
The Role of the Stimulus in Olfactory Plasticity
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
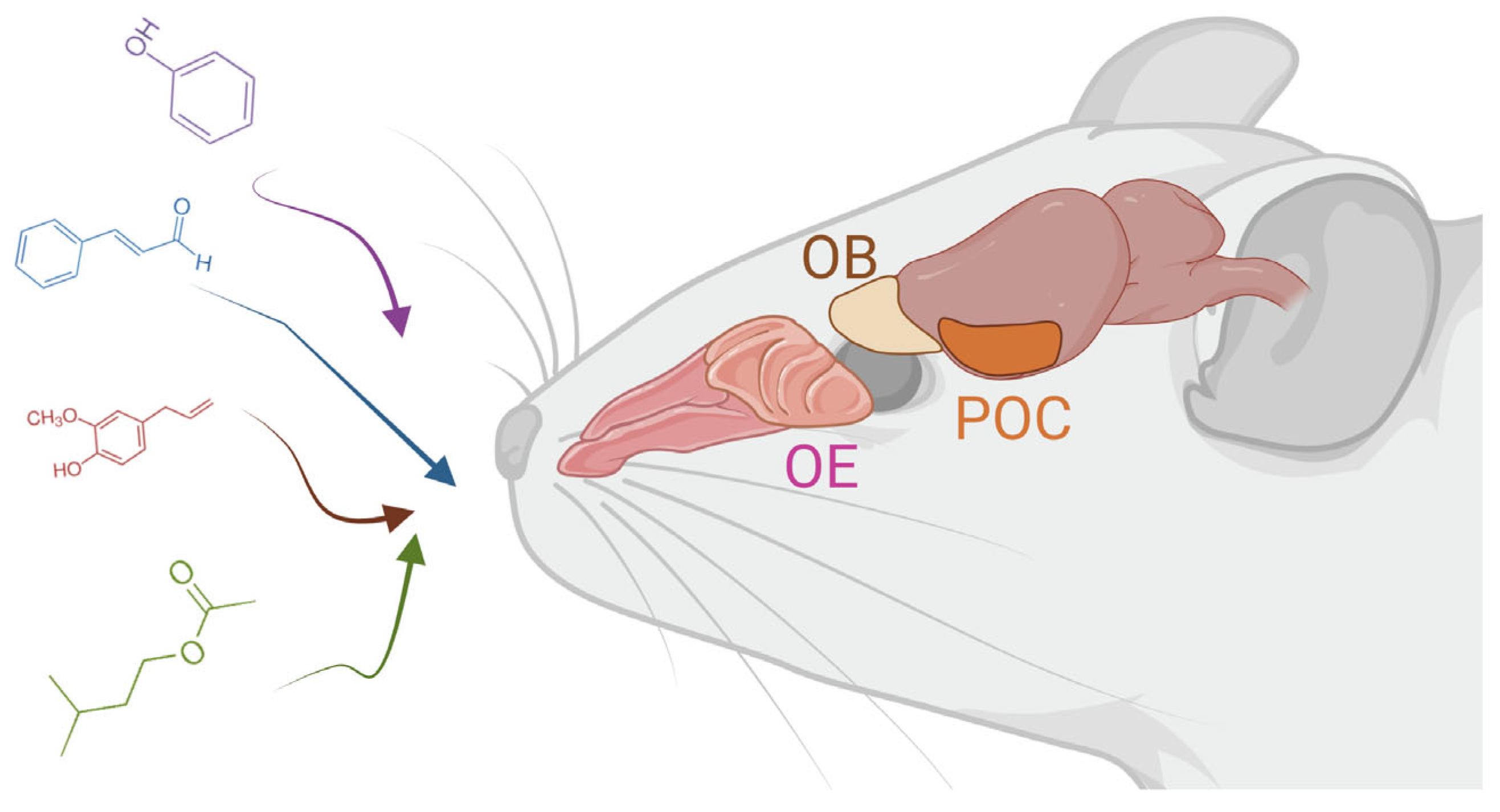
2. The Olfactory System
3. The Effects of Stimulus Deprivation
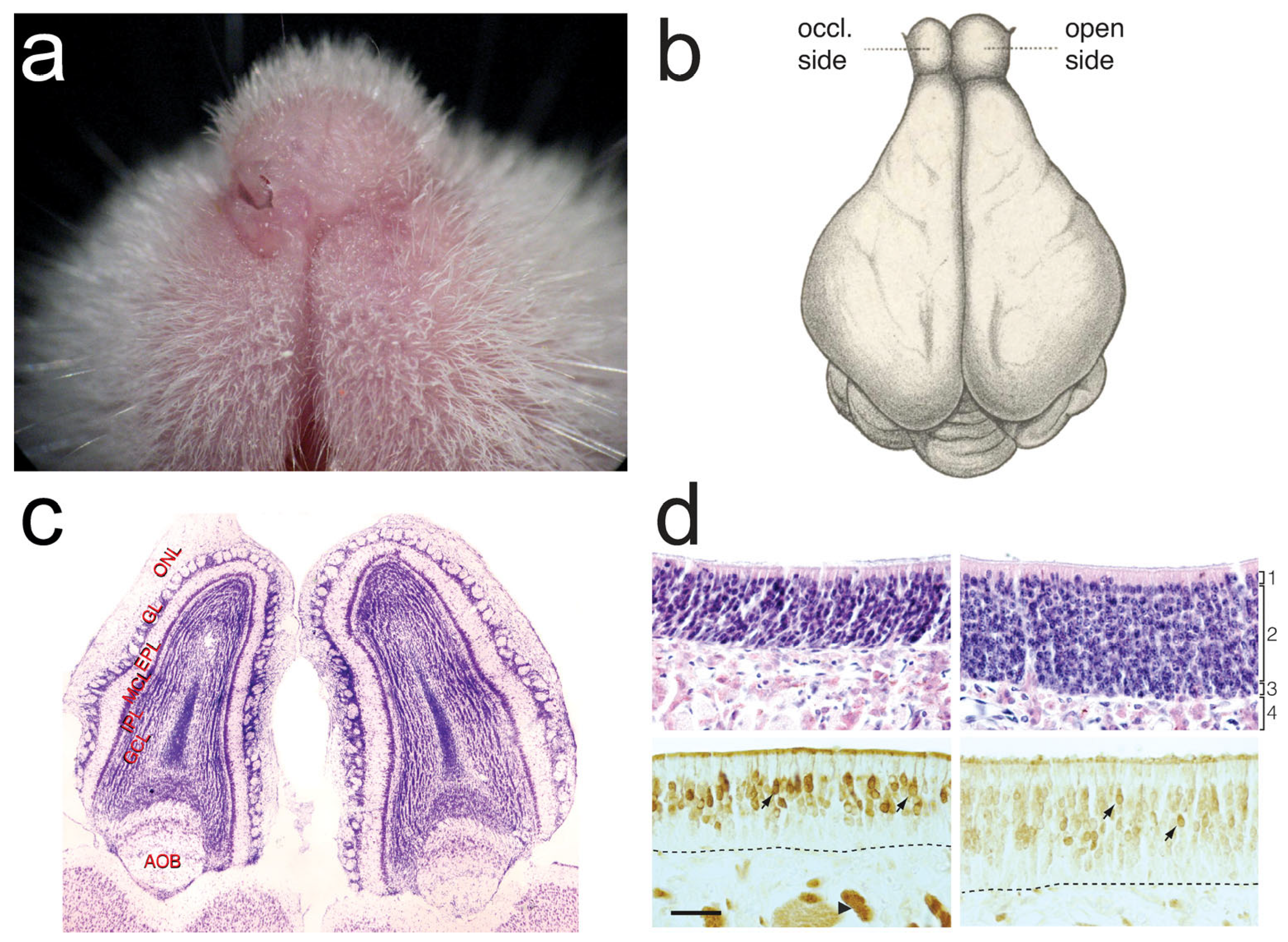
3.1. Deleterious Effects of UNO
3.1.1. Mucosa and OSNs
3.1.2. Olfactory Bulb
3.1.3. Cortex
3.1.4. Neurogenesis
3.1.5. Behavior
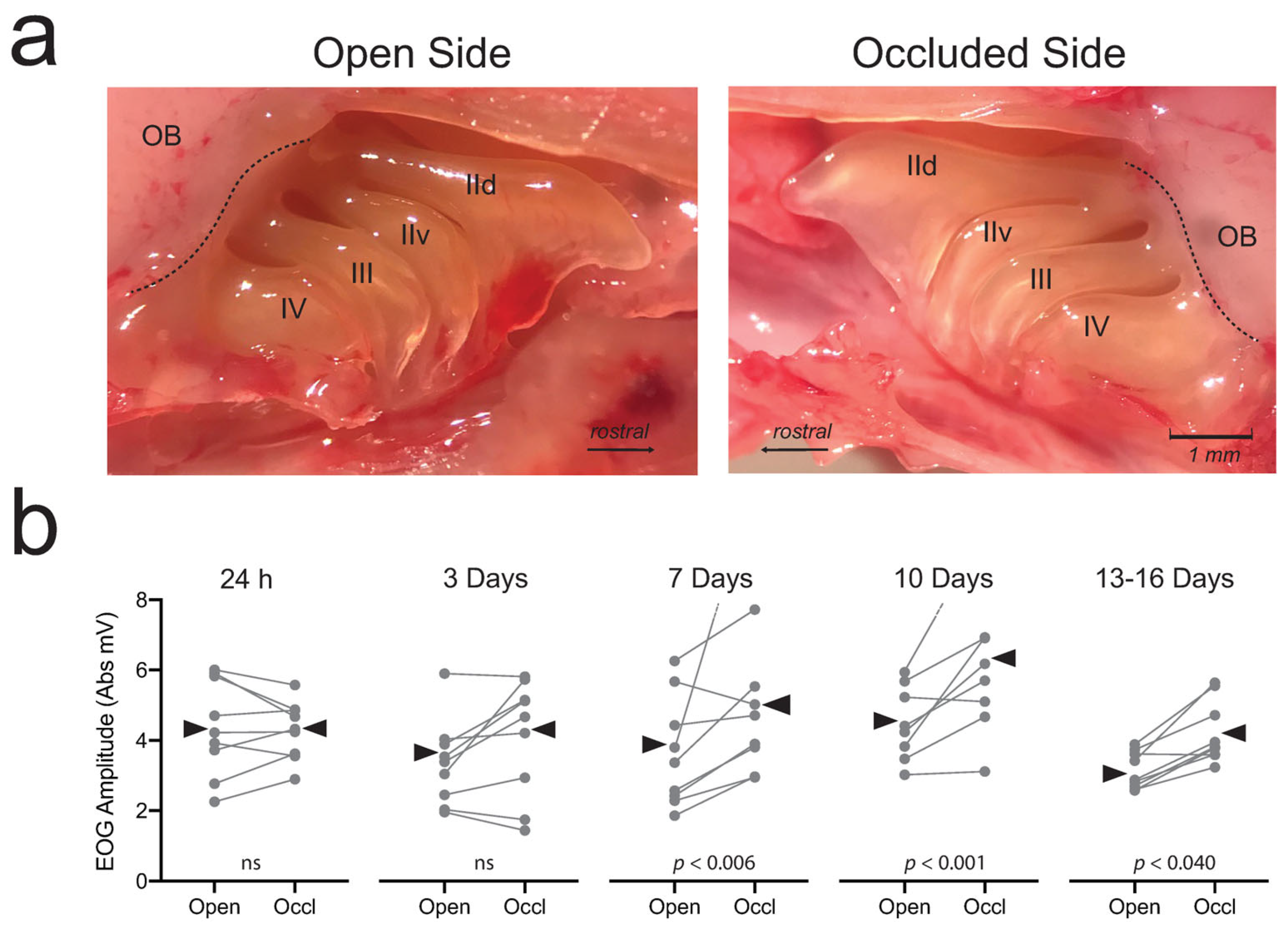
3.2. Compensatory Effects
3.2.1. Mucosa and OSNs
3.2.2. Olfactory Bulb
3.2.3. Olfactory Cortex
3.2.4. Behavior
3.3. Theory
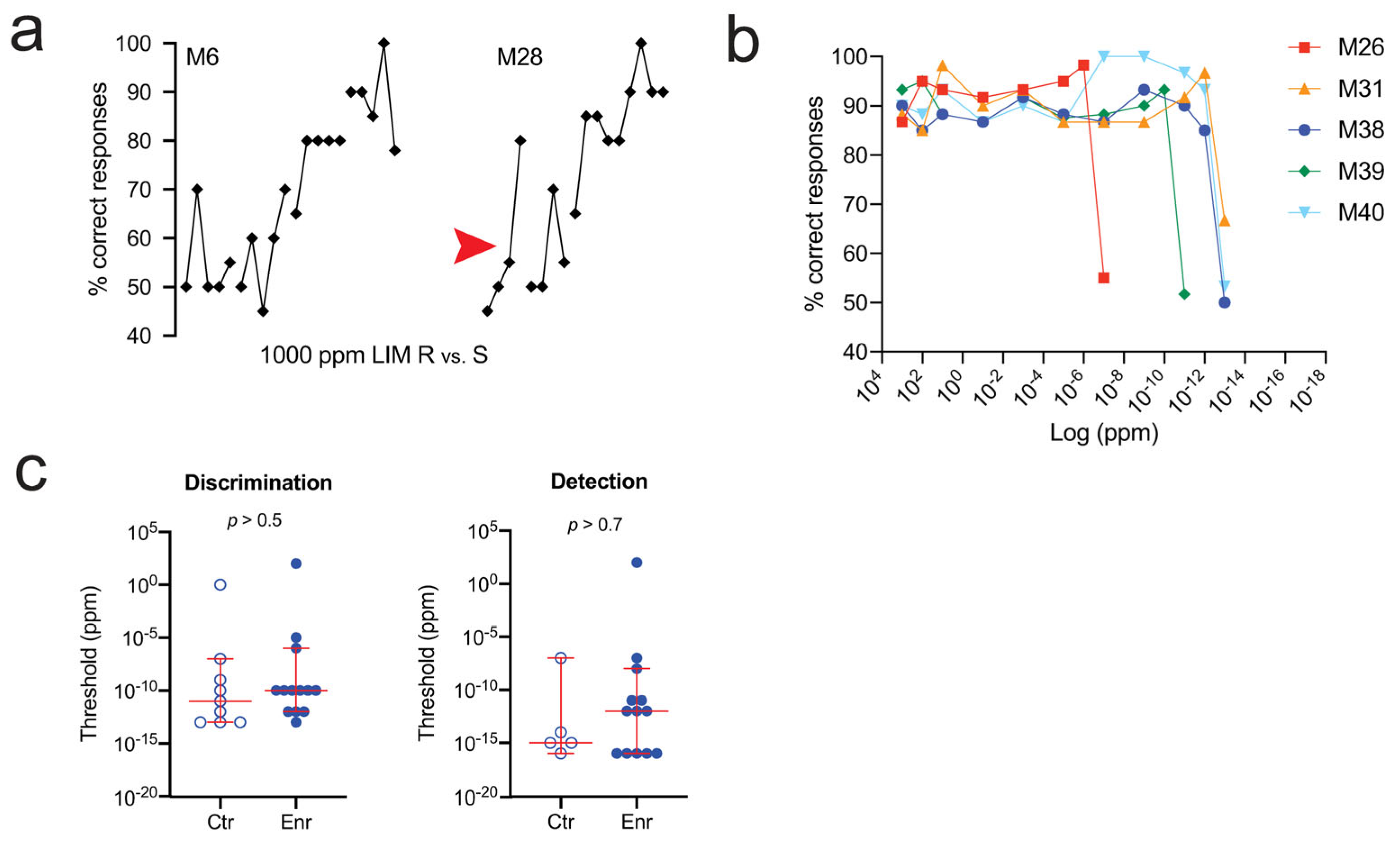
4. Stimulus Enrichment
4.1. Introduction
4.2. Mucosa and OSNs
4.3. Olfactory Bulb
4.4. Olfactory Cortex
4.5. Neurogenesis
4.6. Behavior
4.7. Theory
5. Conclusions
5.1. Overview
5.2. Deprivation
5.3. Enrichment
5.4. Induction
5.5. Neurogenesis
5.6. Aspirations
Author Contributions
Funding
Conflicts of Interest
References
- James, W. The Principles of Psychology; Dover Publications: Mineola, NY, USA, 1950; Volume 1. [Google Scholar]
- Demoor, J. La plasticité morphologique des neurones cérébraux. Arch. Biol. 1896, 14, 723–752. [Google Scholar]
- Zanatta, A.; Cherici, C.; Bargoni, A.; Buzzi, S.; Cani, V.; Mazzarello, P.; Zampieri, F. Vincenzo Malacarne (1744–1816) and the First Description of the Human Cerebellum. Cerebellum 2018, 17, 461–464. [Google Scholar] [CrossRef] [PubMed]
- Gudden, B. Experimentaluntersuchungen über das peripherische und centrale Nervensystem. Arch. Psychiatr. 1870, 2, 693–723. [Google Scholar] [CrossRef]
- Graziadei, P.P.; Monti Graziadei, A.G. Regeneration in the olfactory system of vertebrates. Am. J. Otolaryngol. 1983, 4, 228–233. [Google Scholar] [CrossRef] [PubMed]
- Zou, D.J.; Chesler, A.; Firestein, S. How the olfactory bulb got its glomeruli: A just so story? Nat. Rev. Neurosci. 2009, 10, 611–618. [Google Scholar] [CrossRef] [PubMed]
- Potter, S.M.; Zheng, C.; Koos, D.S.; Feinstein, P.; Fraser, S.E.; Mombaerts, P. Structure and emergence of specific olfactory glomeruli in the mouse. J. Neurosci. 2001, 21, 9713–9723. [Google Scholar] [CrossRef] [PubMed]
- McBride, K.; Slotnick, B.; Margolis, F.L. Does intranasal application of zinc sulfate produce anosmia in the mouse? An olfactometric and anatomical study. Chem. Senses 2003, 28, 659–670. [Google Scholar] [CrossRef]
- Dahlen, J.E.; Jimenez, D.A.; Gerkin, R.C.; Urban, N.N. Morphological analysis of activity-reduced adult-born neurons in the mouse olfactory bulb. Front. Neurosci. 2011, 5, 66. [Google Scholar] [CrossRef]
- Coppola, D.M. Studies of olfactory system neural plasticity: The contribution of the unilateral naris occlusion technique. Neural Plast. 2012, 2012, 351752. [Google Scholar] [CrossRef]
- Coppola, D.M.; White, L.E. Forever young: Neoteny, neurogenesis and a critique of critical periods in olfaction. J. Bioenerg. Biomembr. 2019, 51, 53–63. [Google Scholar] [CrossRef]
- Brunjes, P.C.; Caggiano, A.O.; Korol, D.L.; Stewart, J.S. Unilateral olfactory deprivation: Effects on succinate dehydrogenase histochemistry and [3H]leucine incorporation in the olfactory mucosa. Dev. Brain Res. 1991, 62, 239–244. [Google Scholar] [CrossRef] [PubMed]
- Benson, T.E.; Ryugo, D.K.; Hinds, J.W. Effects of sensory deprivation on the developing mouse olfactory system: A light and electron microscope, morphometric analysis. J. Neurosci. 1984, 4, 638–653. [Google Scholar] [CrossRef]
- Farbman, A.I.; Brunjes, P.C.; Rentfro, L.; Michas, J.; Ritz, S. The effects of unilateral naris occlusion on cell dynamics in the developing rat olfactory epithelium. J. Neurosci. 1988, 8, 3290–3295. [Google Scholar] [CrossRef]
- Stahl, B.; Distel, H.; Hudson, R. Effects of reversible nare occlusion on the development of the olfactory epithelium in the rabbit nasal septum. Cell Tiss. Res. 1990, 259, 275–281. [Google Scholar] [CrossRef] [PubMed]
- Cummings, D.M.; Brunjes, P.C. Changes in cell proliferation in the developing olfactory epithelium following neonatal unilateral naris occlusion. Exp. Neurol. 1994, 128, 124–128. [Google Scholar] [CrossRef] [PubMed]
- Suh, K.S.; Kim, S.Y.; Bae, Y.C.; Ronnett, G.V.; Moon, C. Effects of unilateral naris occlusion on the olfactory epithelium of adult mice. Neuroreport 2006, 17, 1139–1142. [Google Scholar] [CrossRef] [PubMed]
- Mirich, J.M.; Brunjes, P.C. Activity modulates neuronal proliferation in the developing olfactory epithelium. Dev. Brain Res. 2001, 127, 77–80. [Google Scholar] [CrossRef]
- Cummings, D.M.; Belluscio, L. Continuous neural plasticity in the olfactory intrabulbar circuitry. J. Neurosci. 2010, 30, 9172–9180. [Google Scholar] [CrossRef]
- Cavallin, M.A.; Powell, K.; Bijub, K.C.; Fadool, D.A. State-dependent sculpting of olfactory sensory neurons is attributed to sensory enrichment, odor deprivation, and aging. Neurosci. Let. 2010, 483, 90–95. [Google Scholar] [CrossRef]
- Santoro, S.W.; Dulac, C. The activity-dependent histone variant H2BE modulates the life span of olfactory neurons. eLife 2012, 1, e00070. [Google Scholar] [CrossRef]
- Camara, J.A.; Garrosa, M.; Gayoso, M.J. Histological Changes in rat nasal epithelia after unilateral neonatal naris occlusion. Micr. Res. Tech. 2006, 69, 585–594. [Google Scholar] [CrossRef] [PubMed]
- Coppola, D.M.; Craven, B.A.; Seeger, J.; Weiler, E. The effects of naris occlusion on mouse nasal turbinate development. J. Exp. Biol. 2014, 217, 2044–2052. [Google Scholar] [CrossRef] [PubMed]
- Kaneko-Goto, T.; Yoshihara, S.; Miyazaki, H.; Yoshihara, Y. BIG-2 mediates olfactory axon convergence to target glomeruli. Neuron 2008, 57, 834–846. [Google Scholar] [CrossRef]
- Oztokatli, H.; Hörnberg, M.; Berghard, A.; Bohm, S. Retinoic acid receptor and CNGA2 channel signaling are part of a regulatory feedback loop controlling axonal convergence and survival of olfactory sensory neurons. FASEB 2011, 26, 617. [Google Scholar] [CrossRef] [PubMed]
- Williams, E.O.; Sickles, H.M.; Dooley, A.L.; Palumbos, S.; Bisogni, A.J.; Lin, D.M. Delta Protocadherin 10 is Regulated by Activity in the Mouse Main Olfactory System. Front. Neural Circ. 2011, 5, 9. [Google Scholar] [CrossRef] [PubMed]
- Coppola, D.M.; Waggener, C.T. The effects of unilateral naris occlusion on gene expression profiles in mouse olfactory mucosa. J. Mol. Neurosci. 2011, 47, 604–618. [Google Scholar] [CrossRef]
- Ibarra-Soria, X.; Nakahara, T.S.; Lilue, J.; Jiang, Y.; Trimmer, C.; Logan, D.W. Variation in olfactory neuron repertoires is genetically controlled and environmentally modulated. eLife 2017, 6, e21476. [Google Scholar] [CrossRef]
- Tsukahara, T.; Brann, D.H.; Pashkovski, S.L.; Guitchounts, G.; Bozza, T.; Datta, S.R. A transcriptional rheostat couples past activity to future sensory responses. Cell 2021, 184, 6326–6343.e32. [Google Scholar] [CrossRef]
- Horgue, L.F.; Assens, A.; Fodoulian, L.; Marconi, L.; Tuberosa, J.; Haider, A.; Boillat, M.; Carleton, A.; Rodriguez, I. Transcriptional adaptation of olfactory sensory neurons to GPCR identity and activity. Nat. Commun. 2022, 13, 2929. [Google Scholar] [CrossRef]
- Meisami, E. Effects of olfactory deprivation on postnatal growth of the rat olfactory bulb utilizing a new method for production of neonatal unilateral anosmia. Brain Res. 1976, 107, 437–444. [Google Scholar] [CrossRef]
- Meisami, E.; Safari, L. A quantitative study of the effects of early unilateral olfactory deprivation on the number and distribution of mitral and tufted cells and of glomeruli in the rat olfactory bulb. Brain Res. 1981, 221, 81–107. [Google Scholar] [CrossRef] [PubMed]
- Brunjes, P.C. Unilateral naris closure and olfactory system development. Brain Res. Rev. 1994, 19, 146–160. [Google Scholar] [CrossRef] [PubMed]
- Oliva, A.M.; Jones, K.R.; Restrepo, D. Sensory-dependent asymmetry for a urine-responsive olfactory bulb glomerulus. J. Comp. Neurol. 2008, 510, 475–483. [Google Scholar] [CrossRef] [PubMed]
- Oliva, A.M.; Salcedo, E.; Hellier, J.L.; Ly, X.; Koka, K.; Tollin, D.J.; Restrepo, D. Toward a mouse neuroethology in the laboratory environment. PLoS ONE 2010, 5, e11359. [Google Scholar] [CrossRef] [PubMed]
- Inoue, N.; Nishizumi, H.; Ooyama, R.; Mogi, K.; Nishimori, K.; Kikusui, T.; Sakano, H. The olfactory critical period is determined by activity-dependent Sema7A/PlxnC1 signaling within glomeruli. eLife 2021, 10, e65078. [Google Scholar] [CrossRef] [PubMed]
- Frazier, L.L.; Brunjes, P.C. Unilateral odor deprivation: Early postnatal changes in olfactory bulbs cell density and cell number. J. Comp. Neurol. 1988, 269, 355–370. [Google Scholar] [CrossRef] [PubMed]
- Frazier-Cierpial, L.L.; Brunjes, P.C. Early postnatal cellular proliferation and survival in the olfactory bulb and rostral migratory stream of normal and unilaterally odor-deprived rats. J. Comp. Neurol. 1989, 289, 481–492. [Google Scholar] [CrossRef]
- Saghatelyan, A.; Roux, P.; Migliore, M.; Rochefort, C.; Desmaisons, D.; Charneau, P.; Shepherd, G.M.; Lledo, P. Activity-dependent adjustments of the inhibitory network in the olfactory bulb following early postnatal deprivation. Neuron 2005, 46, 103–116. [Google Scholar] [CrossRef]
- Korol, D.L.; Brunjes, P.C. Rapid changes in 2-DG uptake and amino acid incorporation following unilateral odor deprivation: A laminar analysis. Dev. Brain Res. 1990, 52, 75–85. [Google Scholar] [CrossRef]
- Pedersen, P.E.; Shepherd, G.M.; Greer, C.A. Cytochrome oxidase staining in the olfactory epithelium and bulb of normal and odor-deprived neonatal rats. Ann. N. Y. Acad. Sci. 1987, 510, 544–546. [Google Scholar] [CrossRef]
- Meisami, E.; Mousavi, R. Lasting effects of early olfactory deprivation on the growth, DNA, RNA and protein content, and Na-K-ATPase and AChE activity of the rat olfactory bulb. Dev. Brain Res. 1982, 2, 217–229. [Google Scholar] [CrossRef] [PubMed]
- Baker, H. Unilateral neonatal olfactory deprivation alters tyrosine hydroxylase expression but not aromatic amino acid decarboxylase or GABA immunoreactivity. Neuroscience 1990, 36, 761–771. [Google Scholar] [CrossRef] [PubMed]
- Baker, H.; Morell, K.; Stone, D.M.; Maruniak, J.A. Adult naris closure profoundly reduces tyrosine hydroxylase expression in the mouse olfactory bulb. Brain Res. 1993, 614, 109–116. [Google Scholar] [CrossRef] [PubMed]
- Kosaka, T.; Kosaka, K.; Hama, K.; Wu, J.; Nagatsu, I. Differential effect of functional olfactory deprivation on the GABAergic and catecholaminergic traits in the rat main olfactory bulb. Brain Res. 1987, 413, 197–203. [Google Scholar] [CrossRef] [PubMed]
- Baker, H.; Kawano, T.; Margolis, F.L.; Joh, T.H. Transneuronal regulation of tyrosine hydroxylase expression in olfactory bulb of mouse and rat. J. Neurosci. 1983, 3, 69–78. [Google Scholar] [CrossRef] [PubMed]
- Nadi, N.S.; Head, R.; Grillo, M.; Hempstead, J.; Grannot-Reisfeld, N.; Margolis, F.L. Chemical deafferentation of the olfactory bulb: Plasticity of the levels of tyrosine hydroxylase, dopamine and norepinephrine. Brain Res. 1981, 213, 365–377. [Google Scholar] [CrossRef] [PubMed]
- Woo, C.C.; Leon, M. Distribution and development of beta-adrenergic receptors in the rat olfactory bulb. J. Comp. Neurol. 1995, 352, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Brunjes, P.C.; Smith-Crafts, L.K.; McCarty, R. Unilateral odor deprivation: Effects on the development of olfactory bulb catecholamines and behavior. Brain Res. 1985, 354, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, K.A.; Coppola, D.M. Distribution of GluR1 is altered in the olfactory bulb following neonatal naris occlusion. J. Neurobiol. 2003, 54, 326–336. [Google Scholar] [CrossRef]
- Gomez-Pinilla, F.; Guthrie, K.M.; Leon, M.; Nieto-Sampedro, M. NGF receptor increase in the olfactory bulb of the rat after early odor deprivation. Dev. Brain Res. 1989, 48, 161–165. [Google Scholar] [CrossRef] [PubMed]
- McLean, J.H.; Darby-King, A.; Bonnell, W.S. Neonatal olfactory sensory deprivation decreases BDNF in the olfactory bulb of the rat. Dev. Brain Res. 2001, 128, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Biju, K.C.; Mast, T.G.; Fadool, D.A. Olfactory sensory deprivation increases the number of proBDNF-immunoreactive mitral cells in the olfactory bulb of mice. Neurosci. Lett. 2008, 447, 42–47. [Google Scholar] [CrossRef] [PubMed]
- Fadool, D.A.; Tucker, K.; Phillips, J.J.; Simmen, J.A. Brain insulin receptor causes activity-dependent current suppression in the olfactory bulb through multiple phosphorylation of Kv1.3. J. Neurophysiol. 2000, 83, 2332–2348. [Google Scholar] [CrossRef] [PubMed]
- Tucker, K.; Fadool, D.A. Neurotrophin modulation of voltage-gated potassium channels in rat through TrkB receptors is time and sensory experience dependent. J. Physiol. 2002, 542, 413–429. [Google Scholar] [CrossRef] [PubMed]
- Mirich, J.M.; Illig, K.R.; Brunjes, P.C. Experience-dependent activation of extracellular signal-related kinase (ERK) in the olfactory bulb. J. Comp. Neurol. 2004, 479, 234–241. [Google Scholar] [CrossRef] [PubMed]
- Wilson, D.A.; Guthrie, K.M.; Leon, M. Modification of olfactory bulb synaptic inhibition by early unilateral olfactory deprivation. Neurosci. Lett. 1990, 116, 250–256. [Google Scholar] [CrossRef] [PubMed]
- Wilson, D.A.; Wood, J.G. Functional consequences of unilateral olfactory deprivation: Time-course and age sensitivity. Neuroscience 1992, 49, 183–192. [Google Scholar] [CrossRef]
- Wilson, D.A. NMDA receptors mediate expression of one form of functional plasticity induced by olfactory deprivation. Brain Res. 1995, 677, 238–242. [Google Scholar] [CrossRef]
- Philpot, B.D.; Foster, T.C.; Brunjes, P.C. Mitral/tufted cell activity is attenuated and becomes uncoupled from respiration following naris closure. J. Neurobiol. 1997, 33, 374–386. [Google Scholar] [CrossRef]
- Maher, B.J.; McGinley, M.J.; Westbrook, G.L. Experience-dependent maturation of the glomerular microcircuit. Proc. Natl. Acad. Sci. USA 2009, 106, 16865–16870. [Google Scholar] [CrossRef]
- Guthrie, K.M.; Wilson, D.A.; Leon, M. Unilateral olfactory deprivation modifies olfactory bulb function. J. Neurosci. 1990, 10, 3402–3412. [Google Scholar] [CrossRef] [PubMed]
- Yousem, D.M.; Geckle, R.J.; Bilker, W.B.; Kroger, H.; Doty, R.L. Posttraumatic smell loss: Relationship of psychophysical tests and volumes of the olfactory bulbs and tracts and the temporal lobes. Acad. Radiol. 1999, 6, 264–272. [Google Scholar] [CrossRef] [PubMed]
- Buschhüter, D.; Smitka, M.; Puschmann, S.; Gerber, J.C.; Witt, M.; Abolmaali, N.D.; Hummel, T. Correlation between olfactory bulb volume and olfactory function. NeuroImage 2008, 42, 498–502. [Google Scholar] [CrossRef] [PubMed]
- Hummel, T.; Smitka, M.; Puschmann, S.; Gerber, J.C.; Schaal, B.; Buschhüter, D. Correlations between olfactory bulb volume and olfactory function in children and adolescents. Exp. Brain Res. 2011, 214, 285–291. [Google Scholar] [CrossRef]
- Rombaux, P.; Potier, H.; Bertrand, B.; Duprez, T.; Hummel, T. Olfactory bulb volume in patients with sinonasal disease. Am. J. Rhinol. 2008, 22, 598–601. [Google Scholar] [CrossRef] [PubMed]
- Weiss, T.; Soroka, T.; Gorodisky, L.; Shushan, S.; Snitz, K.; Weissgross, R.; Furman-Haran, E.; Dhollander, T.; Sobel, N. Human Olfaction without Apparent Olfactory Bulbs. Neuron 2019, 104, 1023. [Google Scholar] [CrossRef] [PubMed]
- Gudziol, V.; Buschhüter, D.; Abolmaali, N.; Geber, J.; Rombaux, P.; Hummel, T. Increasing olfactory bulb volume due to treatment of chronic rhinosinusitis-a longitudinal study. Brain 2009, 132, 3096–3101. [Google Scholar] [CrossRef] [PubMed]
- Michalakis, S.; Reisert, J.; Geiger, H.; Wetzel, C.; Zong, X.; Bradley, J.; Spehr, M.; Hüttl, S.; Gerstner, A.; Pfeifer, A.; et al. Loss of CNGB1 protein leads to olfactory dysfunction and subciliary cyclic nucleotide-gated channel trapping. J. Biol. Chem. 2006, 281, 35156–35166. [Google Scholar] [CrossRef]
- Wilson, D.A.; Best, A.R.; Brunjes, P.C. Trans-neuronal modification of anterior piriform cortical circuitry in the rat. Brain Res. 2000, 853, 317–322. [Google Scholar] [CrossRef]
- Kim, H.H.; Puche, A.C.; Margolis, F.L. Odorant deprivation reversibly modulates transsynaptic changes in the NR2B-mediated CREB pathway in mouse piriform cortex. J. Neurosci. 2006, 26, 9548–9559. [Google Scholar] [CrossRef]
- Franks, K.M.; Isaacson, J.S. Synapse-specific downregulation of NMDA receptors by early experience: A critical period for plasticity of sensory input to olfactory cortex. Neuron 2005, 47, 101–114. [Google Scholar] [CrossRef] [PubMed]
- Best, A.R.; Wilson, D.A. A postnatal critical period for plasticity of cortical afferents but not cortical association fibers in rat piriform cortex. Brain Res. 2003, 961, 81–87. [Google Scholar] [CrossRef]
- Wu, K.N.; Tan, B.K.; Howard, J.D.; Conley, D.B.; Gottfried, J.A. Olfactory input is critical for sustaining odor quality codes in human orbitofrontal cortex. Nat. Neurosci. 2012, 15, 1313–1319. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.L.; Brunjes, P.C. Development of the anterior olfactory nucleus in normal and unilaterally odor deprived rats. J. Comp. Neurol. 1990, 301, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Gokoffski, K.K.; Kawauchi, S.; Wu, H.H.; Santos, R.; Hollenbeck, P.L.W.; Lander, A.D.; Calof, A.L. Feedback Regulation of Neurogenesis in the Mammalian Olfactory Epithelium: New Insights from Genetics and Systems Biology. In The Neurobiology of Olfaction; Menini, A., Ed.; CRC Press: Boca Raton, FL, USA, 2010; Chapter 10. [Google Scholar]
- Gaun, V.; Martens, J.R.; Schwob, J.E. Lifespan of mature olfactory sensory neurons varies with location in the mouse olfactory epithelium and age of the animal. J. Comp. Neurol. 2022, 530, 2238–2251. [Google Scholar] [CrossRef] [PubMed]
- Petreanu, L.; Alvarez-Buylla, A. Maturation and death of adult-born olfactory bulb granule neurons: Role of olfaction. J. Neurosci. 2002, 22, 6106–6113. [Google Scholar] [CrossRef] [PubMed]
- Mandairon, N.; Sacquet, J.; Jourdan, F.; Didier, A. Long-term fate and distribution of newborn cells in the adult mouse olfactory bulb: Influences of olfactory deprivation. Neuroscience 2006, 141, 443–451. [Google Scholar] [CrossRef]
- Chen, B.; Boesen, A.; Olesch, F.T.; Hummel, T. A life without smell: Olfactory function in people working in odorless rooms. Laryngoscope 2023, in press. [Google Scholar] [CrossRef]
- Coppola, D.M.; Waguespack, A.; Reems, M.; Butman, M.L.; Cherry, J. Naris occlusion alters transductory protein immunoreactivity in olfactory epithelium. Hist. Histopath. 2006, 21, 487–501. [Google Scholar]
- Ottoson, D. Analysis of electrical activity in the olfactory epithelium. Acta Physiol. Scan. Suppl. 1956, 35, 1–83. [Google Scholar]
- Scott, J.W.; Scott-Johnson, P.E. The electroolfactogram: A review of its history and uses. Microsc. Res. Tech. 2002, 58, 152. [Google Scholar] [CrossRef] [PubMed]
- Waggener, C.; Coppola, D.M. Naris occlusion alters the electroolfactogram: Evidence for compensatory plasticity in the olfactory system. Neurosci. Let. 2007, 427, 112–116. [Google Scholar] [CrossRef]
- Barber, C.; Coppola, D.M. Compensatory plasticity in the olfactory epithelium: Age, timing, and reversibility. J. Neurophys. 2015, 114, 2023–2032. [Google Scholar] [CrossRef] [PubMed]
- Fitzwater, E.; Coppola, D.M. Olfactory deprivation and enrichment: An identity of opposites? Chem. Senses 2021, 46, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Tyler, W.J.; Petzold, G.C.; Pal, S.K.; Murthy, V.N. Experience-dependent modification of primary sensory synapses in the mammalian olfactory bulb. J. Neurosci. 2007, 27, 9427–9438. [Google Scholar] [CrossRef] [PubMed]
- Kass, M.D.; Pottackal, J.; Turkel, D.J.; McGann, J.P. Changes in the neural representation of odorants after olfactory deprivation in the adult mouse olfactory bulb. Chem. Senses 2013, 38, 77–89. [Google Scholar] [CrossRef] [PubMed]
- Wilson, D.A.; Sullivan, R.M. The D2 antagonist spiperone mimics the effects of olfactory deprivation on mitral/tufted cell odor response patterns. J. Neurosci. 1995, 15, 5574–5581. [Google Scholar] [CrossRef]
- George, N.M.; Polese, A.G.; Merle, L.; Macklin, W.B.; Restrepo, D. Excitable Axonal Domains Adapt to Sensory Deprivation in the Olfactory System. J. Neurosci. 2022, 42, 1491–1509. [Google Scholar] [CrossRef]
- Guthrie, K.M.; Pullara, J.M.; Marshall, J.F.; Leon, M. Olfactory deprivation increases dompamine D2 receptor density in the rat olfactory bulb. Synapse 1991, 8, 61–70. [Google Scholar] [CrossRef]
- Leon, M. Compensatory responses to early olfactory restriction. Ann. N. Y. Acad. Sci. 1998, 855, 104–108. [Google Scholar] [CrossRef]
- Slotnick, B.M.; Pazos, A.J. Rats with one olfactory bulb removed and the contralateral naris closed can detect odors. Physiol. Behav. 1990, 48, 37–40. [Google Scholar] [CrossRef] [PubMed]
- Hunt, N.L.; Slotnick, B.M. Functional capacity of the rat olfactory bulb after neonatal naris occlusion. Chem. Senses 1991, 16, 131–142. [Google Scholar] [CrossRef]
- Coppola, D.M.; Coltrane, J.A.; Arsov, I. Retronasal or internasal olfaction can mediate odor-guided behaviors in newborn mice. Physiol. Behav. 1994, 56, 729–736. [Google Scholar] [CrossRef] [PubMed]
- Angely, C.; Coppola, D.M. How does long-term odor deprivation affect the olfactory capacity of adult mice? Behav. Brain Funct. 2010, 6, 26. [Google Scholar] [CrossRef]
- Katz, L.C.; Shatz, C.J. Synaptic activity and the construction of cortical circuits. Science 1996, 274, 1133–1138. [Google Scholar] [CrossRef] [PubMed]
- Lodovichi, C. Topographic organization in the olfactory bulb. Cell Tissue Res. 2021, 383, 457–472. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.R.; Power, J.; Barnea, G.; O’Donnell, S.; Brown, H.E.; Osborne, J.; Axel, R.; Gogos, J.A. Spontaneous neural activity is required for the establishment and maintenance of the olfactory sensory map. Neuron 2004, 42, 553–566. [Google Scholar] [CrossRef]
- Daw, N.W. Visual Development; Springer: New York, NY, USA, 2006. [Google Scholar]
- Fox, K.; Wong, R.O.L. A Comparison of experience-dependent plasticity in the visual and somatosensory systems. Neuron 2005, 48, 465–477. [Google Scholar] [CrossRef]
- Liman, E.R.; Buck, L.B. A second subunit of the olfactory cyclic nucleotide-gated channel confers high sensitivity to cAMP. Neuron 1994, 13, 611–621. [Google Scholar] [CrossRef]
- Bradley, J.; Reisert, J.; Frings, S. Regulation of cyclic nucleotide-gated channels. Curr. Opin. Neurobiol. 2005, 15, 343–349. [Google Scholar] [CrossRef]
- Turrigano, G.G.; Nelson, S.B. Homeostatic plasticity in the developing nervous system. Nat. Rev. Neurosci. 2004, 5, 97–107. [Google Scholar] [CrossRef]
- Wysocki, C.J.; Dorries, K.M.; Beauchamp, G.K. Ability to perceive androstenone can be acquired by ostensibly anosmic people. Proc. Natl. Acad. Sci. USA 1989, 86, 7976–7978. [Google Scholar] [CrossRef] [PubMed]
- Wysocki, C.J.; Whitney, G.; Tucker, D. Specific anosmia in the laboratory mouse. Behav. Genet. 1977, 7, 171–188. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.W.; Wysocki, C.J.; Gold, G.H. Induction of olfactory receptor sensitivity in mice. Science 1993, 260, 998–1000. [Google Scholar] [CrossRef] [PubMed]
- Yee, K.K.; Wysocki, C.J. Odorant exposure increases olfactory sensitivity: Olfactory epithelium is implicated. Physiol. Behav. 2001, 72, 705–711. [Google Scholar] [CrossRef] [PubMed]
- Mainland, J.D.; Bremner, E.A.; Young, N.; Johnson, B.; Khan, B.M.; Bensafi, M.; Sobel, N. One nostril knows what the other learns. Nature 2002, 49, 802. [Google Scholar] [CrossRef] [PubMed]
- Hudson, R.; Distel, H. Induced peripheral sensitivity in the developing vertebrate olfactory system. Ann. N. Y. Acad. Sci. 1998, 855, 109–115. [Google Scholar] [CrossRef] [PubMed]
- Hudson, R. From molecule to mind, the role of experience in shaping olfactory function. J. Comp. Physiol. A 1999, 185, 297–304. [Google Scholar] [CrossRef]
- Duchamp-Viret, P.; Boyer, J.; La Villa, F.; Coureaud, G. Brief olfactory learning drives perceptive sensitivity in newborn rabbits: New insights in peripheral processing of odor mixtures and induction. Physiol. Behav. 2021, 229, 113217. [Google Scholar] [CrossRef]
- Watt, W.C.; Sakano, H.; Lee, Z.Y.; Reusch, J.E.; Trinh, K.; Storm, D.R. Odorant stimulation enhances survival of olfactory sensory neurons via MAPK and CREB. Neuron 2004, 41, 955–967. [Google Scholar] [CrossRef]
- Dewaele, A.; Persuy, M.A.; Badonnel, K.; Meunier, N.; Durieux, D.; Castille, J.; Favreau-Peigne, A.; Baly, C. Chronic perinatal odour exposure with heptaldehyde affects odour sensitivity and olfactory system homeostasis in preweaning mice. Behav. Brain Res. 2018, 347, 414–424. [Google Scholar] [CrossRef] [PubMed]
- Barbour, J.; Neuhaus, E.M.; Piechura, H.; Stoepel, N.; Mashukova, A.; Brunert, D.; Sitek, B.; Stühler, K.; Meyer, H.E.; Hatt, H.; et al. New insight into stimulus-induced plasticity of the olfactory epithelium in Mus musculus by quantitative proteomics. J. Proteome Res. 2008, 7, 1594–1605. [Google Scholar] [CrossRef] [PubMed]
- Cadiou, H.; Aoude, I.; Tazir, B.; Molinas, A.; Fenech, C.; Meunier, N.; Grosmaitre, X. Postnatal odorant exposure induces peripheral olfactory plasticity at the cellular level. J. Neurosci. 2014, 34, 4857–4870. [Google Scholar] [CrossRef] [PubMed]
- Monjaraz-Fuentes, F.; Millan-Adalco, D.; Palomero-Rivero, M.; Hudson, R.; Drucker-Colin, R. Recovery of glomerular morphology in the olfactory bulb of young mice after disruption caused by continuous odorant exposure. Brain Res. 2017, 1670, 6–13. [Google Scholar] [CrossRef] [PubMed]
- Jones, S.V.; Choi, D.C.; Davis, M.; Ressler, K.J. Learning-dependent structural plasticity in the adult olfactory pathway. J. Neurosci. 2008, 28, 13106–13111. [Google Scholar] [CrossRef] [PubMed]
- Morrison, F.G.; Dias, B.G.; Ressler, K.J. Extinction reverses olfactory fear-conditioned increases in neuron number and glomerular size. Proc. Natl. Acad. Sci. USA 2015, 112, 12846–12851. [Google Scholar] [CrossRef] [PubMed]
- Dias, B.G.; Ressler, K.J. Parental olfactory experience influences behavior and neural structure in subsequent generations. Nat. Neurosci. 2014, 17, 89–96. [Google Scholar] [CrossRef]
- Von der Weid, B.; Rossier, D.; Lindup, M.; Tuberosa, J.; Widmer, A.; Col, J.D.; Kan, C.; Carleton, A.; Rodriguez, I. Large-scale transcriptional profiling of chemosensory neurons identifies receptor-ligand pairs in vivo. Nat. Neurosci. 2015, 18, 1455–1463. [Google Scholar] [CrossRef]
- Doving, K.B.; Pinching, A.J. Selective degeneration of neurons in the olfactory bulb following prolonged odour exposure. Brain Res. 1973, 52, 115–129. [Google Scholar] [CrossRef]
- Laing, D.G.; Panhuber, H.; Pittman, E.A.; Willcox, M.E.; Eagleson, G.K. Prolonged exposure to an odor or deodorized air alters the size of mitral cells in the olfactory bulb. Brain Res. 1985, 336, 81–87. [Google Scholar] [CrossRef]
- Panhuber, H.; Laing, D.G. The size of mitral cells is altered when rats are exposed to an odor from their day of birth. Brain Res. 1987, 431, 133–140. [Google Scholar] [CrossRef]
- Rosselli-Austin, L.; Williams, J. Enriched neonatal odor exposure leads to increased numbers of olfactory bulb mitral and granule cells, Brain Res. Dev. Brain Res. 1990, 51, 135–137. [Google Scholar] [CrossRef] [PubMed]
- Johnson, M.C.; Biju, K.C.; Hoffman, J.; Fadool, D.A. Odor enrichment sculpts the abundance of olfactory bulb mitral cells. Neurosci. Lett. 2013, 541, 173–178. [Google Scholar] [CrossRef]
- Kerr, M.A.; Belluscio, L. Olfactory experience accelerates glomerular refinement in the mammalian olfactory bulb. Nat. Neurosci. 2006, 9, 484–486. [Google Scholar] [CrossRef] [PubMed]
- Todrank, J.; Heth, G.; Restrepo, D. Effects of in utero odorant exposure on neuroanatomical development of the olfactory bulb and odour preferences. Proc. Biol. Sci. 2011, 278, 1949–1955. [Google Scholar] [CrossRef] [PubMed]
- Valle-Leija, P.; Blanco-Hernandez, E.; Drucker-Colin, R.; Gutierrez-Ospina, G.; Vidaltamayo, R. Supernumerary formation of olfactory glomeruli induced by chronic odorant exposure: A constructivist expression of neural plasticity. PLoS ONE 2012, 7, e35358. [Google Scholar] [CrossRef] [PubMed]
- Kass, M.D.; Moberly, A.H.; Rosenthal, M.C.; Guang, S.A.; McGann, J.P. Odor-specific, olfactory marker protein-mediated sparsening of primary olfactory input to the brain after odor exposure. J. Neurosci. 2013, 33, 6594–6602. [Google Scholar] [CrossRef]
- Kass, M.D.; Guang, S.A.; Moberly, A.H.; McGann, J.P. Changes in olfactory sensory neuron physiology and olfactory perceptual learning after odorant exposure in adult mice. Chem. Senses 2016, 41, 123–133. [Google Scholar] [CrossRef]
- Abraham, N.M.; Vincis, R.; Lagier, S.; Rodriguez, I.; Carleton, A. Long term functional plasticity of sensory inputs mediated by olfactory learning. eLife 2014, 3, e02109. [Google Scholar] [CrossRef]
- Hebb, D.O. The effects of early experience on problem solving at maturity. Am. Psychol. 1947, 2, 306–307. [Google Scholar]
- Krech, D.; Rosenzweig, M.R.; Bennett, E.L. Effects of environmental complexity and training on brain chemistry. J. Comp. Physiol. Psychol. 1960, 53, 509–519. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Soto, R.; Pimentel-Farfan, A.K.; Adan-Castro, E.; Clapp, C.; Peña-Ortega, F. Main olfactory bulb reconfiguration by prolonged passive olfactory experience correlates with increased brain-derived neurotrophic factor and improved innate olfaction. Eur. J. Neurosci. 2022, 55, 1141–1161. [Google Scholar]
- Rochefort, C.; Gheusi, G.; Vincent, J.D.; Lledo, P.M. Enriched odor exposure increases the number of newborn neurons in the adult olfactory bulb and improves odor memory. J. Neurosci. 2002, 22, 2679–2689. [Google Scholar] [CrossRef] [PubMed]
- Chan, W.; Singh, S.; Keshav, T.; Dewan, R.; Eberly, C.; Maurer, R.; Nunez-Parra, A.; Araneda, R.C. Mice Lacking M1 and M3 Muscarinic Acetylcholine Receptors Have Impaired Odor Discrimination and Learning. Front. Synaptic Neurosci. 2017, 9, 4. [Google Scholar] [CrossRef] [PubMed]
- Chu, M.W.; Li, W.L.; Komiyama, T. Balancing the Robustness and Efficiency of Odor Representations during Learning. Neuron 2016, 92, 174–186. [Google Scholar] [CrossRef]
- Liu, A.; Urban, N.N. Prenatal and early postnatal odorant exposure heightens odor-evoked Mitral cell responses in the mouse olfactory bulb. eNeuro 2017, 4, ENEURO.0129-17.2017. [Google Scholar] [CrossRef] [PubMed]
- Haberly, L.B.; Bower, J.M. Olfactory cortex: Model circuit for study of associative memory? Trends Neurosci. 1989, 12, 258–264. [Google Scholar] [CrossRef]
- Wilson, D.A. Habituation of odor responses in the rat anterior piriform cortex. J. Neurophysiol. 1998, 79, 1425–1440. [Google Scholar] [CrossRef]
- Wilson, D.A. Rapid, experience-induced enhancement in odorant discrimination by anterior piriform cortex neurons. J. Neurophysiol. 2003, 90, 65–72. [Google Scholar] [CrossRef]
- Barnes, D.C.; Hofacer, R.D.; Zaman, A.R.; Rennaker, R.L.; Wilson, D.A. Olfactory perceptual stability and discrimination. Nat. Neurosci. 2008, 11, 1378–1380. [Google Scholar] [CrossRef]
- Chapuis, J.; Wilson, D.A. Bidirectional plasticity of cortical pattern recognition and behavioral sensory acuity. Nat. Neurosci. 2011, 15, 155–161. [Google Scholar] [CrossRef]
- Wilson, D.A. Single-unit activity in piriform cortex during slow-wave state is shaped by recent odor experience. J. Neurosci. 2010, 30, 1760–1765. [Google Scholar] [CrossRef] [PubMed]
- Shakhawat, A.M.; Harley, C.W.; Yuan, Q. Arc visualization of odor objects reveals experience-dependent ensemble sharpening, separation, and merging in anterior piriform cortex in adult rat. J. Neurosci. 2014, 34, 10206–10210. [Google Scholar] [CrossRef] [PubMed]
- Pashkovski, S.L.; Iurilli, G.; Brann, D.; Chicharro, D.; Drummey, K.; Franks, K.M.; Panzeri, S.; Datta, S.R. Structure and flexibility in cortical representations of odour space. Nature 2020, 583, 253–258. [Google Scholar] [CrossRef] [PubMed]
- Lledo, P.M.; Saghatelyan, A. Integrating new neurons into the adult olfactory bulb: Joining the network, life-death decisions, and the effects of sensory experience. Trends Neurosci. 2005, 28, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Lazarini, F.; Lledo, P.M. Is adult neurogenesis essential for olfaction? Trends Neurosci. 2011, 34, 20–30. [Google Scholar] [CrossRef] [PubMed]
- Alonso, M.; Lepousez, G.; Sebastien, W.; Bardy, C.; Gabellec, M.M.; Torquet, N.; Lledo, P.M. Activation of adult-born neurons facilitates learning and memory. Nat. Neurosci. 2012, 15, 897–904. [Google Scholar] [CrossRef] [PubMed]
- Avaro, V.; Hummel, T.; Calegari, F. Scent of stem cells: How can neurogenesis make us smell better? Front. Neurosci. 2022, 16, 964395. [Google Scholar] [CrossRef]
- Engen, T. Effect of practice and instruction on olfactory thresholds. Percept. Mot. Skills 1960, 10, 195–198. [Google Scholar] [CrossRef]
- Rabin, M.D.; Cain, W.S. Determinants of measured olfactory sensitivity. Percept. Psychophys. 1986, 39, 281–286. [Google Scholar] [CrossRef]
- Rabin, M.D. Experience facilitates olfactory quality discrimination. Percept. Psychophys. 1988, 44, 532–540. [Google Scholar] [CrossRef] [PubMed]
- Poletti, S.C.; Michel, E.; Hummel, T. Olfactory training using heavy and light weight molecule odors. Perception 2017, 46, 343–351. [Google Scholar] [CrossRef] [PubMed]
- Cunzeman, P.J.; Slotnick, B.M. Prolonged exposure to odors in the rat: Effects on odor detection and on mitral cells. Chem. Senses 1984, 9, 229–239. [Google Scholar] [CrossRef]
- Blount, A.; Coppola, D.M. The effect of odor enrichment on olfactory acuity: Olfactometric testing in mice using two mirror-molecular pairs. PLoS ONE 2020, 15, e0233250. [Google Scholar] [CrossRef] [PubMed]
- Voznessenskaya, V.; Parfyonova, V.M.; Wysocki, C.J. Induced olfactory sensitivity in rodents: A general phenomenon. Adv. Bio. Sci. 1995, 93, 399–406. [Google Scholar]
- Voznessenskaya, V.V.; Wysocki, C.J.; Chukhrai, E.S.; Poltorack, O.M.; Atyaksheva, L.F. Long-lasting effects of chemical exposure in mice. In Advance in Chemical Signals in Vertebrates; Johnston, R.E., Muller-Schwarze, D., Sorensen, P.W., Eds.; Kluwer Academic/Plenum Publishers: New York, NY, USA, 1999. [Google Scholar]
- Smotherman, W.P. In utero chemosensory experience alters taste preferences and corticosterone responsiveness. Behav. Neural. Biol. 1982, 36, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Nolte, D.L.; Mason, J.R. Maternal ingestion of orthoaminoacetophenone during gestation affects intake by offspring. Physiol. Behav. 1995, 58, 925–928. [Google Scholar] [CrossRef] [PubMed]
- Fletcher, M.L.; Wilson, D.A. Experience modifies olfactory acuity: Acetylcholine-dependent learning decreases behavioral generalization between similar odorants. J. Neurosci. 2002, 22, RC201. [Google Scholar] [CrossRef]
- Mandairon, N.; Stack, C.; Linster, C. Olfactory enrichment improves the recognition of individual components in mixtures. Physiol. Behav. 2006, 89, 379–384. [Google Scholar] [CrossRef]
- Mandairon, N.; Stack, C.; Kiselycznyk, C.; Linster, C. Enrichment to odors improves olfactory discrimination in adult rats. Behav. Neurosci. 2006, 120, 173–179. [Google Scholar] [CrossRef]
- Vinera, J.; Kermen, F.; Sacquet, J.; Didier, A.; Mandairon, N.; Richard, M. Olfactory perceptual learning requires action of noradrenaline in the olfactory bulb: Comparison with olfactory associative learning. Learn. Mem. 2015, 22, 192–196. [Google Scholar] [CrossRef] [PubMed]
- Escanilla, O.; Mandairon, N.; Linster, C. Odor-reward learning and enrichment have similar effects on odor perception. Physiol. Behav. 2008, 94, 621–626. [Google Scholar] [CrossRef] [PubMed]
- Fleming, G.; Wright, B.A.; Wilson, D.A. The Value of Homework: Exposure to Odors in the Home Cage Enhances Odor-Discrimination Learning in Mice. Chem. Senses 2019, 44, 135–143. [Google Scholar] [CrossRef] [PubMed]
- Hall, N.J.; Smith, D.W.; Wynne, C.D. Effect of odor preexposure on acquisition of an odor discrimination in dogs. Learn. Behav. 2014, 42, 144–152. [Google Scholar] [CrossRef] [PubMed]
- Konstantinidis, I.; Tsakiropoulou, E.; Bekiaridou, P.; Kazantzidou, C.; Constantinidis, J. Use of olfactory training in post-traumatic and postinfectious olfactory dysfunction. Laryngoscope 2013, 123, E85–E90. [Google Scholar] [CrossRef] [PubMed]
- Damm, M.; Pikart, L.K.; Reimann, H.; Burkert, S.; Göktas, Ö.; Haxel, B.; Hüttenbrink, K.B. Olfactory training is helpful in postinfectious olfactory loss: A randomized, controlled, multicenter study. Laryngoscope 2014, 124, 826–831. [Google Scholar] [CrossRef] [PubMed]
- Sorokowska, A.; Drechsler, E.; Karwowski, M.; Hummel, T. Effects of olfactory training: A meta-analysis. Rhinology 2017, 55, 17–26. [Google Scholar] [CrossRef]
- Huang, T.; Wei, Y.; Wu, D. Effects of olfactory training on posttraumatic olfactory dysfunction: A systematic review and meta-analysis. Int. Forum Allergy Rhinol. 2021, 11, 1102–1112. [Google Scholar] [CrossRef]
- Zambom-Ferraresi, F.; Fernández-Irigoyen, J.; Lachén-Montes, M.; Cartas-Cejudo, P.; Lasarte, J.J.; Casares, N.; Fernández, S.; Cedeño-Veloz, B.A.; Maraví-Aznar, E.; Uzcanga-Lacabe, M.I.; et al. Olfactory characterization and training in older adults: Protocol study. Front. Aging Neurosci. 2021, 13, 757081. [Google Scholar] [CrossRef]
- Haehner, A.; Tosch, C.; Wolz, M.; Klingelhoefer, L.; Fauser, M.; Storch, A.; Reichmann, H.; Hummel, T. Olfactory training in patients with Parkinson’s disease. PLoS ONE 2013, 8, e61680. [Google Scholar] [CrossRef]
- Woo, C.C.; Miranda, B.; Sathishkumar, M.; Dehkordi-Vakil, F.; Yassa, M.A.; Leon, M. Overnight olfactory enrichment using an odorant diffuser improves memory and modifies the uncinate fasciculus in older adults. Front. Neurosci. 2023, 17, 1200448. [Google Scholar] [CrossRef] [PubMed]
- Altman, J.; Das, G.D. Autoradiographic examination of the effects of enriched environment on the rate of glial multiplication in the adult rat brain. Nature 1964, 204, 1161–1163. [Google Scholar] [CrossRef] [PubMed]
- Diamond, M.C.; Krech, D.; Rosenzweig, M.R. The effects of an enriched environment on the histology of the rat cerebral cortex. J. Comp. Neurol. 1964, 123, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Gibson, E.J. Principles of Perceptual Learning and Development; Appleton-Century-Crofts: East Norwalk, CT, USA, 1969. [Google Scholar]
- Field, D.J. What is the goal of sensory coding? Neural Comp. 1994, 6, 559–601. [Google Scholar] [CrossRef]
- Thompson, R.F.; Spencer, W.A. Habituation: A model phenomenon for the study of neuronal substrates of behavior. Psychol. Rev. 1966, 73, 16–43. [Google Scholar] [CrossRef] [PubMed]
- Slotnick, B.; Coppola, D.M. Odor-cued taste avoidance: A simple and robust test of mouse olfaction. Chem. Senses 2015, 40, 269–278. [Google Scholar] [CrossRef] [PubMed]
- Semke, E.; Distel, H.; Hudson, R. Specific enhancement of olfactory receptor sensitivity associated with foetal learning of food odors in the rabbit. Naturwissenschaften 1995, 82, 148–149. [Google Scholar] [CrossRef] [PubMed]
- Moreno, M.M.; Linster, C.; Escanilla, O.; Sacquet, J.; Didier, A.; Mandairon, N. Olfactory perceptual learning requires adult neurogenesis. Proc. Natl. Acad. Sci. USA 2009, 106, 17980–17985. [Google Scholar] [CrossRef]
- Sailor, K.A.; Schinder, A.F.; Lledo, P.M. Adult neurogenesis beyond the niche: Its potential for driving brain plasticity. Curr. Opin. Neurobiol. 2017, 42, 111–117. [Google Scholar] [CrossRef]
- Khan, M.; Vaes, E.; Mombaerts, P. Temporal patterns of odorant receptor gene expression in adult and aged mice. Mol. Cell. Neurosci. 2013, 57, 120–129. [Google Scholar] [CrossRef]
- Furudono, Y.; Sone, Y.; Takizawa, K.; Hirono, J.; Sato, T. Relationship between peripheral receptor code and perceived odor quality. Chem. Senses 2009, 34, 151–158. [Google Scholar] [CrossRef]
- Enwere, E.; Shingo, T.; Gregg, C.; Fujikawa, H.; Ohta, S.; Weiss, S. Aging results in reduced epidermal growth factor receptor signaling, diminished olfactory neurogenesis, and deficits in fine olfactory discrimination. J. Neurosci. 2004, 24, 8354–8365. [Google Scholar] [CrossRef] [PubMed]
- Breton-Provencher, V.; Lemasson, M.; Peralta, M.R.; Saghatelyan, A. Interneurons produced in adulthood are required for the normal functioning of the olfactory bulb network and for the execution of selected olfactory behaviors. J. Neurosci. 2009, 29, 15245–15257. [Google Scholar] [CrossRef] [PubMed]
- Sakamoto, M.; Imayoshi, I.; Ohtsuka, T.; Yamaguchi, M.; Mori, K.; Kageyama, R. Continuous neurogenesis in the adult forebrain is required for innate olfactory responses. Proc. Natl. Acad. Sci. USA 2011, 108, 8479–8484. [Google Scholar] [CrossRef] [PubMed]
- Grelat, A.; Benoit, L.; Wagner, S.; Moigneu, C.; Lledo, P.M.; Alonso, M. Adult-born neurons boost odor-reward association. Proc. Natl. Acad. Sci. USA 2018, 115, 2514–2519. [Google Scholar] [CrossRef] [PubMed]
- Sanai, N.; Nguyen, T.; Ihrie, R.A.; Mirzadeh, Z.; Tsai, H.H.; Wong, M.; Gupta, N.; Berger, M.S.; Huang, E.; Garcia-Verdugo, J.M.; et al. Corridors of migrating neurons in the human brain and their decline during infancy. Nature 2011, 478, 382–386. [Google Scholar] [CrossRef] [PubMed]
- Laska, M. Human and animal olfactory capabilities compared. In Handbook of Odor; Buettner, A., Ed.; Springer: New York, NY, USA, 2017; pp. 675–689. [Google Scholar]
- Kaslin, J.; Ganz, J.; Brand, M. Proliferation, neurogenesis and regeneration in the non-mammalian vertebrate brain. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2008, 363, 101–122. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Coppola, D.M.; Reisert, J. The Role of the Stimulus in Olfactory Plasticity. Brain Sci. 2023, 13, 1553. https://doi.org/10.3390/brainsci13111553
Coppola DM, Reisert J. The Role of the Stimulus in Olfactory Plasticity. Brain Sciences. 2023; 13(11):1553. https://doi.org/10.3390/brainsci13111553
Chicago/Turabian StyleCoppola, David M., and Johannes Reisert. 2023. "The Role of the Stimulus in Olfactory Plasticity" Brain Sciences 13, no. 11: 1553. https://doi.org/10.3390/brainsci13111553