
Transcriptome Sequencing Reveal That Rno-Rsf1_0012 Participates in Levodopa-Induced Dyskinesia in Parkinson’s Disease Rats via Binding to Rno-mir-298-5p

Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. 6-OHDA Lesion and L-DOPA Administration

2.3. Immunofluorescence Analysis
2.4. Western Blotting
2.5. CircRNAs Extraction and Sequencing
2.6. Differential Expression Analysis of circRNAs
2.7. GO and KEGG Analyses
2.8. qRT-PCR
2.9. Competing Endogenous RNAs Network
2.10. Fluorescence In Situ Hybridization (FISH)
2.11. Dual-Luciferase Reporter Assay
2.12. Statistical Analysis
3. Results
3.1. Validation of the LID Rat Models
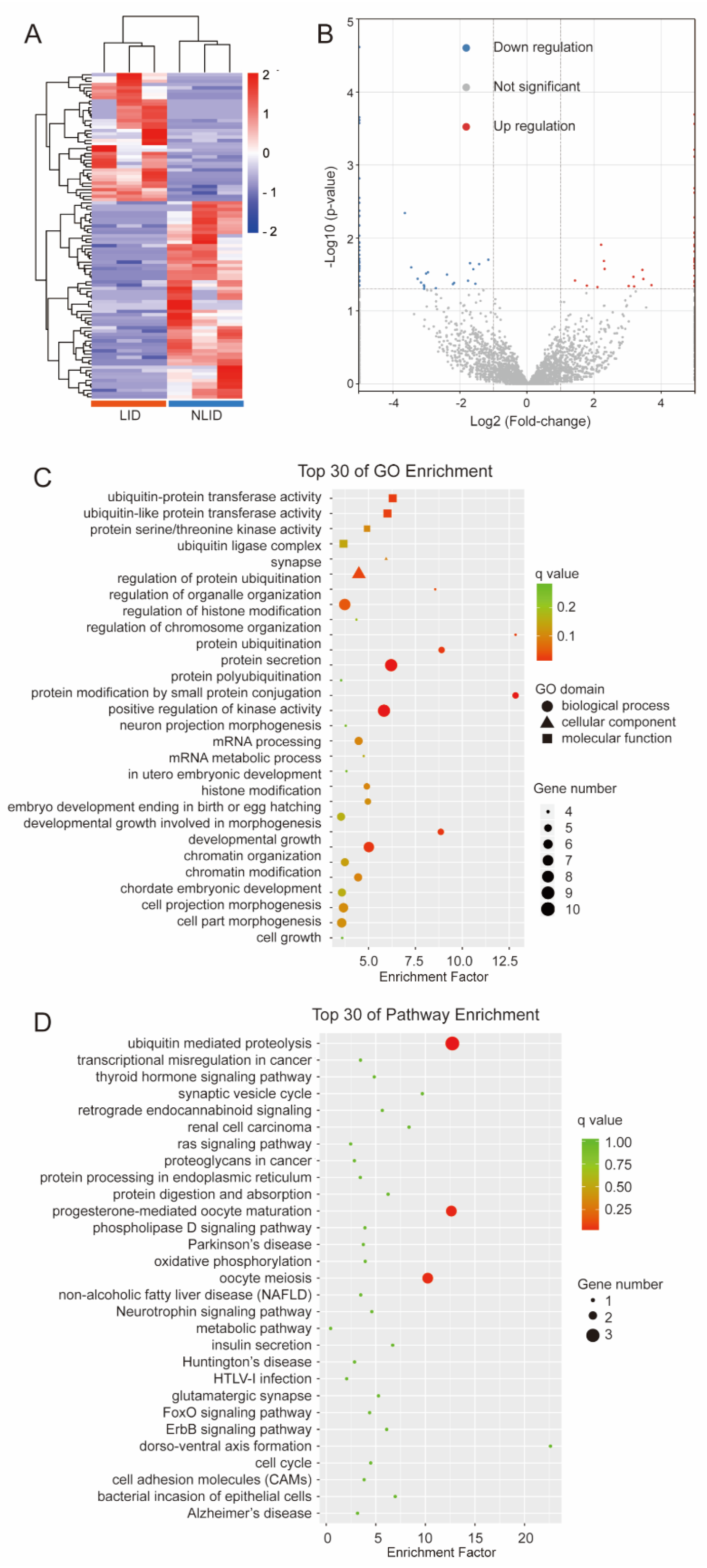
3.2. circRNAs Expression Profiles of LID and NLID Rats
3.3. Functional Annotation of the Host Genes of DEcircRNAs
3.4. Rno-Rsf1_0012 Expression Validated by qRT-PCR
3.5. Rno-Rsf1_0012 Regulates Expression of Target Genes via rno-miR-298-5p
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- McGregor, M.M.; Nelson, A.B. Circuit Mechanisms of Parkinson’s Disease. Neuron 2019, 101, 1042–1056. [Google Scholar] [CrossRef] [PubMed]
- Dexter, D.T.; Jenner, P. Parkinson disease: From pathology to molecular disease mechanisms. Free Radic. Biol. Med. 2013, 62, 132–144. [Google Scholar] [CrossRef] [PubMed]
- Ahlskog, J.E.; Muenter, M.D. Frequency of levodopa-related dyskinesias and motor fluctuations as estimated from the cumulative literature. Mov. Disord. 2001, 16, 448–458. [Google Scholar] [CrossRef]
- Scott, N.W.; Macleod, A.D.; Counsell, C.E. Motor complications in an incident Parkinson’s disease cohort. Eur. J. Neurol. 2016, 23, 304–312. [Google Scholar] [CrossRef] [PubMed]
- Jenner, P. Preventing and controlling dyskinesia in Parkinson’s disease—A view of current knowledge and future opportunities. Mov. Disord. 2008, 23 (Suppl. 3), S585–S598. [Google Scholar] [CrossRef]
- Thanvi, B.; Lo, N.; Robinson, T. Levodopa-induced dyskinesia in Parkinson’s disease: Clinical features, pathogenesis, prevention and treatment. Postgrad. Med. J. 2007, 83, 384–388. [Google Scholar] [CrossRef]
- Li, X.; Yang, L.; Chen, L.L. The Biogenesis, Functions, and Challenges of Circular RNAs. Mol. Cell 2018, 71, 428–442. [Google Scholar] [CrossRef]
- Kristensen, L.S.; Andersen, M.S.; Stagsted, L.V.W.; Ebbesen, K.K.; Hansen, T.B.; Kjems, J. The biogenesis, biology and characterization of circular RNAs. Nat. Rev. Genet. 2019, 20, 675–691. [Google Scholar] [CrossRef]
- Gasparini, S.; Licursi, V.; Presutti, C.; Mannironi, C. The Secret Garden of Neuronal circRNAs. Cells 2020, 9, 1815. [Google Scholar] [CrossRef]
- Liu, J.; Zhao, K.; Huang, N.; Zhang, N. Circular RNAs and human glioma. Cancer Biol. Med. 2019, 16, 11–23. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.L.; Su, M.; Wu, D.P. Functional roles of circular RNAs in Alzheimer’s disease. Ageing Res. Rev. 2020, 60, 101058. [Google Scholar] [CrossRef] [PubMed]
- Hanan, M.; Simchovitz, A.; Yayon, N.; Vaknine, S.; Cohen-Fultheim, R.; Karmon, M.; Madrer, N.; Rohrlich, T.M.; Maman, M.; Bennett, E.R.; et al. A Parkinson’s disease CircRNAs Resource reveals a link between circSLC8A1 and oxidative stress. EMBO Mol. Med. 2020, 12, e11942. [Google Scholar] [CrossRef]
- Barros, A.S.; Crispim, R.Y.G.; Cavalcanti, J.U.; Souza, R.B.; Lemos, J.C.; Cristino Filho, G.; Bezerra, M.M.; Pinheiro, T.F.M.; de Vasconcelos, S.M.M.; Macêdo, D.S.; et al. Impact of the Chronic Omega-3 Fatty Acids Supplementation in Hemiparkinsonism Model Induced by 6-Hydroxydopamine in Rats. Basic Clin. Pharmacol. Toxicol. 2017, 120, 523–531. [Google Scholar] [CrossRef] [PubMed]
- Steece-Collier, K.; Stancati, J.A.; Collier, N.J.; Sandoval, I.M.; Mercado, N.M.; Sortwell, C.E.; Collier, T.J.; Manfredsson, F.P. Genetic silencing of striatal CaV1.3 prevents and ameliorates levodopa dyskinesia. Mov. Disord. 2019, 34, 697–707. [Google Scholar] [CrossRef]
- Lundblad, M.; Andersson, M.; Winkler, C.; Kirik, D.; Wierup, N.; Cenci, M.A. Pharmacological validation of behavioural measures of akinesia and dyskinesia in a rat model of Parkinson’s disease. Eur. J. Neurosci. 2002, 15, 120–132. [Google Scholar] [CrossRef]
- Li, H. Aligning sequence reads, clone sequences and assembly contigs with BWA-MEM. arXiv preprint 2013, arXiv:1303.3997. [Google Scholar]
- Gao, Y.; Wang, J.; Zhao, F. CIRI: An efficient and unbiased algorithm for de novo circular RNA identification. Genome Biol. 2015, 16, 4. [Google Scholar] [CrossRef] [PubMed]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef]
- Kumar, S.; Reddy, P.H. Are circulating microRNAs peripheral biomarkers for Alzheimer’s disease? Biochim. Biophys. Acta 2016, 1862, 1617–1627. [Google Scholar] [CrossRef]
- Mao, X.; Cai, T.; Olyarchuk, J.G.; Wei, L. Automated genome annotation and pathway identification using the KEGG Orthology (KO) as a controlled vocabulary. Bioinformatics 2005, 21, 3787–3793. [Google Scholar] [CrossRef]
- Miranda, K.C.; Huynh, T.; Tay, Y.; Ang, Y.S.; Tam, W.L.; Thomson, A.M.; Lim, B.; Rigoutsos, I. A pattern-based method for the identification of MicroRNA binding sites and their corresponding heteroduplexes. Cell 2006, 126, 1203–1217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Wang, X. miRDB: An online database for prediction of functional microRNA targets. Nucleic Acids Res. 2020, 48, D127–D131. [Google Scholar] [CrossRef] [PubMed]
- Jenner, P. Molecular mechanisms of L-DOPA-induced dyskinesia. Nat. Rev. Neurosci. 2008, 9, 665–677. [Google Scholar] [CrossRef] [PubMed]
- Berton, O.; Guigoni, C.; Li, Q.; Bioulac, B.H.; Aubert, I.; Gross, C.E.; Dileone, R.J.; Nestler, E.J.; Bezard, E. Striatal overexpression of DeltaJunD resets L-DOPA-induced dyskinesia in a primate model of Parkinson disease. Biol. Psychiatry 2009, 66, 554–561. [Google Scholar] [CrossRef]
- Pavon, N.; Martin, A.B.; Mendialdua, A.; Moratalla, R. ERK phosphorylation and FosB expression are associated with L-DOPA-induced dyskinesia in hemiparkinsonian mice. Biol. Psychiatry 2006, 59, 64–74. [Google Scholar] [CrossRef]
- Schuster, S.; Nadjar, A.; Guo, J.T.; Li, Q.; Ittrich, C.; Hengerer, B.; Bezard, E. The 3-hydroxy-3-methylglutaryl-CoA reductase inhibitor lovastatin reduces severity of L-DOPA-induced abnormal involuntary movements in experimental Parkinson’s disease. J. Neurosci. 2008, 28, 4311–4316. [Google Scholar] [CrossRef]
- Han, C.L.; Liu, Y.P.; Sui, Y.P.; Chen, N.; Du, T.T.; Jiang, Y.; Guo, C.J.; Wang, K.L.; Wang, Q.; Fan, S.Y.; et al. Integrated transcriptome expression profiling reveals a novel lncRNA associated with L-DOPA-induced dyskinesia in a rat model of Parkinson’s disease. Aging 2020, 12, 718–739. [Google Scholar] [CrossRef]
- Tay, Y.; Rinn, J.; Pandolfi, P.P. The multilayered complexity of ceRNA crosstalk and competition. Nature 2014, 505, 344–352. [Google Scholar] [CrossRef]
- Konradi, C.; Westin, J.E.; Carta, M.; Eaton, M.E.; Kuter, K.; Dekundy, A.; Lundblad, M.; Cenci, M.A. Transcriptome analysis in a rat model of L-DOPA-induced dyskinesia. Neurobiol. Dis. 2004, 17, 219–236. [Google Scholar] [CrossRef]
- Zhang, Y.; Meredith, G.E.; Mendoza-Elias, N.; Rademacher, D.J.; Tseng, K.Y.; Steece-Collier, K. Aberrant restoration of spines and their synapses in L-DOPA-induced dyskinesia: Involvement of corticostriatal but not thalamostriatal synapses. J. Neurosci. 2013, 33, 11655–11667. [Google Scholar] [CrossRef]
