

Identification of Adipocytokine Pathway-Related Genes in Epilepsy and Its Effect on the Peripheral Immune Landscape
Abstract
:1. Introduction
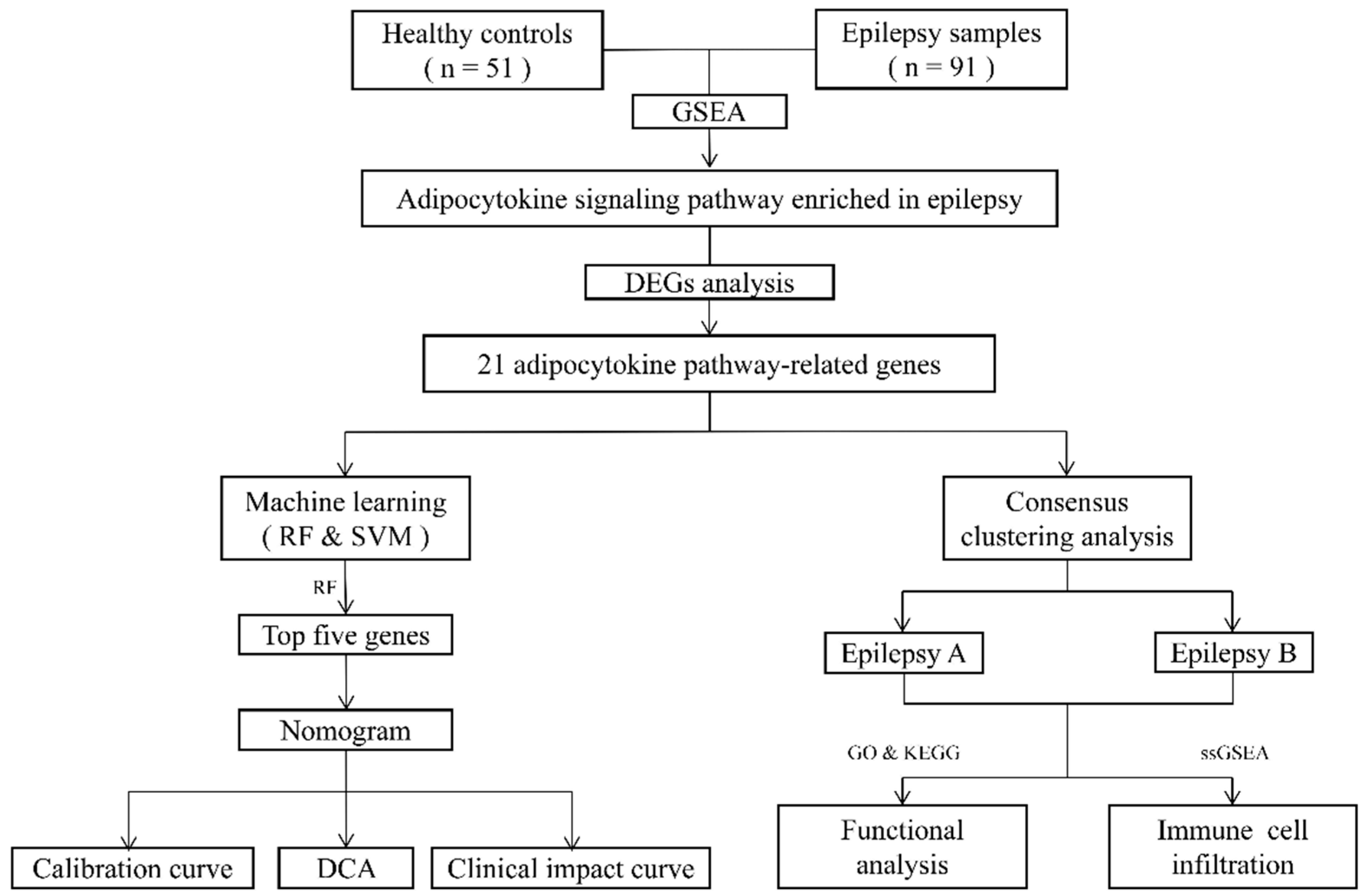
2. Materials and Methods
2.1. Data Preparation
2.2. Gene Set Enrichment Analysis
2.3. Difference Analysis of Adipocytokine Pathway-Related Genes
2.4. Selection of Disease-Related Genes
2.5. Construction and Evaluation of a Nomogram
