Advanced glycation endproduct-induced aging of the retinal pigment epithelium and choroid: A comprehensive transcriptional response. Proc. Natl. Acad. Sci. USA 2005, 102, 11846–11851. [Google Scholar] [CrossRef] [Green Version]
- Yoo, D.Y.; Kim, W.; Lee, C.H.; Na Shin, B.; Nam, S.M.; Choi, J.H.; Won, M.-H.; Yoon, Y.S.; Hwang, I.K. Melatonin improves D-galactose-induced aging effects on behavior, neurogenesis, and lipid peroxidation in the mouse dentate gyrus via increasing pCREB expression. J. Pineal Res. 2012, 52, 21–28. [Google Scholar] [CrossRef]
- Sun, K.; Yang, P.; Zhao, R.; Bai, Y.; Guo, Z. Matrine Attenuates D-Galactose-Induced Aging-Related Behavior in Mice via Inhibition of Cellular Senescence and Oxidative Stress. Oxid. Med. Cell. Longev. 2018, 2018, 7108604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Høydal, M.A.; Wisløff, U.; Kemi, O.J.; Ellingsen, Ø. Running speed and maximal oxygen uptake in rats and mice: Practical implications for exercise training. Eur. J. Cardiovasc. Prev. Rehabil. 2007, 14, 753–760. [Google Scholar] [CrossRef] [PubMed]
- Vorhees, C.V.; Williams, M.T. Morris water maze: Procedures for assessing spatial and related forms of learning and memory. Nat. Protoc. 2006, 1, 848–858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haider, S.; Liaquat, L.; Shahzad, S.; Sadir, S.; Madiha, S.; Batool, Z.; Tabassum, S.; Saleem, S.; Naqvi, F.; Perveen, T. A high dose of short term exogenous D-galactose administration in young male rats produces symptoms simulating the natural aging process. Life Sci. 2015, 124, 110–119. [Google Scholar] [CrossRef] [PubMed]
- Chiroma, S.M.; Moklas, M.A.M.; Taib, C.N.M.; Baharuldin, M.T.H.; Amon, Z. D-galactose and aluminium chloride induced rat model with cognitive impairments. Biomed. Pharmacother. 2018, 103, 1602–1608. [Google Scholar] [CrossRef]
- Ali, T.; Badshah, H.; Kim, T.H.; Kim, M.O. Melatonin attenuates D-galactose-induced memory impairment, neuroinflammation and neurodegeneration via RAGE/NF-K B/JNK signaling pathway in aging mouse model. J. Pineal Res. 2015, 58, 71–85. [Google Scholar] [CrossRef]
- Zhang, B.; Lian, W.; Zhao, J.; Wang, Z.; Liu, A.; Du, G. DL0410 Alleviates Memory Impairment in D-Galactose-Induced Aging Rats by Suppressing Neuroinflammation via the TLR4/MyD88/NF-κB Pathway. Oxid. Med. Cell. Longev. 2021, 2021, 6521146. [Google Scholar] [CrossRef]
- Cao, N.; Liao, T.; Liu, J.; Fan, Z.; Zeng, Q.; Zhou, J.; Pei, H.; Xi, J.; He, L.; Chen, L.; et al. Clinical-grade human umbilical cord-derived mesenchymal stem cells reverse cognitive aging via improving synaptic plasticity and endogenous neurogenesis. Cell Death Dis. 2017, 8, e2996. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.Q.; Wu, C.; He, X.F.; Wu, D.; He, X.; Liang, F.Y.; Dai, G.Y.; Pei, Z.; Xu, G.Q.; Lan, Y. Effects of Voluntary Wheel-Running Types on Hippocampal Neurogenesis and Spatial Cognition in Middle-Aged Mice. Front. Cell. Neurosci. 2018, 12, 177. [Google Scholar] [CrossRef]
- Laroche, S.; Davis, S.; Jay, T.M. Plasticity at hippocampal to prefrontal cortex synapses: Dual roles in working memory and consolidation. Hippocampus 2000, 10, 438–446. [Google Scholar] [CrossRef] [PubMed]
- Bailey, C.H.; Kandel, E.R.; Harris, K.M. Structural Components of Synaptic Plasticity and Memory Consolidation. Cold Spring Harb. Perspect. Biol. 2015, 7, a021758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weyhersmüller, A.; Hallermann, S.; Wagner, N.; Eilers, J. Rapid active zone remodeling during synaptic plasticity. J. Neurosci. 2011, 31, 6041–6052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Savtchenko, L.P.; Rusakov, D.A. The optimal height of the synaptic cleft. Proc. Natl. Acad. Sci. USA 2007, 104, 1823–1828. [Google Scholar] [CrossRef] [Green Version]