- Espay, A.J.; Morgante, F.; Merola, A.; Fasano, A.; Marsili, L.; Fox, S.H.; Bezard, E.; Picconi, B.; Calabresi, P.; Lang, A.E. Levodopa-induced dyskinesia in Parkinson disease: Current and evolving concepts. Ann. Neurol. 2018, 84, 797–811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cenci, M.A.; Lee, C.S.; Björklund, A. L-DOPA-induced dyskinesia in the rat is associated with striatal overexpression of prodynorphin- and glutamic acid decarboxylase mRNA. Eur. J. Neurosci. 1998, 10, 2694–2706. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.S.; Cenci, M.A.; Schulzer, M.; Björklund, A. Embryonic ventral mesencephalic grafts improve levodopa-induced dyskinesia in a rat model of Parkinson’s disease. Brain 2000, 123 Pt 7, 1365–1379. [Google Scholar] [CrossRef]
- Steece-Collier, K.; Collier, T.J.; Lipton, J.W.; Stancati, J.A.; Winn, M.E.; Cole-Strauss, A.; Sellnow, R.; Conti, M.M.; Mercado, N.M.; Nillni, E.A.; et al. Striatal Nurr1, but not FosB expression links a levodopa-induced dyskinesia phenotype to genotype in Fisher 344 vs. Lewis hemiparkinsonian rats. Exp. Neurol. 2020, 330, 113327. [Google Scholar] [CrossRef] [PubMed]
- Sellnow, R.C.; Steece-Collier, K.; Altwal, F.; Sandoval, I.M.; Kordower, J.H.; Collier, T.J.; Sortwell, C.E.; West, A.R.; Manfredsson, F.P. Striatal Nurr1 Facilitates the Dyskinetic State and Exacerbates Levodopa-Induced Dyskinesia in a Rat Model of Parkinson’s Disease. J. Neurosci. 2020, 40, 3675–3691. [Google Scholar] [CrossRef]
- Lindgren, H.S.; Rylander, D.; Ohlin, K.E.; Lundblad, M.; Cenci, M.A. The “motor complication syndrome” in rats with 6-OHDA lesions treated chronically with L-DOPA: Relation to dose and route of administration. Behav. Brain Res. 2007, 177, 150–159. [Google Scholar] [CrossRef]
- Santini, E.; Alcacer, C.; Cacciatore, S.; Heiman, M.; Herve, D.; Greengard, P.; Girault, J.A.; Valjent, E.; Fisone, G. L-DOPA activates ERK signaling and phosphorylates histone H3 in the striatonigral medium spiny neurons of hemiparkinsonian mice. J. Neurochem. 2009, 108, 621–633. [Google Scholar] [CrossRef]
- Olanow, C.W. The pathogenesis of cell death in Parkinson’s disease--2007. Mov. Disord. 2007, 22 (Suppl. 17), S335–S342. [Google Scholar] [CrossRef]
- Sang, Q.; Liu, X.; Wang, L.; Qi, L.; Sun, W.; Wang, W.; Sun, Y.; Zhang, H. CircSNCA downregulation by pramipexole treatment mediates cell apoptosis and autophagy in Parkinson’s disease by targeting miR-7. Aging 2018, 10, 1281–1293. [Google Scholar] [CrossRef]