2.6. Consensus Clustering of 21 Adipocytokine Pathway-Related Genes
2.7. Identification of Immune Microenvironment
2.8. Functional Enrichment Analysis of Distinct Adipocytokine Signal Patterns
3. Results
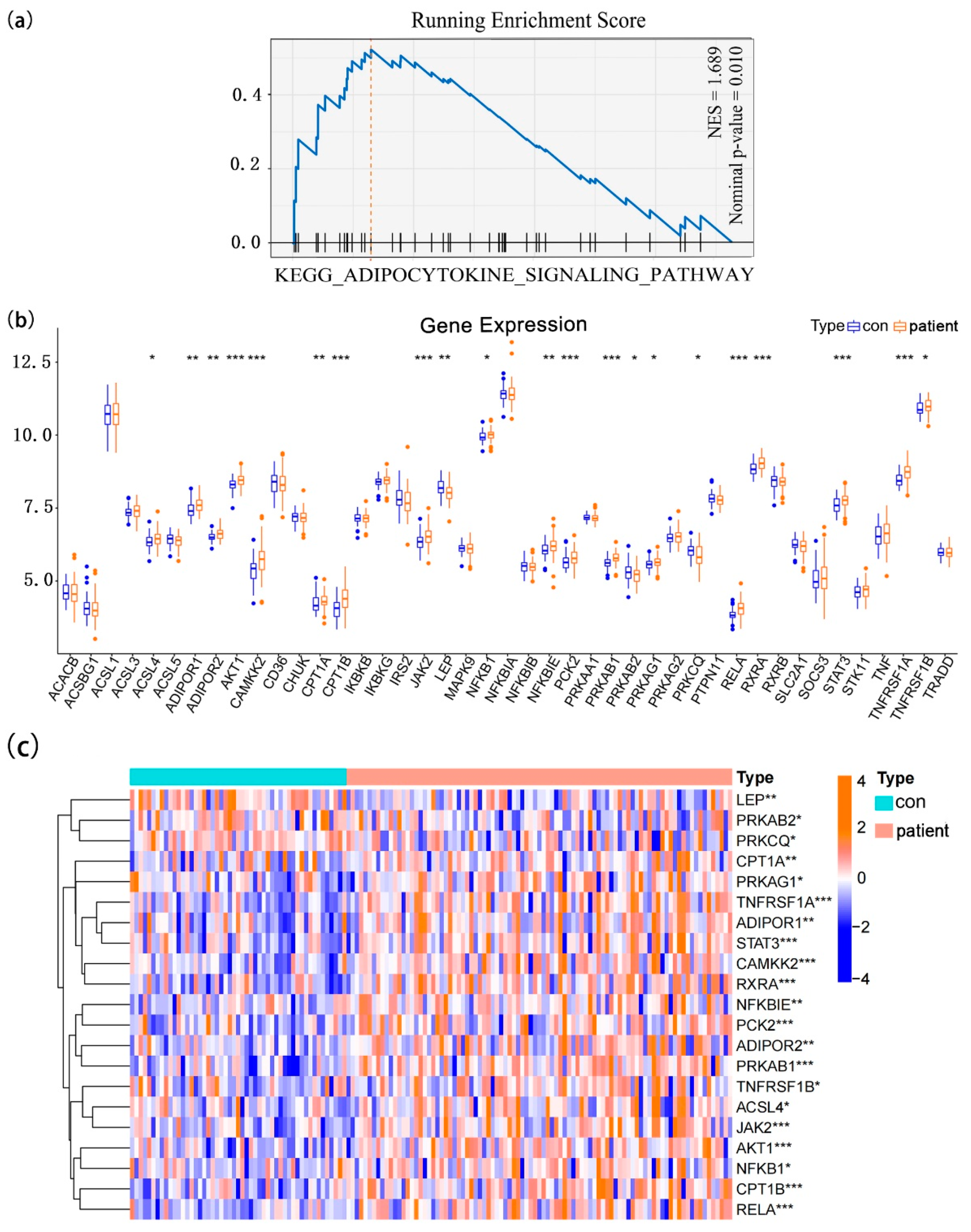
3.1. The Landscape of Adipocytokine Pathway-Related Genes in Normal and Epilepsy Samples
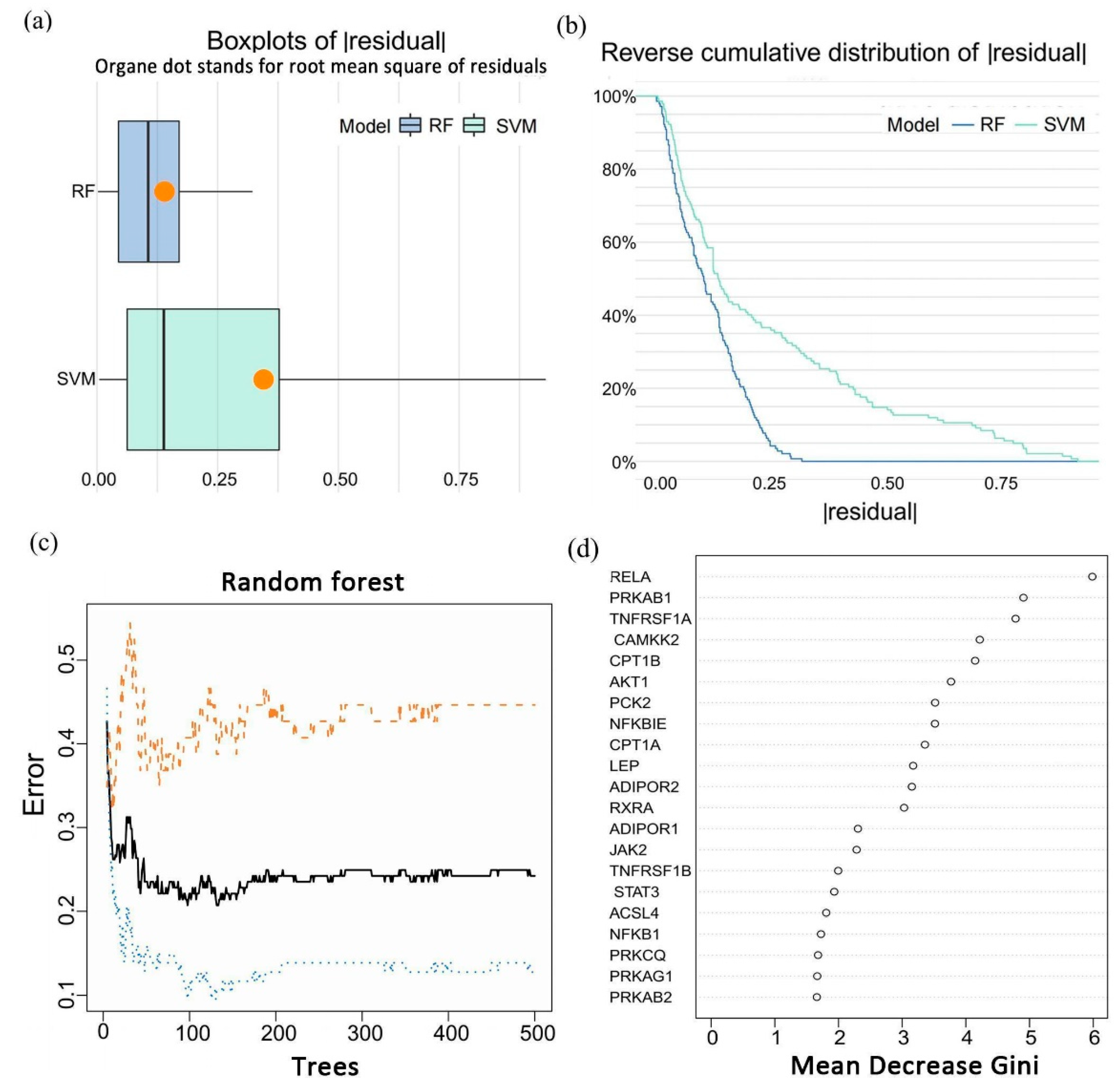
3.2. Identification of a Five-Gene Signature for Epilepsy