- Thiel, G. Synapsin I, synapsin II, and synaptophysin: Marker proteins of synaptic vesicles. Brain Pathol. 1993, 3, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Sun, M.; Bernard, L.P.; Zhang, H. Postsynaptic density 95 (PSD-95) serine 561 phosphorylation regulates a conformational switch and bidirectional dendritic spine structural plasticity. J. Biol. Chem. 2017, 292, 16150–16160. [Google Scholar] [CrossRef] [Green Version]
- Jeanneret, V.; Ospina, J.P.; Díaz, A.; Manrique, L.G.; Merino, P.; Gutierrez, L.; Torre, E.; Wu, F.; Cheng, L.; Yepes, M. Tissue-type plasminogen activator protects the postsynaptic density in the ischemic brain. J. Cereb. Blood Flow Metab. 2018, 38, 1896–1910. [Google Scholar] [CrossRef]
- Park, H.; Poo, M.M. Neurotrophin regulation of neural circuit development and function. Nat. Rev. Neurosci. 2013, 14, 7–23. [Google Scholar] [CrossRef]
- Cheng, P.L.; Song, A.H.; Wong, Y.H.; Wang, S.; Zhang, X.; Poo, M.-M. Self-amplifying autocrine actions of BDNF in axon development. Proc. Natl. Acad. Sci. USA 2011, 108, 18430–18435. [Google Scholar] [CrossRef] [Green Version]
- Di Benedetto, G.; Iannucci, L.F.; Surdo, N.C.; Zanin, S.; Conca, F.; Grisan, F.; Gerbino, A.; Lefkimmiatis, K. Compartmentalized Signaling in Aging and Neurodegeneration. Cells 2021, 10, 464. [Google Scholar] [CrossRef]
- Blokland, A.; Heckman, P.; Vanmierlo, T.; Schreiber, R.; Paes, D.; Prickaerts, J. Phosphodiesterase Type 4 Inhibition in CNS Diseases. Trends Pharmacol. Sci. 2019, 40, 971–985. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.T.; Huang, Y.; Masood, A.; Stolinski, L.R.; Li, Y.; Zhang, L.; Dlaboga, D.; Jin, S.-L.C.; Conti, M.; O’Donnell, J.M. Anxiogenic-like behavioral phenotype of mice deficient in phosphodiesterase 4B (PDE4B). Neuropsychopharmacology 2008, 33, 1611–1623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmetterer, K.G.; Goldhahn, K.; Ziegler, L.S.; Gerner, M.C.; Schmidt, R.L.; Themanns, M.; Zebedin-Brandl, E.; Trapin, D.; Leitner, J.; Pickl, W.F.; et al. Overexpression of PDE4A Acts as Checkpoint Inhibitor Against cAMP-Mediated Immunosuppression in vitro. Front. Immunol. 2019, 10, 1790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Miguel, Z.; Khoury, N.; Betley, M.J.; Lehallier, B.; Willoughby, D.; Olsson, N.; Yang, A.C.; Hahn, O.; Lu, N.; Vest, R.T.; et al. Exercise plasma boosts memory and dampens brain inflammation via clusterin. Nature 2021, 600, 494–499. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.T.; Huang, Y.; Jin, S.-L.C.; A Frith, S.; Suvarna, N.; Conti, M.; O′donnell, J.M. Antidepressant-like profile and reduced sensitivity to rolipram in mice deficient in the PDE4D phosphodiesterase enzyme. Neuropsychopharmacology 2002, 27, 587–595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Streese, L.; Khan, A.W.; Deiseroth, A.; Hussain, S.; Suades, R.; Tiaden, A.; Kyburz, D.; Cosentino, F.; Hanssen, H. High-intensity interval training modulates retinal microvascular phenotype and DNA methylation of p66Shc gene: A randomized controlled trial (EXAMIN AGE). Eur. Heart J. 2020, 41, 1514–1519. [Google Scholar] [CrossRef] [Green Version]
- Takizawa, T.; Nakashima, K.; Namihira, M.; Ochiai, W.; Uemura, A.; Yanagisawa, M.; Fujita, N.; Nakao, M.; Taga, T. DNA methylation is a critical cell-intrinsic determinant of astrocyte differentiation in the fetal brain. Dev. Cell. 2001, 1, 749–758. [Google Scholar] [CrossRef] [Green Version]
- Fernandes, J.; Arida, R.M.; Gomez-Pinilla, F. Physical exercise as an epigenetic modulator of brain plasticity and cognition. Neurosci. Biobehav. Rev. 2017, 80, 443–456. [Google Scholar] [CrossRef]
- Tomiga, Y.; Sakai, K.; Ra, S.; Kusano, M.; Ito, A.; Uehara, Y.; Takahashi, H.; Kawanaka, K.; Soejima, H.; Higaki, Y. Short-term running exercise alters DNA methylation patterns in neuronal nitric oxide synthase and brain-derived neurotrophic factor genes in the mouse hippocampus and reduces anxiety-like behaviors. Faseb. J. 2021, 35, e21767. [Google Scholar] [CrossRef]
- Brenet, F.; Moh, M.; Funk, P.; Feierstein, E.; Viale, A.J.; Socci, N.D.; Scandura, J. DNA methylation of the first exon is tightly linked to transcriptional silencing. PLoS ONE 2011, 6, e14524. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Forward Primer (5′→3′) | Reverse Primer (5′→3′) |
|---|---|---|
| PDE4A | GGCGTCTCCAACCAGTTCCTAATC | TCTCTTCTTGCAGCAGCTTGAATC |
| PDE4B | GACTGGTACTTCATGCCGCCTTC | ATCAGGTCATCGCCGTGTTGTTC |
| PDE4D | TCTGGCGTCCTCCTCCTTGATAAC | CTTTGTGGGGTTGCTCAGGTCTG |
| GAPDH | GACATGCCGCCTGGAGAAAC | AGCCCAGGATGCCCTTTAGT |
| Gene/region | Sequence 5′→3′ |
|---|---|
| PDE4D promoter 1: site1(40552422), site2(40552449) | PCR primer forward: AGTTTTGATGATTGGGTTTGTAT PCR primer reverse: AACCCCTAAAAACTTATACTTCCTCTT Pyrosequencing primer: GGGTTTGTATTTTTATATATGTTAG |
| PDE4D promoter 2: Site3(40553196) | PCR primer forward: GAGTGGGTGTGTGGATGAGTA PCR primer reverse: AAAAAACCCCCCATCCCAA Pyrosequencing primer: AGTATTTTTGTGGAGGT |
| PDE4D promoter 3: Site4 (40553356) | PCR primer forward: GGGGTGTTGAGTTTTTATAGGAATAAG PCR primer reverse: ACTAAACTACTAACCAAACAAAACACATA Pyrosequencing primer: GGTTATATATGTGGTTGG |
| PDE4D promoter 4: Site5(40553875) | PCR primer forward: GGGAGTATTTTGTTTTTTTTTTTAAGAAGT PCR primer reverse: CCAACAAACAACATTAACATAACACTAT Pyrosequencing primer: CTAACAATCTTACAATTTACCA |
| PDE4D promoter 5: Site6(40553976 ) | PCR primer forward: AGATTGTTAGGGATATTAATTGTAATGAT PCR primer reverse: CCAACAAACAACATTAACATAACACTAT Pyrosequencing primer: ACATAACACTATCTACTACAAAA |
| PDE4D Exon-1 1: Site1(40554193 ) Site2(40554213) Site3(40554217) Site4(40554232) | PCR primer forward: TTGAAGGTAAGAGGAGGAAGTTG PCR primer reverse: AACCCATACTAAAACTCCTATTAATTC Pyrosequencing primer: AGAGGAGGAAGTTGG |
| PDE4D Exon-1 2: Site5(40554278) Site6(40554286) Site7(40554291) Site8(40554308) Site9(40554310) Site10(40554341) Site11(40554360) Site12(40554368) | PCR primer forward: TGTGGAAGGGTTATTAAAGGTATT PCR primer reverse: ATCCAACTTCCTCCTCTTACC Pyrosequencing primer: GGGTTATTAAAGGTATTTATTTAG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, J.; Gao, Q.; Gao, J.; Lv, L.; Liu, R.; Wu, Y.; Li, X.; Jin, Y.; Wang, L. Moderate-Intensity Intermittent Training Alters the DNA Methylation Pattern of PDE4D Gene in Hippocampus to Improve the Ability of Spatial Learning and Memory in Aging Rats Reduced by D-Galactose. Brain Sci. 2023, 13, 422. https://doi.org/10.3390/brainsci13030422
Zhang J, Gao Q, Gao J, Lv L, Liu R, Wu Y, Li X, Jin Y, Wang L. Moderate-Intensity Intermittent Training Alters the DNA Methylation Pattern of PDE4D Gene in Hippocampus to Improve the Ability of Spatial Learning and Memory in Aging Rats Reduced by D-Galactose. Brain Sciences. 2023; 13(3):422. https://doi.org/10.3390/brainsci13030422
Chicago/Turabian StyleZhang, Jinmei, Qiaojing Gao, Jun Gao, Liting Lv, Renfan Liu, Yi Wu, Xue Li, Yu Jin, and Lu Wang. 2023. "Moderate-Intensity Intermittent Training Alters the DNA Methylation Pattern of PDE4D Gene in Hippocampus to Improve the Ability of Spatial Learning and Memory in Aging Rats Reduced by D-Galactose" Brain Sciences 13, no. 3: 422. https://doi.org/10.3390/brainsci13030422