
Acute or Chronic Exposure to Corticosterone Promotes Wakefulness in Mice

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. CORT Exposure
2.3. Plasma CORT Measurements
2.4. Surgery
2.5. EEG/EMG Recordings and Analysis
2.6. OF Test
2.7. SPT
2.8. EPM
2.9. TST
2.10. NSF Test
2.11. Self-Grooming Counts
2.12. Immunohistochemistry
2.13. Cell Counting
2.14. Statistical Analysis
3. Results
3.1. Acute CORT Administration Induced a Rapid Increase in Wakefulness and a Decrease in REM and NREM Sleep Accompanied by the Altered EEG Delta Power
3.2. Chronic CORT Administration Elevated CORT Levels and Induced Depressive-like Behaviour in Mice
3.3. Chronic CORT Administration Promoted Wakefulness and REM Sleep While It Suppressed NREM Sleep
3.4. Changes in Wakefulness, REM Sleep, NREM Sleep, and REM Sleep Ratio Induced by Chronic CORT Exposure Predominantly Occurred during the Light Phase
3.5. Chronic CORT Administration Increased Episodes of Wakefulness, REM Sleep, and NREM Sleep, Prolonged REM Sleep Duration and Shortened NREM Sleep Duration
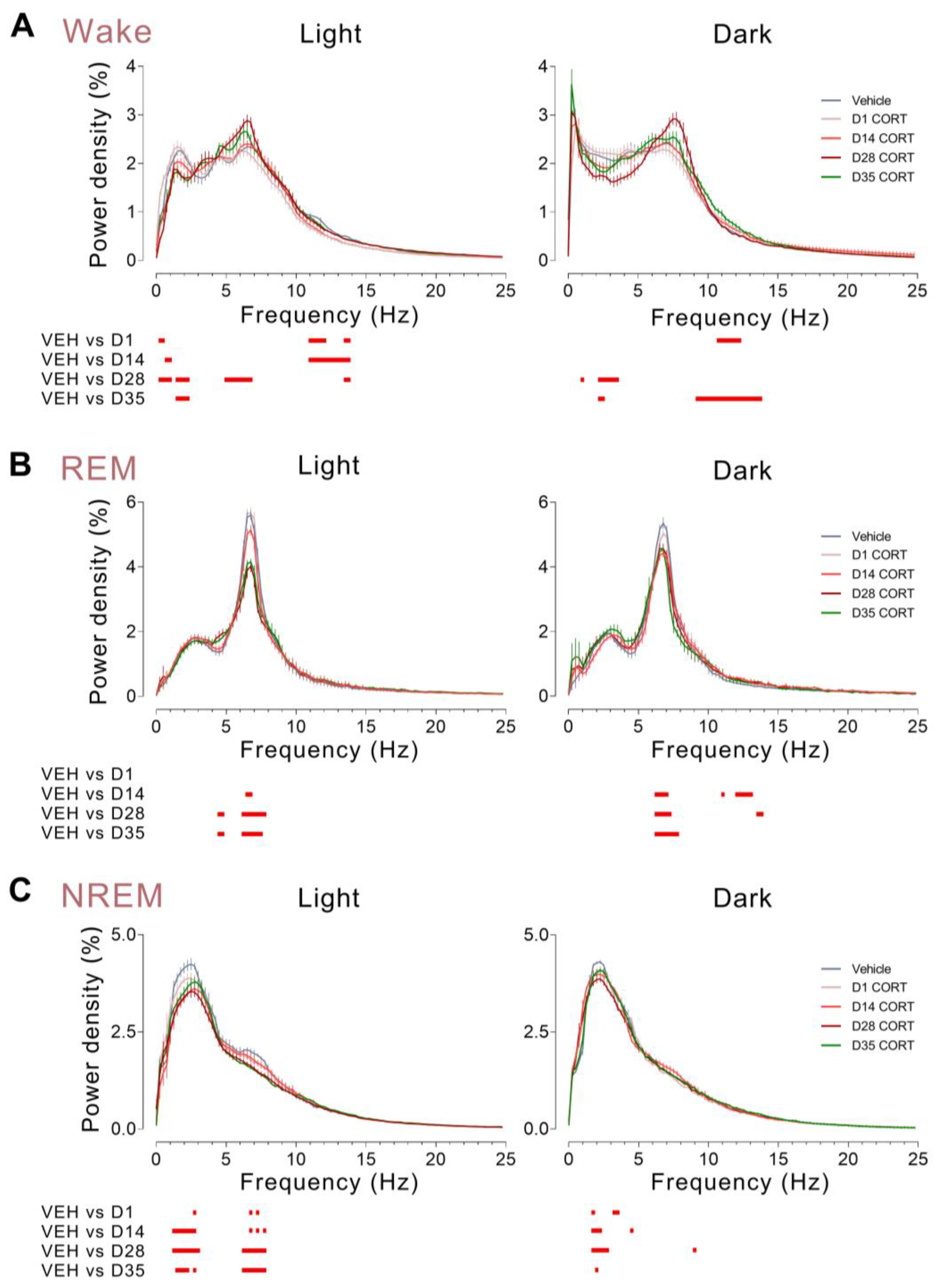
3.6. Chronic CORT Administration Decreased EEG Delta Activity and Increased Theta Activity during Wakefulness, While It Decreased EEG Theta Activity and Delta Activity during REM and NREM Sleep, Respectively
3.7. Effects of Acute and Chronic CORT Administration on c-Fos Protein Expression in Certain Brain Regions
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Aserinsky, E.; Kleitman, N. Regularly occurring periods of eye motility, and concomitant phenomena, during sleep. Science 1953, 118, 273–274. [Google Scholar] [CrossRef] [PubMed]
- Baglioni, C.; Nanovska, S.; Regen, W.; Spiegelhalder, K.; Feige, B.; Nissen, C.; Reynolds, C.F.; Riemann, D. Sleep and mental disorders: A meta-analysis of polysomnographic research. Psychol. Bull. 2016, 142, 969–990. [Google Scholar] [CrossRef] [PubMed]
- Pandi-Perumal, S.R.; Monti, J.M.; Burman, D.; Karthikeyan, R.; BaHammam, A.S.; Spence, D.W.; Brown, G.M.; Narashimhan, M. Clarifying the role of sleep in depression: A narrative review. Psychiatry Res. 2020, 291, 113239. [Google Scholar] [CrossRef] [PubMed]
- Murphy, M.J.; Peterson, M.J. Sleep Disturbances in Depression. Sleep Med. Clin. 2015, 10, 17–23. [Google Scholar] [CrossRef]
- Riemann, D.; Krone, L.B.; Wulff, K.; Nissen, C. Sleep, insomnia, and depression. Neuropsychopharmacology 2020, 45, 74–89. [Google Scholar] [CrossRef]
- Baglioni, C.; Battagliese, G.; Feige, B.; Spiegelhalder, K.; Nissen, C.; Voderholzer, U.; Lombardo, C.; Riemann, D. Insomnia as a predictor of depression: A meta-analytic evaluation of longitudinal epidemiological studies. J. Affect. Disord. 2011, 135, 10–19. [Google Scholar] [CrossRef]
- Byrne, E.M.; Timmerman, A.; Wray, N.R.; Agerbo, E. Sleep Disorders and Risk of Incident Depression: A Population Case–Control Study. Twin Res. Hum. Genet. 2019, 22, 140–146. [Google Scholar] [CrossRef]
- Scott, A.J.; Webb, T.L.; Martyn-St James, M.; Rowse, G.; Weich, S. Improving sleep quality leads to better mental health: A meta-analysis of randomised controlled trials. Sleep Med. Rev. 2021, 60, 101556. [Google Scholar] [CrossRef]
- Yang, L.; Zhao, Y.; Wang, Y.; Liu, L.; Zhang, X.; Li, B.; Cui, R. The Effects of Psychological Stress on Depression. Curr. Neuropharmacol. 2015, 13, 494–504. [Google Scholar] [CrossRef]
- Hammen, C.; Kim, E.Y.; Eberhart, N.K.; Brennan, P.A. Chronic and acute stress and the prediction of major depression in women. Depress. Anxiety 2009, 26, 718–723. [Google Scholar] [CrossRef]
- Seo, J.S.; Wei, J.; Qin, L.; Kim, Y.; Yan, Z.; Greengard, P. Cellular and molecular basis for stress-induced depression. Mol. Psychiatry 2017, 22, 1440–1447. [Google Scholar] [CrossRef] [PubMed]
- Li, S.B.; Borniger, J.C.; Yamaguchi, H.; Hedou, J.; Gaudilliere, B.; de Lecea, L. Hypothalamic circuitry underlying stress-induced insomnia and peripheral immunosuppression. Sci. Adv. 2020, 6, eabc2590. [Google Scholar] [CrossRef] [PubMed]
- Cano, G.; Mochizuki, T.; Saper, C.B. Neural circuitry of stress-induced insomnia in rats. J. Neurosci. 2008, 28, 10167–10184. [Google Scholar] [CrossRef]
- Yu, X.; Zhao, G.; Wang, D.; Wang, S.; Li, R.; Li, A.; Wang, H.; Nollet, M.; Chun, Y.Y.; Zhao, T.; et al. A specific circuit in the midbrain detects stress and induces restorative sleep. Science 2022, 377, 63–72. [Google Scholar] [CrossRef] [PubMed]
- Yasugaki, S.; Liu, C.Y.; Kashiwagi, M.; Kanuka, M.; Honda, T.; Miyata, S.; Yanagisawa, M.; Hayashi, Y. Effects of 3 Weeks of Water Immersion and Restraint Stress on Sleep in Mice. Front. Neurosci. 2019, 13, 1072. [Google Scholar] [CrossRef]
- Xu, Y.X.; Liu, G.Y.; Ji, Z.Z.; Li, Y.Y.; Wang, Y.L.; Wu, X.Y.; Liu, J.L.; Ma, D.X.; Zhong, M.K.; Gao, C.B.; et al. Restraint stress induced anxiety and sleep in mice. Front. Psychiatry 2023, 14, 1090420. [Google Scholar] [CrossRef]
- Russell, G.; Lightman, S. The human stress response. Nat. Rev. Endocrinol. 2019, 15, 525–534. [Google Scholar] [CrossRef]
- Stokes, P.E. The potential role of excessive cortisol induced by HPA hyperfunction in the pathogenesis of depression. Eur. Neuropsychopharmacol. 1995, 5 (Suppl. S1), 77–82. [Google Scholar] [CrossRef]
- Sapse, A.T. Cortisol, high cortisol diseases and anti-cortisol therapy. Psychoneuroendocrinology 1997, 22 (Suppl. S1), S3–S10. [Google Scholar] [CrossRef]
- Zhao, Y.; Ma, R.; Shen, J.; Su, H.; Xing, D.; Du, L. A mouse model of depression induced by repeated corticosterone injections. Eur. J. Pharmacol. 2008, 581, 113–120. [Google Scholar] [CrossRef]
- Vazquez-Palacios, G.; Retana-Marquez, S.; Bonilla-Jaime, H.; Velazquez-Moctezuma, J. Further definition of the effect of corticosterone on the sleep-wake pattern in the male rat. Pharmacol. Biochem. Behav. 2001, 70, 305–310. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wang, Z.; Zhang, X.; Cui, S.; Cui, X. The Correlation between Depression Behaviors and Sleep Parameters after Repeated Corticosterone Injection in Rat. Basic Clin. Pharmacol. 2014, 115, 216. [Google Scholar]
- Cui, X.Y.; Yang, G.; Cui, S.Y.; Cao, Q.; Huang, Y.L.; Ding, H.; Ye, H.; Zhang, X.Q.; Wang, Z.J.; Zhang, Y.H. Sleep patterns deteriorate over time in chronic corticosterone-treated rats. Neurosci. Lett. 2018, 682, 74–78. [Google Scholar] [CrossRef]
- Le Dantec, Y.; Hache, G.; Guilloux, J.P.; Guiard, B.P.; David, D.J.; Adrien, J.; Escourrou, P. NREM sleep hypersomnia and reduced sleep/wake continuity in a neuroendocrine mouse model of anxiety/depression based on chronic corticosterone administration. Neuroscience 2014, 274, 357–368. [Google Scholar] [CrossRef] [PubMed]
- Sousa, N.; Madeira, M.D.; Paula-Barbosa, M.M. Effects of corticosterone treatment and rehabilitation on the hippocampal formation of neonatal and adult rats. An unbiased stereological study. Brain Res. 1998, 794, 199–210. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Wang, H.J.; Wang, D.R.; Qu, W.M.; Huang, Z.L. Red light at intensities above 10 lx alters sleep-wake behavior in mice. Light Sci. Appl. 2017, 6, e16231. [Google Scholar] [CrossRef]
- Xu, Q.; Xu, X.H.; Qu, W.M.; Lazarus, M.; Urade, Y.; Huang, Z.L. A mouse model mimicking human first night effect for the evaluation of hypnotics. Pharmacol. Biochem. Behav. 2014, 116, 129–136. [Google Scholar] [CrossRef]
- Dieterich, A.; Srivastava, P.; Sharif, A.; Stech, K.; Floeder, J.; Yohn, S.E.; Samuels, B.A. Chronic corticosterone administration induces negative valence and impairs positive valence behaviors in mice. Transl. Psychiatry 2019, 9, 337. [Google Scholar] [CrossRef]
- Luo, Y.J.; Li, Y.D.; Wang, L.; Yang, S.R.; Yuan, X.S.; Wang, J.; Cherasse, Y.; Lazarus, M.; Chen, J.F.; Qu, W.M.; et al. Nucleus accumbens controls wakefulness by a subpopulation of neurons expressing dopamine D1 receptors. Nat. Commun. 2018, 9, 1576. [Google Scholar] [CrossRef]
- Shi, H.Y.; Xu, W.; Guo, H.; Dong, H.; Qu, W.M.; Huang, Z.L. Lesion of intergeniculate leaflet GABAergic neurons attenuates sleep in mice exposed to light. Sleep 2020, 43, zsz212. [Google Scholar] [CrossRef]
- Zhao, Y.N.; Jiang, J.B.; Tao, S.Y.; Zhang, Y.; Chen, Z.K.; Qu, W.M.; Huang, Z.L.; Yang, S.R. GABAergic neurons in the rostromedial tegmental nucleus are essential for rapid eye movement sleep suppression. Nat. Commun. 2022, 13, 7552. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Jiang, J.B.; Xu, W.; Zhang, M.T.; Chen, H.; Shi, H.Y.; Wang, L.; He, M.; Lazarus, M.; Li, S.Q.; et al. Parasubthalamic calretinin neurons modulate wakefulness associated with exploration in male mice. Nat. Commun. 2023, 14, 2346. [Google Scholar] [CrossRef] [PubMed]
- Qu, W.M.; Xu, X.H.; Yan, M.M.; Wang, Y.Q.; Urade, Y.; Huang, Z.L. Essential role of dopamine D2 receptor in the maintenance of wakefulness, but not in homeostatic regulation of sleep, in mice. J. Neurosci. 2010, 30, 4382–4389. [Google Scholar] [CrossRef] [PubMed]
- Kalueff, A.V.; Tuohimaa, P. Mouse grooming microstructure is a reliable anxiety marker bidirectionally sensitive to GABAergic drugs. Eur. J. Pharmacol. 2005, 508, 147–153. [Google Scholar] [CrossRef]
- Zhang, Z.; Liu, W.Y.; Diao, Y.P.; Xu, W.; Zhong, Y.H.; Zhang, J.Y.; Lazarus, M.; Liu, Y.Y.; Qu, W.M.; Huang, Z.L. Superior Colliculus GABAergic Neurons Are Essential for Acute Dark Induction of Wakefulness in Mice. Curr. Biol. 2019, 29, 637–644.e3. [Google Scholar] [CrossRef]
- Levoe, S.N.; Flannery, B.M.; Brignolo, L.; Imai, D.M.; Koehne, A.; Austin, A.T.; Bruun, D.A.; Tancredi, D.J.; Lein, P.J. Factors influencing adverse skin responses in rats receiving repeated subcutaneous injections and potential impact on neurobehavior. Curr. Neurobiol. 2014, 5, 1–10. [Google Scholar]
- Du Preez, A.; Law, T.; Onorato, D.; Lim, Y.M.; Eiben, P.; Musaelyan, K.; Egeland, M.; Hye, A.; Zunszain, P.A.; Thuret, S.; et al. The type of stress matters: Repeated injection and permanent social isolation stress in male mice have a differential effect on anxiety- and depressive-like behaviours, and associated biological alterations. Transl. Psychiatry 2020, 10, 325. [Google Scholar] [CrossRef]
- Jones, J.R.; Chaturvedi, S.; Granados-Fuentes, D.; Herzog, E.D. Circadian neurons in the paraventricular nucleus entrain and sustain daily rhythms in glucocorticoids. Nat. Commun. 2021, 12, 5763. [Google Scholar] [CrossRef]