3.3. Construction and Evaluation of a Diagnostic Nomogram
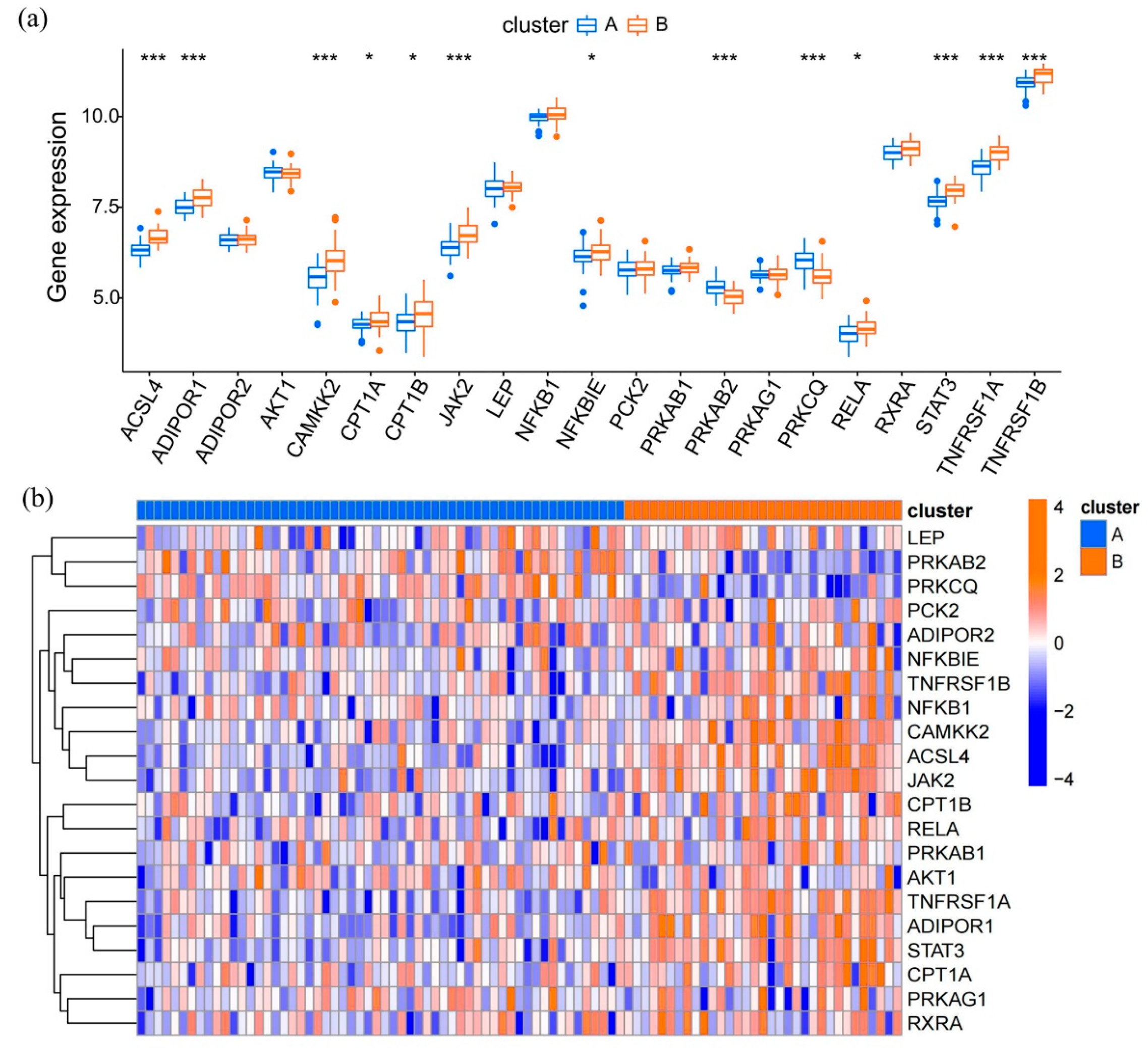
3.4. Expression Patterns of Adipocytokine Pathway-Related Genes in Epilepsy
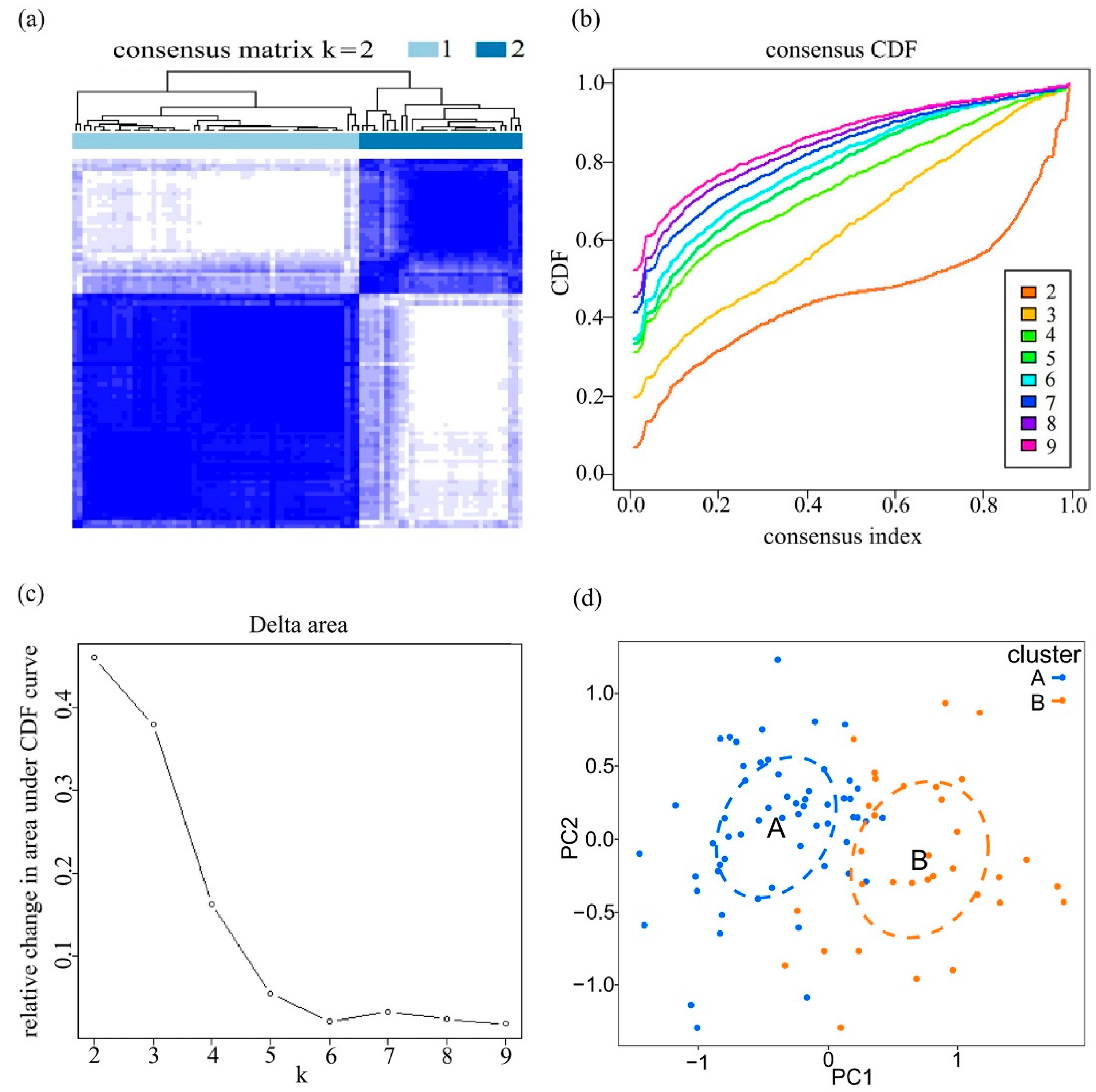
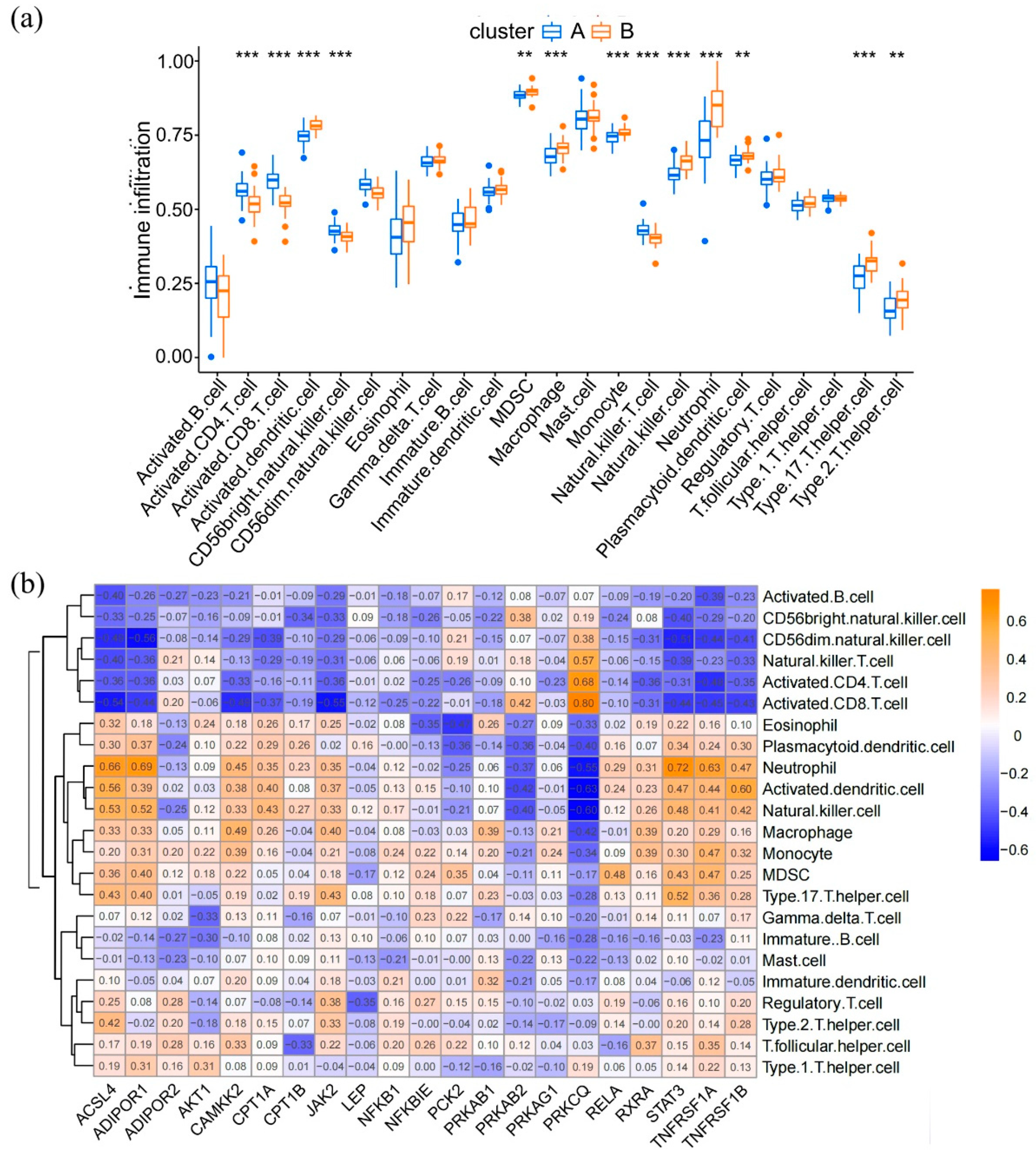
3.5. Immune Landscapes in Distinct Adipocytokine Signal Expression Patterns
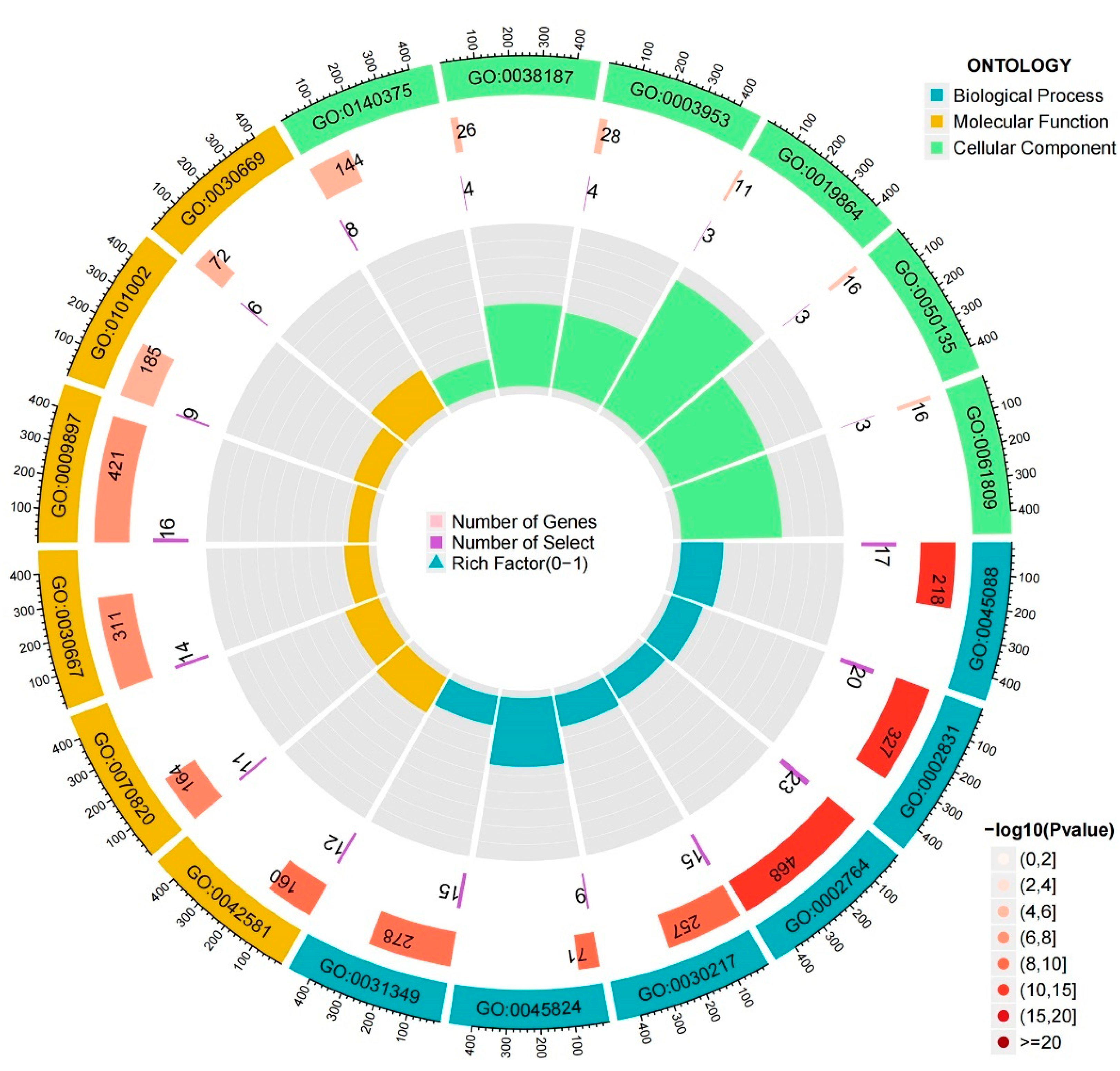
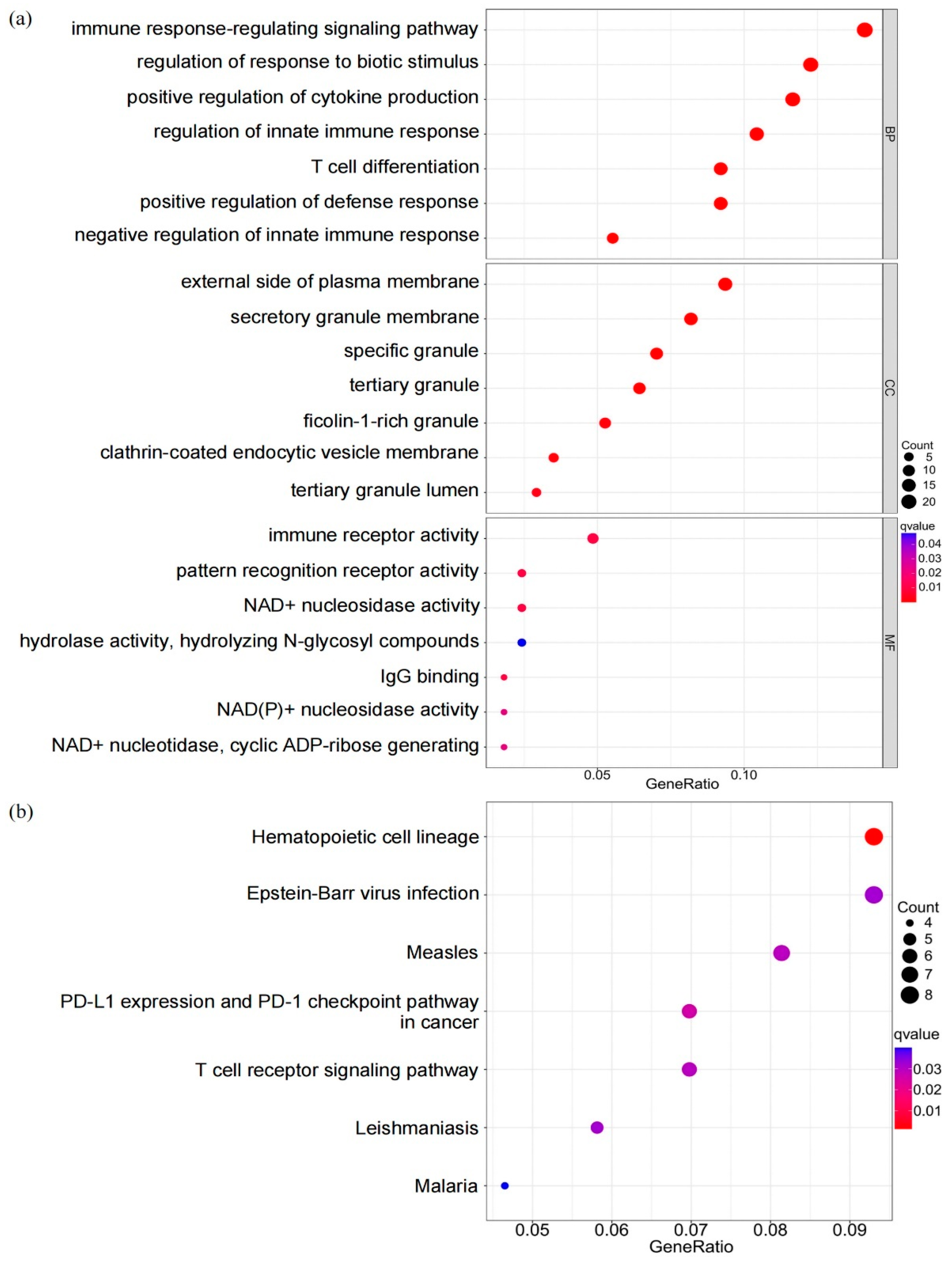