- Kumar, L.; Shamsuzzama; Jadiya, P.; Haque, R.; Shukla, S.; Nazir, A. Functional Characterization of Novel Circular RNA Molecule, circzip-2 and Its Synthesizing Gene zip-2 in C. elegans Model of Parkinson’s Disease. Mol. Neurobiol. 2018, 55, 6914–6926. [Google Scholar] [CrossRef]
- Surguchov, A. Intracellular Dynamics of Synucleins: “Here, There and Everywhere”. Int. Rev. Cell Mol. Biol. 2015, 320, 103–169. [Google Scholar] [CrossRef] [PubMed]
- Utal, A.K.; Stopka, A.L.; Roy, M.; Coleman, P.D. PEP-19 immunohistochemistry defines the basal ganglia and associated structures in the adult human brain, and is dramatically reduced in Huntington’s disease. Neuroscience 1998, 86, 1055–1063. [Google Scholar] [CrossRef]
- Scoles, D.R.; Meera, P.; Schneider, M.D.; Paul, S.; Dansithong, W.; Figueroa, K.P.; Hung, G.; Rigo, F.; Bennett, C.F.; Otis, T.S.; et al. Antisense oligonucleotide therapy for spinocerebellar ataxia type 2. Nature 2017, 544, 362–366. [Google Scholar] [CrossRef]
- Dansithong, W.; Paul, S.; Figueroa, K.P.; Rinehart, M.D.; Wiest, S.; Pflieger, L.T.; Scoles, D.R.; Pulst, S.M. Ataxin-2 regulates RGS8 translation in a new BAC-SCA2 transgenic mouse model. PLoS Genet. 2015, 11, e1005182. [Google Scholar] [CrossRef]
- Liu, Q.; Pan, Y.; Li, X.J.; Li, S. Molecular Mechanisms and Therapeutics for SCA17. Neurotherapeutics 2019, 16, 1097–1105. [Google Scholar] [CrossRef] [PubMed]
- Coutelier, M.; Coarelli, G.; Monin, M.L.; Konop, J.; Davoine, C.S.; Tesson, C.; Valter, R.; Anheim, M.; Behin, A.; Castelnovo, G.; et al. A panel study on patients with dominant cerebellar ataxia highlights the frequency of channelopathies. Brain 2017, 140, 1579–1594. [Google Scholar] [CrossRef]
- Stevanin, G.; Fujigasaki, H.; Lebre, A.S.; Camuzat, A.; Jeannequin, C.; Dode, C.; Takahashi, J.; San, C.; Bellance, R.; Brice, A.; et al. Huntington’s disease-like phenotype due to trinucleotide repeat expansions in the TBP and JPH3 genes. Brain 2003, 126, 1599–1603. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ID | Sequence of Primers | |
|---|---|---|
| rno-Rsf1_0012 | Forward | 5′-GCCTTCCGAATCACCCAGAA-3′ |
| Reverse | 5′-GAATCCATTGACCGCTCATCAG-3′ | |
| rno-Rims2_0060 | Forward | 5′-GGCTCACAAGACAGGATTCTATT-3′ |
| Reverse | 5′-GCTTTCTGTCTGAAGGCATGT-3′ | |
| rno-N4bp1_0001 | Forward | 5′-GCCATTACGAGTACATCAAAGGG-3′ |
| Reverse | 5′-AACACAGAGGTCAGCACAAGTA-3′ | |
| rno-Ick_0003 | Forward | 5′-AAGGACTGGCGTTCATTCACA-3′ |
| Reverse | 5′-GATGGCAGCACCAGCACAA-3′ | |