- Blasco-Serra, A.; Gonzalez-Soler, E.M.; Cervera-Ferri, A.; Teruel-Marti, V.; Valverde-Navarro, A.A. A standardization of the Novelty-Suppressed Feeding Test protocol in rats. Neurosci. Lett. 2017, 658, 73–78. [Google Scholar] [CrossRef]
- Gould, T.D.; Dao, D.T.; Kovacsics, C.E. The Open Field Test. In Mood and Anxiety Related Phenotypes in Mice: Characterization Using Behavioral Tests; Gould, T.D., Ed.; Humana Press: Totowa, NJ, USA, 2009; pp. 1–20. [Google Scholar]
- Bullitt, E. Expression of c-fos-like protein as a marker for neuronal activity following noxious stimulation in the rat. J. Comp. Neurol. 1990, 296, 517–530. [Google Scholar] [CrossRef]
- Born, J.; DeKloet, E.R.; Wenz, H.; Kern, W.; Fehm, H.L. Gluco- and antimineralocorticoid effects on human sleep: A role of central corticosteroid receptors. Am. J. Physiol. 1991, 260 Pt 1, E183–E188. [Google Scholar] [CrossRef] [PubMed]
- Long, S.; Ding, R.; Wang, J.; Yu, Y.; Lu, J.; Yao, D. Sleep Quality and Electroencephalogram Delta Power. Front. Neurosci. 2021, 15, 803507. [Google Scholar] [CrossRef] [PubMed]
- Lo, Y.; Hsiao, Y.-T.; Chang, F.-C. Use Electroencephalogram Entropy as an Indicator to Detect Stress-Induced Sleep Alteration. Appl. Sci. 2022, 12, 4812. [Google Scholar] [CrossRef]
- Demuyser, T.; Deneyer, L.; Bentea, E.; Albertini, G.; Van Liefferinge, J.; Merckx, E.; De Prins, A.; De Bundel, D.; Massie, A.; Smolders, I. In-depth behavioral characterization of the corticosterone mouse model and the critical involvement of housing conditions. Physiol. Behav. 2016, 156, 199–207. [Google Scholar] [CrossRef] [PubMed]
- Hoifodt, R.S.; Waterloo, K.; Wang, C.E.A.; Eisemann, M.; Figenschau, Y.; Halvorsen, M. Cortisol levels and cognitive profile in major depression: A comparison of currently and previously depressed patients. Psychoneuroendocrinology 2019, 99, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Vgontzas, A.N.; Tsigos, C.; Bixler, E.O.; Stratakis, C.A.; Zachman, K.; Kales, A.; Vela-Bueno, A.; Chrousos, G.P. Chronic insomnia and activity of the stress system: A preliminary study. J. Psychosom. Res. 1998, 45, 21–31. [Google Scholar] [CrossRef]
- Sainio, E.L.; Lehtola, T.; Roininen, P. Radioimmunoassay of total and free corticosterone in rat plasma: Measurement of the effect of different doses of corticosterone. Steroids 1988, 51, 609–622. [Google Scholar] [CrossRef]
- Raadsheer, F.C.; Hoogendijk, W.J.; Stam, F.C.; Tilders, F.J.; Swaab, D.F. Increased numbers of corticotropin-releasing hormone expressing neurons in the hypothalamic paraventricular nucleus of depressed patients. Neuroendocrinology 1994, 60, 436–444. [Google Scholar] [CrossRef]
- Samuels, B.; Hen, R. Novelty-Suppressed Feeding in the Mouse. Neuromethods 2011, 63, 107–121. [Google Scholar]
- Vgontzas, A.N.; Bixler, E.O.; Lin, H.M.; Prolo, P.; Mastorakos, G.; Vela-Bueno, A.; Kales, A.; Chrousos, G.P. Chronic insomnia is associated with nyctohemeral activation of the hypothalamic-pituitary-adrenal axis: Clinical implications. J. Clin. Endocrinol. Metab. 2001, 86, 3787–3794. [Google Scholar] [CrossRef]
- Benca, R.M.; Peterson, M.J. Insomnia and depression. Sleep Med. 2008, 9, S3–S9. [Google Scholar] [CrossRef] [PubMed]
- Rush, A.J.; Erman, M.K.; Giles, D.E.; Schlesser, M.A.; Carpenter, G.; Vasavada, N.; Roffwarg, H.P. Polysomnographic Findings in Recently Drug-Free and Clinically Remitted Depressed Patients. Arch. Gen. Psychiatry 1986, 43, 878–884. [Google Scholar] [CrossRef] [PubMed]
- Palagini, L.; Baglioni, C.; Ciapparelli, A.; Gemignani, A.; Riemann, D. REM sleep dysregulation in depression: State of the art. Sleep Med. Rev. 2013, 17, 377–390. [Google Scholar] [CrossRef] [PubMed]
- Klimesch, W. EEG alpha and theta oscillations reflect cognitive and memory performance: A review and analysis. Brain Res. Rev. 1999, 29, 169–195. [Google Scholar] [CrossRef] [PubMed]
- Chapotot, F.; Buguet, A.; Gronfier, C.; Brandenberger, G. Hypothalamo-pituitary-adrenal axis activity is related to the level of central arousal: Effect of sleep deprivation on the association of high-frequency waking electroencephalogram with cortisol release. Neuroendocrinology 2001, 73, 312–321. [Google Scholar] [CrossRef] [PubMed]
- Hegde, P.; Jayakrishnan, H.R.; Chattarji, S.; Kutty, B.M.; Laxmi, T.R. Chronic stress-induced changes in REM sleep on θ oscillations in the rat hippocampus and amygdala. Brain Res. 2011, 1382, 155–164. [Google Scholar] [CrossRef]
- Duman, R.S.; Sanacora, G.; Krystal, J.H. Altered Connectivity in Depression: GABA and Glutamate Neurotransmitter Deficits and Reversal by Novel Treatments. Neuron 2019, 102, 75–90. [Google Scholar] [CrossRef]
- Magariños, A.M.; McEwen, B.S. Stress-induced atrophy of apical dendrites of hippocampal CA3c neurons: Involvement of glucocorticoid secretion and excitatory amino acid receptors. Neuroscience 1995, 69, 89–98. [Google Scholar] [CrossRef]
- Perrier, J.; Clochon, P.; Bertran, F.; Couque, C.; Bulla, J.; Denise, P.; Bocca, M.-L. Specific EEG sleep pattern in the prefrontal cortex in primary insomnia. PLoS ONE 2015, 10, e0116864. [Google Scholar] [CrossRef]