3.6. Biological Characteristics of DEGs in Different Patterns
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Thijs, R.D.; Surges, R.; O’Brien, T.J.; Sander, J.W. Epilepsy in adults. Lancet 2019, 393, 689–701. [Google Scholar] [CrossRef]
- Scheffer, I.E.; Berkovic, S.; Capovilla, G.; Connolly, M.B.; French, J.; Guilhoto, L.; Hirsch, E.; Jain, S.; Mathern, G.W.; Moshe, S.L.; et al. ILAE classification of the epilepsies: Position paper of the ILAE Commission for Classification and Terminology. Epilepsia 2017, 58, 512–521. [Google Scholar] [CrossRef] [PubMed]
- Falco-Walter, J. Epilepsy-Definition, Classification, Pathophysiology, and Epidemiology. Semin. Neurol. 2020, 40, 617–623. [Google Scholar] [CrossRef] [PubMed]
- Kanner, A.M.; Bicchi, M.M. Antiseizure Medications for Adults with Epilepsy: A Review. JAMA 2022, 327, 1269–1281. [Google Scholar] [CrossRef] [PubMed]
- Amanat, M.; Thijs, R.D.; Salehi, M.; Sander, J.W. Seizures as a clinical manifestation in somatic autoimmune disorders. Seizure 2019, 64, 59–64. [Google Scholar] [CrossRef]
- Ong, M.-S.; Kohane, I.S.; Cai, T.; Gorman, M.P.; Mandl, K.D. Population-level evidence for an autoimmune etiology of epilepsy. JAMA Neurol. 2014, 71, 569–574. [Google Scholar] [CrossRef]
- Vezzani, A.; Balosso, S.; Ravizza, T. Neuroinflammatory pathways as treatment targets and biomarkers in epilepsy. Nat. Rev. Neurol. 2019, 15, 459–472. [Google Scholar] [CrossRef]
- Stolarczyk, E. Adipose tissue inflammation in obesity: A metabolic or immune response? Curr. Opin. Pharmacol. 2017, 37, 35–40. [Google Scholar] [CrossRef]
- Fantuzzi, G. Adipose tissue, adipokines, and inflammation. J. Allergy Clin. Immunol. 2005, 115, 911–919. [Google Scholar] [CrossRef]
- Tilg, H.; Moschen, A.R. Adipocytokines: Mediators linking adipose tissue, inflammation and immunity. Nat. Rev. Immunol. 2006, 6, 772–783. [Google Scholar] [CrossRef]
- Palmio, J.; Vuolteenaho, K.; Lehtimäki, K.; Nieminen, R.; Peltola, J.; Moilanen, E. CSF and plasma adipokines after tonic–clonic seizures. Seizure 2016, 39, 10–12. [Google Scholar] [CrossRef]
- Xu, L.; Rensing, N.; