
Circulating Endocannabinoids and N-Acylethanolamines in Individuals with Cannabis Use Disorder—Preliminary Findings

 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Methods
2.1. Participants
2.2. Extractions of NAEs and Arachidonoylglycerols
2.3. Identification and Separation Using LC-MS/MS Analysis
2.4. Analysis of Cannabinoids and Metabolites
2.5. FAAH Genotyping
2.6. Statistical Analysis
3. Results
3.1. Demographics
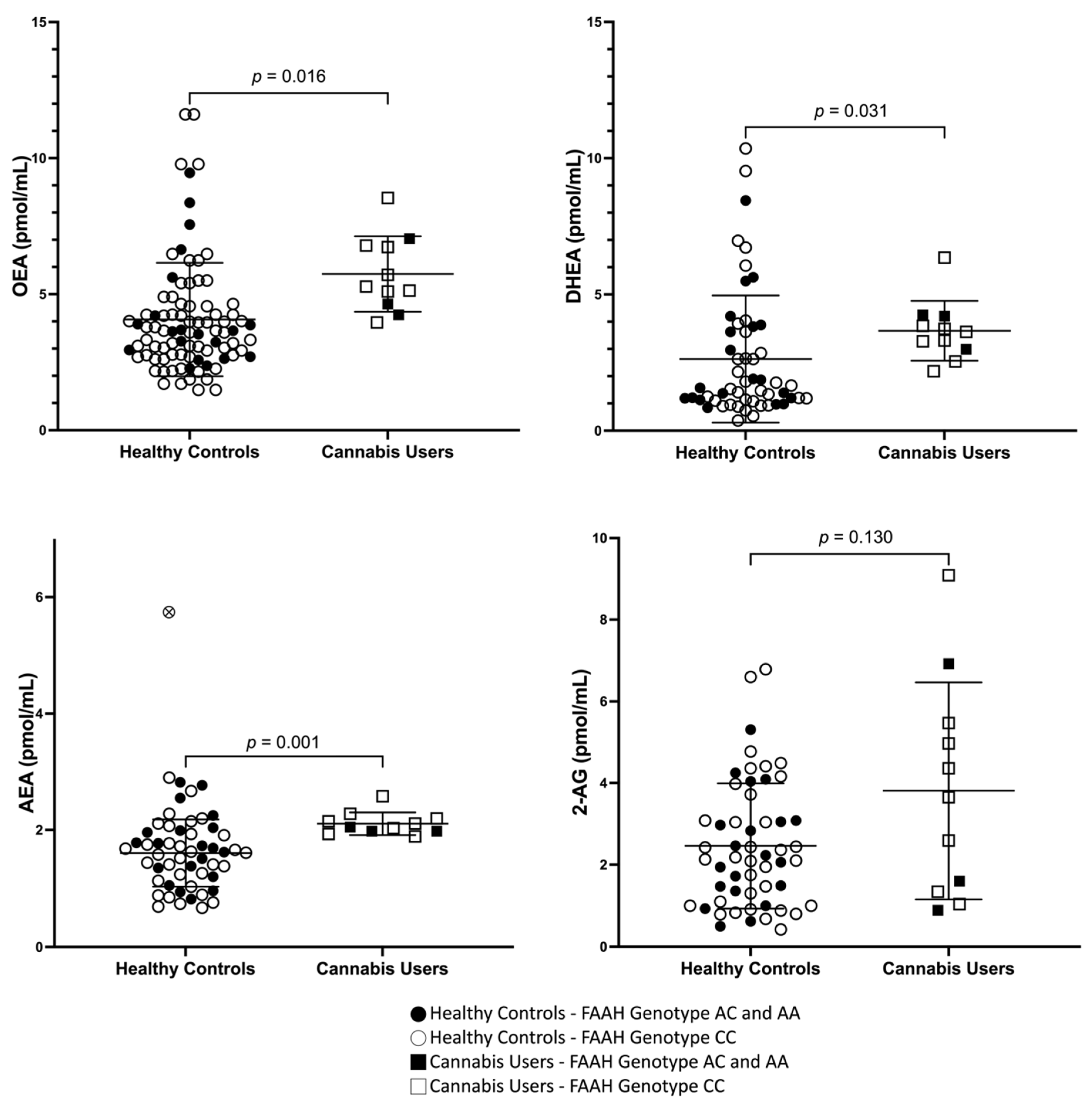
3.2. Peripheral Concentrations of AEA, OEA, DHEA, and 2-AG in Cannabis Users and Healthy Controls
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Marconi, A.; Di Forti, M.; Lewis, C.M.; Murray, R.M.; Vassos, E. Meta-analysis of the association between the level of cannabis use and risk of psychosis. Schizophr. Bull. 2016, 42, 1262–1269. [Google Scholar] [CrossRef]
- Moore, T.H.; Zammit, S.; Lingford-Hughes, A.; Barnes, T.R.E.; Jones, P.B.; Burke, M.; Lewis, G. Cannabis use and risk of psychotic or affective mental health outcomes: A systematic review. Lancet Psychiatry 2007, 370, 319–328. [Google Scholar] [CrossRef]
- Connor, J.P.; Stjepanović, D.; Le Foll, B.; Hoch, E.; Budney, A.J.; Hall, W.D. Cannabis use and cannabis use disorder. Nat. Rev. Dis. Primers 2021, 7, 16. [Google Scholar] [CrossRef]
- Marshall, K.; Gowing, L.; Ali, R.; Le Foll, B. Pharmacotherapies for cannabis dependence. Cochrane Database Syst. Rev. 2014, 12, CD008940. [Google Scholar] [CrossRef]
- Bobb, A.J.; Hill, K.P. Behavioral Interventions and Pharmacotherapies for Cannabis Use Disorder. Curr. Treat. Options Psychiatry 2014, 1, 163–174. [Google Scholar] [CrossRef]
- Nielsen, S.; Sabioni, P.; Gowing, L.; Le Foll, B. Pharmacotherapies for Cannabis Use Disorders: Clinical Challenges and Promising Therapeutic Agents. In Substance Use Disorders; Springer: Cham, Switzerland, 2019; pp. 1–18. [Google Scholar]
- United Nations Office On Drugs and Crime. World Drug Report; Sales No. E.15.XI.6; United Nations Publication: New York, NY, USA, 2015. [Google Scholar]
- Matheson, J.; Le Foll, B. Cannabis legalization and acute harm from high potency cannabis products: A narrative review and recommendations for public health. Front. Psychiatry 2020, 11, 591979. [Google Scholar] [CrossRef]
- D’Souza, D.C.; DiForti, M.; Ganesh, S.; George, T.P.; Hall, W.; Hjorthøj, C.; Howes, O.; Keshavan, M.; Murray, R.M.; Nguyen, T.B.; et al. Consensus paper of the WFSBP task force on cannabis, cannabinoids and psychosis. World J. Biol. Psychiatry 2022, 23, 719–742. [Google Scholar] [CrossRef]
- Matsuda, L.A.; Lolait, S.J.; Brownstein, M.J.; Young, A.C.; Bonner, T.I. Structure of a cannabinoid receptor and functional expression of the cloned cDNA. Nature 1990, 346, 561–564. [Google Scholar] [CrossRef]
- Howlett, A.C.; Johnson, M.R.; Melvin, L.S.; Milne, G.M. Nonclassical cannabinoid analgetics inhibit adenylate cyclase: Development of a cannabinoid receptor model. Mol. Pharmacol. 1988, 33, 297–302. [Google Scholar]
- Howlett, A.C. Pharmacology of cannabinoid receptors. Annu. Rev. Pharmacol. Toxicol. 1995, 35, 607–634. [Google Scholar] [CrossRef]