- Besedovsky, L.; Ngo, H.-V.V.; Dimitrov, S.; Gassenmaier, C.; Lehmann, R.; Born, J. Auditory closed-loop stimulation of EEG slow oscillations strengthens sleep and signs of its immune-supportive function. Nat. Commun. 2017, 8, 1984. [Google Scholar] [CrossRef]
- Hu, H.; Cui, Y.; Yang, Y. Circuits and functions of the lateral habenula in health and in disease. Nat. Rev. Neurosci. 2020, 21, 277–295. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.R.; Zhong, Y.H.; Jiang, S.; Xu, W.; Xiao, L.; Wang, Z.; Qu, W.M.; Huang, Z.L. Dysfunctions of the paraventricular hypothalamic nucleus induce hypersomnia in mice. eLife 2021, 10, e69909. [Google Scholar] [CrossRef] [PubMed]
- Peng, B.B.; Xu, Q.K.; Liu, J.; Guo, S.; Borgland, S.L.; Liu, S. Corticosterone Attenuates Reward-Seeking Behavior and Increases Anxiety via D2 Receptor Signaling in Ventral Tegmental Area Dopamine Neurons. J. Neurosci. 2021, 41, 1566–1581. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Li, W.; Ma, Y.; Tossell, K.; Harris, J.J.; Harding, E.C.; Ba, W.; Miracca, G.; Wang, D.; Li, L.; et al. GABA and glutamate neurons in the VTA regulate sleep and wakefulness. Nat. Neurosci. 2019, 22, 106–119. [Google Scholar] [CrossRef] [PubMed]
- Eban-Rothschild, A.; Rothschild, G.; Giardino, W.J.; Jones, J.R.; de Lecea, L. VTA dopaminergic neurons regulate ethologically relevant sleep-wake behaviors. Nat. Neurosci. 2016, 19, 1356–1366. [Google Scholar] [CrossRef]
- Wei, J.; Zhong, P.; Qin, L.; Tan, T.; Yan, Z. Chemicogenetic Restoration of the Prefrontal Cortex to Amygdala Pathway Ameliorates Stress-Induced Deficits. Cereb. Cortex 2018, 28, 1980–1990. [Google Scholar] [CrossRef]
- Prager, E.M.; Bergstrom, H.C.; Wynn, G.H.; Braga, M.F. The basolateral amygdala γ-aminobutyric acidergic system in health and disease. J. Neurosci. Res. 2016, 94, 548–567. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yao, Z.; Zhang, B.-X.; Chen, H.; Jiang, X.-W.; Qu, W.-M.; Huang, Z.-L. Acute or Chronic Exposure to Corticosterone Promotes Wakefulness in Mice. Brain Sci. 2023, 13, 1472. https://doi.org/10.3390/brainsci13101472
Yao Z, Zhang B-X, Chen H, Jiang X-W, Qu W-M, Huang Z-L. Acute or Chronic Exposure to Corticosterone Promotes Wakefulness in Mice. Brain Sciences. 2023; 13(10):1472. https://doi.org/10.3390/brainsci13101472
Chicago/Turabian StyleYao, Zhen, Bei-Xuan Zhang, Hui Chen, Xiao-Wei Jiang, Wei-Min Qu, and Zhi-Li Huang. 2023. "Acute or Chronic Exposure to Corticosterone Promotes Wakefulness in Mice" Brain Sciences 13, no. 10: 1472. https://doi.org/10.3390/brainsci13101472