| rno-Stk39_0001 | Forward | 5′-GCTCTTCTCTGCTGGCTTGG-3′ |
| Reverse | 5′-GGCTTACCTTGGCTTTCTGGAA-3′ | |
| rno-Ell2_0005 | Forward | 5′-GGTGGGTGCTTGTTAAGTATATTAC-3′ |
| Reverse | 5′-GCTGCTTGATCTTCTGATATTCTTG-3′ | |
| PCP4 | Forward | 5′-CTCACTGCCAGAGGAGGAATG-3′ |
| Reverse | 5′-AATTCTTCTTGGACCTTCTTCTGC-3′ | |
| TBP | Forward | 5′-CTTCAGTCCAATGATGCCTTACG-3′ |
| Reverse | 5′-CTGCTGCTGCTGCTGTCTT-3′ | |
| ID | Type | Fold Change | Regulation |
|---|---|---|---|
| rno-Ick_0003 | exon | 13.13123 | UP |
| rno-N4bp1_0001 | exon | 11.10113 | UP |
| rno-Ell2_0005 | exon | 10.88539 | UP |
| rno-Rims2_0060 | exon | 9.183134 | UP |
| rno-Stk39_0001 | exon | 9.045527 | UP |
| rno-Rsf1_0012 | exon | 8.201328 | UP |
| rno-Chd2_0001 | exon | 4.98735 | UP |
| rno-Trip12_0024 | exon | 4.913605 | UP |
| rno-Arl8b_0001 | exon | 4.636859 | UP |
| rno-Dmd_0004 | exon | 4.296785 | UP |
| rno-Ralgps2_0004 | exon | 6.565114 | DOWN |
| rno-Susd1_0002 | exon | 7.722112 | DOWN |
| circRNA.15164 | intergenic region | 8.034623 | DOWN |
| rno-Sergef_0005 | exon | 8.351556 | DOWN |
| circRNA.4818 | exon | 8.367079 | DOWN |
| rno-Kdm4c_0013 | exon | 8.40449 | DOWN |
| rno-Pcsk5_0002 | exon | 8.931029 | DOWN |
| rno-Prex2_0027 | exon | 9.574243 | DOWN |
| rno-Rps6ka5_0004 | exon | 10.93379 | DOWN |
| rno-Slc16a10_0001 | exon | 12.47348 | DOWN |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Han, C.-L.; Wang, Q.; Liu, C.; Li, Z.-B.; Du, T.-T.; Sui, Y.-P.; Zhang, X.; Zhang, J.-G.; Xiao, Y.-L.; Cai, G.-E.; et al. Transcriptome Sequencing Reveal That Rno-Rsf1_0012 Participates in Levodopa-Induced Dyskinesia in Parkinson’s Disease Rats via Binding to Rno-mir-298-5p. Brain Sci. 2022, 12, 1206. https://doi.org/10.3390/brainsci12091206
Han C-L, Wang Q, Liu C, Li Z-B, Du T-T, Sui Y-P, Zhang X, Zhang J-G, Xiao Y-L, Cai G-E, et al. Transcriptome Sequencing Reveal That Rno-Rsf1_0012 Participates in Levodopa-Induced Dyskinesia in Parkinson’s Disease Rats via Binding to Rno-mir-298-5p. Brain Sciences. 2022; 12(9):1206. https://doi.org/10.3390/brainsci12091206
Chicago/Turabian StyleHan, Chun-Lei, Qiao Wang, Chong Liu, Zhi-Bao Li, Ting-Ting Du, Yun-Peng Sui, Xin Zhang, Jian-Guo Zhang, Yi-Lei Xiao, Guo-En Cai, and et al. 2022. "Transcriptome Sequencing Reveal That Rno-Rsf1_0012 Participates in Levodopa-Induced Dyskinesia in Parkinson’s Disease Rats via Binding to Rno-mir-298-5p" Brain Sciences 12, no. 9: 1206. https://doi.org/10.3390/brainsci12091206