- Bisogno, T.; Berrendero, F.; Ambrosino, G.; Cebeira, M.; Ramos, J.A.; Fernandez-Ruiz, J.J.; Di Marzo, V. Brain regional distribution of endocannabinoids: Implications for their biosynthesis and biological function. Biochem. Biophys. Res. Commun. 1999, 256, 377–380. [Google Scholar] [CrossRef] [PubMed]
- Cristino, L.; Bisogno, T.; Di Marzo, V. Cannabinoids and the expanded endocannabinoid system in neurological disorders. Nat. Rev. Neurol. 2020, 16, 9–29. [Google Scholar] [CrossRef] [PubMed]
- Maccarrone, M.; Bab, I.; Bíró, T.; Cabral, G.A.; Dey, S.K.; Di Marzo, V.; Konje, J.C.; Kunos, G.; Mechoulam, R.; Pacher, P.; et al. Endocannabinoid signaling at the periphery: 50 years after THC. Trends Pharmacol. Sci. 2015, 36, 277–296. [Google Scholar] [CrossRef] [PubMed]
- Nyilas, R.; Dudok, B.; Urbán, G.M.; Mackie, K.; Watanabe, M.; Cravatt, B.F.; Freund, T.F.; Katona, I. Enzymatic machinery for endocannabinoid biosynthesis associated with calcium stores in glutamatergic axon terminals. J. Neurosci. 2008, 28, 1058–1063. [Google Scholar] [CrossRef]
- Rodríguez-Muñoz, M.; Sánchez-Blázquez, P.; Callado, L.F.; Meana, J.J.; Garzón-Niño, J. Schizophrenia and depression, two poles of endocannabinoid system deregulation. Transl. Psychiatry 2017, 7, 1291. [Google Scholar] [CrossRef]
- Hillard, C. Circulating Endocannabinoids: From Whence Do They Come and Where are They Going? Neuropsychopharmacology 2018, 43, 155–172. [Google Scholar] [CrossRef]
- Ceccarini, J.; Kuepper, R.; Kemels, D.; van Os, J.; Henquet, C.; Van Laere, K. [F]MK-9470 PET measurement of cannabinoid CB receptor availability in chronic cannabis users. Addict. Biol. 2015, 20, 357–367. [Google Scholar] [CrossRef]
- Sugiura, T.; Kondo, S.; Sukagawa, A.; Nakane, S.; Shinoda, A.; Itoh, K.; Yamashita, A.; Waku, K. 2-Arachidonoylgylcerol: A possible endogenous cannabinoid receptor ligand in brain. Biochem. Biophys. Res. Commun. 1995, 215, 89–97. [Google Scholar] [CrossRef]
- Devane, W.A.; Hanuš, L.; Breuer, A.; Pertwee, R.G.; Stevenson, L.A.; Griffin, G.; Gibson, D.; Mandelbaum, A.; Etinger, A.; Mechoulam, R. Isolation and structure of a brain constituent that binds to the cannabinoid receptor. Science 1992, 258, 1946–1949. [Google Scholar] [CrossRef]
- Hillard, C.J. Biochemistry and pharmacology of the endocannabinoids arachidonylethanolamide and 2-arachidonylglycerol. Prostaglandins Other Lipid Médiat 2000, 61, 3–18. [Google Scholar] [CrossRef]
- Melis, M.; Pistis, M.; Perra, S.; Muntoni, A.L.; Pillolla, G.; Gessa, G.L. Endocannabinoids mediate presynaptic inhibition of glutamatergic transmission in rat ventral tegmental area dopamine neurons through activation of CB1 receptors. J. Neurosci. 2004, 24, 53–62. [Google Scholar] [CrossRef] [PubMed]
- Zygmunt, P.M.; Petersson, J.; Andersson, D.A.; Chuang, H.-H.; Sørgård, M.; Di Marzo, V.; Julius, D.; Högestätt, E.D. Vanilloid receptors on sensory nerves mediate the vasodilator action of anandamide. Nature 1999, 400, 452–457. [Google Scholar] [CrossRef]
- Okamoto, Y.; Morishita, J.; Tsuboi, K.; Tonai, T.; Ueda, N. Molecular characterization of a phospholipase D generating anandamide and its congeners. J. Biol. Chem. 2004, 27, 5298–5305. [Google Scholar] [CrossRef]
- Dinh, T.P.; Carpenter, D.; Leslie, F.M.; Freund, T.F.; Katona, I.; Sensi, S.L.; Piomelli, D. Brain monoglyceride lipase participating in endocannabinoid inactivation. Proc. Natl. Acad. Sci. USA 2002, 99, 10819–10824. [Google Scholar] [CrossRef]
- Mechoulam, R.; Parker, L.A. The endocannabinoid system and the brain. Annu. Rev. Psychol. 2013, 64, 21–47. [Google Scholar] [CrossRef] [PubMed]
- D’Souza, D.C.; Cortes-Briones, J.; Creatura, G.; Bluez, G.; Thurnauer, H.; Deaso, E.; Bielen, K.; Surti, T.; Radhakrishnan, R.; Gupta, A.; et al. Efficacy and safety of a fatty acid amide hydrolase inhibitor (PF-04457845) in the treatment of cannabis withdrawal and dependence in men: A double-blind, placebo-controlled, parallel group, phase 2a single-site randomised controlled trial. Lancet Psychiatry 2019, 6, 35–45. [Google Scholar] [CrossRef] [PubMed]
- Schlosburg, J.E.; Carlson, B.L.A.; Ramesh, D.; Abdullah, R.A.; Long, J.Z.; Cravatt, B.F.; Lichtman, A.H. Inhibitors of endocannabinoid-metabolizing enzymes reduce precipitated withdrawal responses in THC-dependent mice. AAPS J. 2009, 11, 342–352. [Google Scholar] [CrossRef]
- Haney, M.; Bedi, G.; Cooper, Z.D.; Herrmann, E.S.; Reed, S.C.; Foltin, R.W.; Patel, S. Impact of cyclooxygenase-2 inhibition on cannabis withdrawal and circulating endocannabinoids in daily cannabis smokers. Addict. Biol. 2022, 27, e13183. [Google Scholar] [CrossRef]
- Bilbao, A.; Serrano, A.; Cippitelli, A.; Pavón, F.J.; Giuffrida, A.; Suárez, J.; García-Marchena, N.; Baixeras, E.; Gómez de Heras, R.; Orio, L.; et al. Role of the satiety factor oleoylethanolamide in alcoholism. Addict. Biol. 2016, 21, 859–872. [Google Scholar] [CrossRef]
- Justinova, Z.; Munzar, P.; Panlilio, L.V.; Yasar, S.; Redhi, G.H.; Tanda, G.; Goldberg, S.R. Blockade of THC-seeking behavior and relapse in monkeys by the cannabinoid CB(1)-receptor antagonist rimonabant. Neuropsychopharmacology 2008, 33, 2870–2877. [Google Scholar] [CrossRef]
- Petersen, G.; Sørensen, C.; Schmid, P.C.; Artmann, A.; Tang-Christensen, M.; Hansen, S.H.; Larsen, P.J.; Schmid, H.H.; Hansen, H.S. Intestinal levels of anandamide and oleoylethanolamide in food-deprived rats are regulated through their precursors. Biochim. Biophys. Acta (BBA)-Mol. Cell Biol. Lipids 2006, 1761, 43–150. [Google Scholar] [CrossRef] [PubMed]
- Lo Verme, J.; Gaetani, S.; Fu, J.; Oveisi, F.; Burton, K.; Piomelli, D. Regulation of food intake by oleoylethanolamide. Cell. Mol. Life Sci. 2005, 62, 708–716. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.Y.; Spector, A.A. Synaptamide, endocannabinoid-like derivative of docosahexaenoic acid with cannabinoid-independent function. Prostaglandins Leukot. Essent. Fat. Acids 2013, 88, 121–125. [Google Scholar] [CrossRef]
- Di Marzo, V.; Melck, D.; Bisogno, T.; De Petrocellis, L. Endocannabinoids: Endogenous cannabinoid receptor ligands with neuromodulatory action. Trends Neurosci. 1998, 21, 521–528. [Google Scholar] [CrossRef] [PubMed]
- Meijerink, J.; Poland, M.; Balvers, M.G.J.; Plastina, P.; Lute, C.; Dwarkasing, J.; van Norren, K.; Witkamp, R.F. Inhibition of COX-2-mediated eicosanoid production plays a major role in the anti-inflammatory effects of the endocannabinoid N-docosahexaenoylethanolamine (DHEA) in macrophages. Br. J. Pharmacol. 2015, 172, 24–37. [Google Scholar] [CrossRef]
- Hirvonen, J.; Goodwin, R.S.; Li, C.T.; Terry, G.E.; Zoghbi, S.S.; Morse, C.; Pike, V.W.; Volkow, N.D.; Huestis, M.A.; Innis, R.B. Reversible and regionally selective downregulation of brain cannabinoid CB1 receptors in chronic daily cannabis smokers. Mol. Psychiatry 2012, 17, 642–649. [Google Scholar] [CrossRef]
- Zanotti-Fregonara, P.; Hirvonen, J.; Lyoo, C.H.; Zoghbi, S.S.; Rallis-Frutos, D.; Huestis, M.A.; Morse, C.; Pike, V.W.; Innis, R.B. Population-based input function modeling for [(18)F]FMPEP-d 2, an inverse agonist radioligand for cannabinoid CB1 receptors: Validation in clinical studies. PLoS ONE 2013, 8, e60231. [Google Scholar] [CrossRef]
- Di Marzo, V.; Berrendero, F.; Bisogno, T.; Gonzalez, S.; Cavaliere, P.; Romero, J.; Cebeira, M.; Ramos, J.A.; Fernandez-Ruiz, J.J. Enhancement of anandamide formation in the limbic forebrain and reduction of endocannabinoid contents in the striatum of delta9-tetrahydrocannabinol-tolerant rats. J. Neurochem. 2000, 74, 1627–1635. [Google Scholar] [CrossRef]
- Boileau, I.; Mansouri, E.; Williams, B.; Le Foll, B.; Rusjan, P.; Mizrahi, R.; Tyndale, R.F.; Huestis, M.A.; Payer, D.E.; Wilson, A.A.; et al. Fatty Acid Amide Hydrolase Binding in Brain of Cannabis Users: Imaging With the Novel Radiotracer [11C]CURB. Biol. Psychiatry 2016, 80, 691–701. [Google Scholar] [CrossRef]
- Ahmed, F.; Torrens, A.; Mahler, S.V.; Ferlenghi, F.; Huestis, M.A.; Piomelli, D. A Sensitive Ultrahigh-Performance Liquid Chromatography/Tandem Mass Spectrometry Method for the Simultaneous Analysis of Phytocannabinoids and Endocannabinoids in Plasma and Brain. Cannabis Cannabinoid Res. 2022; ahead of print. [Google Scholar] [CrossRef]
- Gonzalez, S.; Fernandez-Ruiz, J.; Di Marzo, V.; Hernandez, M.; Arevalo, C.; Nicanor, C.; Cascio, M.G.; Ambrosio, E.; Ramos, J.A. Behavioral and molecular changes elicited by acute administration of SR141716 to Delta9-tetrahydrocannabinol-tolerant rats: An experimental model of cannabinoid abstinence. Drug Alcohol Depend. 2004, 74, 159–170. [Google Scholar] [CrossRef] [PubMed]
- Ellgren, M.; Artmann, A.; Tkalych, O.; Gupta, A.; Hansen, H.S.; Hansen, S.H.; Devi, L.A.; Hurd, Y.L. Dynamic changes of the endogenous cannabinoid and opioid mesocorticolimbic systems during adolescence: THC effects. Eur. Neuropsychopharmacol. 2008, 18, 826–834. [Google Scholar] [CrossRef] [PubMed]
- Thieme, U.; Schelling, G.; Hauer, D.; Greif, R.; Dame, T.; Laubender, R.P.; Bernhard, W.; Thieme, D.; Campolongo, P.; Theiler, L. Quantification of anandamide and 2-arachidonoylglycerol plasma levels to examine potential influences of tetrahydrocannabinol application on the endocannabinoid system in humans. Drug Test. Anal. 2013, 6, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Kearney-Ramos, T.; Herrmann, E.S.; Belluomo, I.; Matias, I.; Vallée, M.; Monlezun, S.; Piazza, P.V.; Haney, M. The relationship between circulating endogenous cannabinoids and the effects of smoked cannabis. Cannabis Cannabinoid Res. 2022; ahead of print. [Google Scholar] [CrossRef] [PubMed]
- Walter, C.; Ferreirós, N.; Bishay, P.; Geisslinger, G.; Tegeder, I.; Lötsch, J. Exogenous Delta9-tetrahydrocannabinol influences circulating endogenous cannabinoids in humans. J. Clin. Psychopharmacol. 2013, 33, 699–705. [Google Scholar] [CrossRef]
- Potvin, S.; Kouassi, É.; Lipp, O.; Bouchard, R.-H.; Roy, M.-A.; Demers, M.-F.; Gendron, A.; Astarita, G.; Piomelli, D.; Stip, E. Endogenous cannabinoids in patients with schizophrenia and substance use disorder during quetiapine therapy. J. Psychopharmacol. 2008, 22, 262–269. [Google Scholar] [CrossRef] [PubMed]
- Bassir Nia, A.; Gibson, C.L.; Spriggs, S.A.; Jankowski, S.E.; DeFrancisco, D.; Swift, A.; Perkel, C.; Galynker, I.; Honrao, C.; Makriyannis, A.; et al. Cannabis use is associated with low plasma endocannabinoid Anandamide in individuals with psychosis. J. Psychopharmacol. 2023, 37, 484–489. [Google Scholar] [CrossRef]
- Morgan, C.J.; Page, E.; Schaefer, C.; Chatten, K.; Manocha, A.; Gulati, S.; Curran, H.V.; Brandner, B.; Leweke, F.M. Cerebrospinal fluid anandamide levels, cannabis use and psychotic-like symptoms. Br. J. Psychiatry 2013, 202, 381–382. [Google Scholar] [CrossRef]
- Leweke, F.M.; Giuffrida, A.; Koethe, D.; Schreiber, D.; Nolden, B.M.; Kranaster, L.; Neatby, M.A.; Schneider, M.; Gerth, C.W.; Hellmich, M.; et al. Anandamide levels in cerebrospinal fluid of first- episode schizophrenic patients: Impact of cannabis use. Schizophr. Res. 2007, 94, 29–36. [Google Scholar] [CrossRef]
- Cropsey, K.L.; Trent, L.R.; Clark, C.B.; Stevens, E.N.; Lahti, A.C.; Hendricks, P.S. How low should you go? Determining the optimal cutoff for exhaled carbon monoxide to confirm smoking abstinence when using cotinine as reference. Nicotine Tob. Res. Off. J. Soc. Res. Nicotine Tob. 2014, 16, 1348–1355. [Google Scholar] [CrossRef]
- Cooper, A.M.; Sobell, M.B.; Sobell, L.C.; Maisto, S.A. Validity of alcoholic’s self-reports: Duration data. Int. J. Addict. 1981, 16, 401–406. [Google Scholar] [CrossRef] [PubMed]
- Hitsman, B.; Shen, B.; Cohen, R.A.; Morissette, S.B.; Drobes, D.J.; Spring, B.; Schneider, K.; Evans, D.E.; Gulliver, S.B.; Kamholz, B.W.; et al. Measuring smoking-related preoccupation and compulsive drive: Evaluation of the obsessive compulsive smoking scale. Psychopharmacology 2020, 211, 377–387. [Google Scholar] [CrossRef] [PubMed]
- Heishman, S.J.; Evans, R.J.; Singleton, E.G.; Levin, K.H.; Copersino, M.L.; Gorelick, D.A. Reliability and validity of a short form of the Marijuana Craving Questionnaire. Drug Alcohol. Depend. 2009, 102, 35–40. [Google Scholar] [CrossRef]
- Beck, A.T.; Ward, C.H.; Mendelson, M.; Mock, J.; Erbaugh, J. An inventory for measuring depression. Arch. Gen. Psychiatry 1961, 4, 561–571. [Google Scholar] [CrossRef] [PubMed]
- Budney, A.J.; Novy, P.L.; Hughes, J.R. Marijuana withdrawal among adults seeking treatment for marijuana dependence. Addiction 1999, 94, 1311–1322. [Google Scholar] [CrossRef]
- First, M.B.; Gibbon, M.; Spitzer, R.L.; Benjamin, L.S.; Williams, J.B.W. User’s Guide for the Structured Clinical Interview for DSM-IV Axis I Disorders SCID-I: Clinician Version; American Psychiatric Publishing: Washington, DC, USA, 1997; pp. 12–85. [Google Scholar]
- Lin, L.; Metherel, A.H.; Jones, P.J.; Bazinet, R.P. Fatty acid amide hydrolase (FAAH) regulates hypercapnia/ischemia-induced increases in n-acylethanolamines in mouse brain. J. Neurochem. 2017, 142, 662–671. [Google Scholar] [CrossRef]
- Huestis, M.A.; Henningfield, J.E.; Cone, E.J. Absorption of THC and formation of 11-OH-THC and THCCOOH during and after smoking marijuana. J. Anal. Toxicol. 1992, 16, 276–282. [Google Scholar] [CrossRef] [PubMed]
- Best, L.M.; Williams, B.; Le Foll, B.; Mansouri, E.; Bazinet, R.P.; Lin, L.; De Luca, V.; Lagzdins, D.; Rusjan, P.; Tyndale, R.F.; et al. Lower brain fatty acid amide hydrolase in treatment-seeking patients with alcohol use disorder: A positron emission tomography study with [C-11] CURB. Neuropsychopharmacology 2020, 45, 1289–1296. [Google Scholar] [CrossRef]
- Bergamaschi, M.M.; Karschner, E.L.; Goodwin, R.S.; Scheidweiler, K.B.; Hirvonen, J.; Queiroz, R.H.; Huestis, M.A. Impact of prolonged cannabinoid excretion in chronic daily cannabis smokers’ blood on per se drugged driving laws. Clin. Chem. 2013, 59, 519–526. [Google Scholar] [CrossRef]
- Karschner, E.L.; Schwilke, E.W.; Lowe, R.H.; Darwin, W.D.; Herning, R.I.; Cadet, J.L.; Huestis, M.A. Implications of plasma Delta9-tetrahydrocannabinol, 11-hydroxy-THC, and 11-nor-9-carboxy-THC concentrations in chronic cannabis smokers. J. Anal. Toxicol. 2009, 33, 469–477. [Google Scholar] [CrossRef]
- Desrosiers, N.A.; Lee, D.; Concheiro-Guisan, M.; Scheidweiler, K.B.; Gorelick, D.A.; Huestis, M.A. Urinary cannabinoid disposition in occasional and frequent smokers: Is THC-glucuronide in sequential urine samples a marker of recent use in frequent smokers? Clin. Chem. 2014, 60, 361–372. [Google Scholar] [CrossRef]
- Dincheva, I.; Drysdale, A.T.; Hartley, C.A.; Johnson, D.C.; Jing, D.; King, E.C.; Ra, S.; Gray, J.M.; Yang, R.; DeGruccio, A.M.; et al. FAAH genetic variation enhances fronto-amygdala function in mouse and human. Nat. Commun. 2015, 6, 6395. [Google Scholar] [CrossRef]
- Sipe, J.C.; Scott, T.M.; Murray, S.; Harismendy, O.; Simon, G.M.; Cravatt, B.F.; Waalen, J. Biomarkers of endocannabinoid system activation in severe obesity. PLoS ONE 2010, 5, e8792. [Google Scholar] [CrossRef] [PubMed]
- Chiang, K.P.; Gerber, A.L.; Sipe, J.C.; Cravatt, B.F. Reduced cellular expression and activity of the P129T mutant of human fatty acid amide hydrolase: Evidence for a link between defects in the endocannabinoid system and problem drug use. Human. Mol. Genet. 2004, 18, 2113–2119. [Google Scholar] [CrossRef]
- Spohrs, J.; Ulrich, M.; Grön, G.; Prost, M.; Plener, P.L.; Fegert, J.M.; Bindila, L.; Abler, B. Fear extinction learning and anandamide: An fMRI study in healthy humans. Transl. Psychiatry 2021, 11, 161. [Google Scholar] [CrossRef]
- Mato, S.; Chevaleyre, V.; Robbe, D.; Pazos, A.; Castillo, P.E.; Manzoni, O.J. A single in-vivo exposure to Δ9THC blocks endocannabinoid-mediated synaptic plasticity. Nat. Neurosci. 2004, 7, 585. [Google Scholar] [CrossRef] [PubMed]
- Kelley, B.G.; Thayer, S.A. Δ9-Tetrahydrocannabinol antagonizes endocannabinoid modulation of synaptic transmission between hippocampal neurons in culture. Neuropharmacology 2004, 5, 709–715. [Google Scholar] [CrossRef]
- De Petrocellis, L.; Ligresti, A.; Moriello, A.S.; Allarà, M.; Bisogno, T.; Petrosino, S.; Stott, C.G.; Di Marzo, V. Effects of cannabinoids and cannabinoid-enriched Cannabis extracts on TRP channels and endocannabinoid metabolic enzymes. Br. J. Pharmacol. 2010, 163, 1479–1494. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.-L.; Jung, K.-M.; Fotio, Y.; Squire, E.; Palese, F.; Lin, L.; Torrens, A.; Ahmed, F.; Tagne, A.M.; Ramirez, J.; et al. Frequent low-dose Δ9-tetrahydrocannabinol in adolescence disrupts microglia homeostasis and disables responses to microbial infection and social stress in young adulthood. Biol. Psychiatry 2022, 92, 845–860. [Google Scholar] [CrossRef]
- Desfossés, J.; Stip, E.; Bentaleb, L.A.; Lipp, O.; Chiasson, J.-P.; Furtos, A.; Venne, K.; Kouassi, E.; Potvin, S. Plasma endocannabinoid alterations in individuals with substance use disorder are dependent on the “mirror effect” of schizophrenia. Front. Psychiatry 2012, 3, 85. [Google Scholar] [CrossRef]
- Spagnolo, P.A.; Ramchandani, V.A.; Schwandt, M.L.; Kwako, L.E.; George, D.T.; Mayo, L.M.; Hillard, C.J.; Heilig, M. FAAH gene variation moderates stress response and symptom severity in patients with posttraumatic stress disorder and comorbid alcohol dependence. Alcohol. Clin. Exp. Res. 2016, 40, 2426–2434. [Google Scholar] [CrossRef]
- Mayo, L.M.; Asratian, A.; Lindé, J.; Morena, M.; Haataja, R.; Hammar, V.; Augier, G.; Hill, M.N.; Heilig, M. Elevated anandamide, enhanced recall of fear extinction, and attenuated stress responses following inhibition of fatty acid amide hydrolase: A randomized, controlled experimental medicine trial. Biol. Psychiatry 2019, 87, 538–547. [Google Scholar] [CrossRef]
- Villares, J. Chronic use of marijuana decreases cannabinoid receptor binding and mRNA expression in the human brain. Neuroscience 2007, 145, 323–334. [Google Scholar] [CrossRef]
- Sparling, P.B.; Giuffrida, A.; Piomelli, D.; Rosskopf, L.; Dietrich, A. Exercise activates the endocannabinoid system. Neuroreport 2003, 14, 2209–2211. [Google Scholar] [CrossRef] [PubMed]
- Heyman, E.; Gamelin, F.-X.; Goekint, M.; Piscitelli, F.; Roelands, B.; Leclair, E.; Di Marzo, V.; Meeusen, R. Intense exercise increases circulating endocannabinoid and BDNF levels in humans—Possible implications for reward and depression. Psychoneuroendocrinology 2012, 37, 844–851. [Google Scholar] [CrossRef] [PubMed]
- Kuipers, E.N.; Kantae, V.; Maarse, B.C.E.; Van den Berg, S.M.; Van Eenige, R.; Nahon, K.J.; Reifel-Miller, A.; Coskun, T.; de Winther, M.P.J.; Lutgens, E.; et al. High fat diet increases circulating endocannabinoids accompanied by increased synthesis enzymes in adipose tissue. Front. Physiol. 2019, 9, 1913. [Google Scholar] [CrossRef]
- Bluher, M.; Engeli, S.; Kloting, N.; Berndt, J.; Fasshauer, M.; Bátkai, S.; Pacher, P.; Schon, M.R.; Jordan, J.; Stumvoll, M. Dysregulation of the peripheral and adipose tissue endocannabinoid system in human abdominal obesity. Diabetes 2006, 55, 3053–3060. [Google Scholar] [CrossRef]
- Cedernaes, J.; Fanelli, F.; Fazzini, A.; Pagotto, U.; Broman, J.-E.; Vogel, H.; Dickson, S.L.; Schiöth, H.B.; Benedict, C. Sleep restriction alters plasma endocannabinoids concentrations before but not after exercise in humans. Psychoneuroendocrinology 2016, 74, 258–268. [Google Scholar] [CrossRef] [PubMed]
- DiPatrizio, N.V.; Astarita, G.; Schwartz, G.; Li, X.; Piomelli, D. Endocannabinoid signal in the gut controls dietary fat intake. Proc. Natl. Acad. Sci. USA 2011, 108, 12904–12908. [Google Scholar] [CrossRef]
- Koethe, D.; Schreiber, D.; Giuffrida, A.; Mauss, C.; Faulhaber, J.; Heydenreich, B.; Hellmich, M.; Graf, R.; Klosterkötter, J.; Piomelli, D.; et al. Sleep deprivation increases oleoylethanolamide in human cerebrospinal fluid. J. Neural Transm. 2009, 116, 301–305. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Characteristic | Healthy Controls (n = 54) Frequencies/Mean ± SD (Range) | Cannabis Users (n = 11) Frequencies/Mean ± SD (Range) | p Value x2 |
|---|---|---|---|
| Sex (Females/Males), n | 28/26 | 4/7 | 0.35 x2 = 0.88 |
| Age, Years | 28.7 ± 11.5 (19–58) | 33.4 ± 9.2 (20–44) | 0.21 |
| Racial, Ethnic Categories (White/Asian/Black/Hispanic), n | (30/10/12/2) | (8/0/3/0) | 0.56 x2 = 3.006 |
| Body mass Index | 24.3 ± 3.1 (18.7–32.9) | 24.6 ± 4.7 (19.0–30.5) | 0.79 |
| FAAH Genotype (rs324420, C385A), n | 34(CC), 16(AC), 4(AA) | 8(CC), 2(AC), 1(AA) | 0.74 x2 = 0.60 |
| Education, Years | 15.3 ± 2.0 (11–22) | 13.9 ± 3.8 (9–19) | 0.077 |
| Tobacco Cig Smokers (>1 cigarette/day), n | 10 | 6 | 0.01 ** x2 = 6.392 |
| Tobacco Cig Per Day (in smokers) | 10.6 ± 9.7 (1.0–25.0) | 14.8 ± 12.5 (2–35) | 0.50 |
| Alcoholic Drinks Per Week | 4.7 ± 5.7 (0–24.8) | 2.5 ± 2.9 (0–7.8) | 0.065 |
| Fagerstrom Test for Nicotine Dependence, | 3.6 ± 3.2 (0–8) | 3.8 ± 3.3 (1–9) | 0.93 |
| Cannabis Ever Used, n | 32 | 11 | 0.01 ** x2 = 6.20 |
| Characteristic | Mean (SD) | Range (Min–Max) |
|---|---|---|
| Cannabis Age of Onset, Years | 15.73 (3.88) | 10.00–22.00 |
| Current Cannabis Use/Week (Grams) | 7.72 (4.29) | 3.50–14.00 |
| Average Cannabis Use Joints/Week | 19.83 (9.92) | 2.10–35.00 |
| Cannabis Total Years Used | 18.00 (10.35) | 5.00–33.0 |
| Severity of Dependence Scale | 3.27 (2.24) | 0.00–7.00 |
| Marijuana Checklist (n = 10) | 6.00 (3.92) | 1.00–12.00 |
| Obsessive compulsive smoking scale (n = 10) | 14.20 (7.11) | 5.00–25.00 |
| Marijuana Craving Questionnaire | ||
| Compulsivity | 1.58 (0.67) | 1.00–3.00 |
| Emotionality | 3.06 (1.26) | 1.00–5.30 |
| Expectancy | 4.18 (1.27) | 2.00–6.30 |
| Purposefulness | 4.33 (1.56) | 1.00–6.30 |
| Total | 13.15 (4.04) | 5.00–19.30 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boachie, N.; Gaudette, E.; Bazinet, R.P.; Lin, L.; Tyndale, R.F.; Mansouri, E.; Huestis, M.A.; Tong, J.; Le Foll, B.; Kish, S.J.; et al. Circulating Endocannabinoids and N-Acylethanolamines in Individuals with Cannabis Use Disorder—Preliminary Findings. Brain Sci. 2023, 13, 1375. https://doi.org/10.3390/brainsci13101375
Boachie N, Gaudette E, Bazinet RP, Lin L, Tyndale RF, Mansouri E, Huestis MA, Tong J, Le Foll B, Kish SJ, et al. Circulating Endocannabinoids and N-Acylethanolamines in Individuals with Cannabis Use Disorder—Preliminary Findings. Brain Sciences. 2023; 13(10):1375. https://doi.org/10.3390/brainsci13101375
Chicago/Turabian StyleBoachie, Nadia, Erin Gaudette, Richard P. Bazinet, Lin Lin, Rachel F. Tyndale, Esmaeil Mansouri, Marilyn A. Huestis, Junchao Tong, Bernard Le Foll, Stephen J. Kish, and et al. 2023. "Circulating Endocannabinoids and N-Acylethanolamines in Individuals with Cannabis Use Disorder—Preliminary Findings" Brain Sciences 13, no. 10: 1375. https://doi.org/10.3390/brainsci13101375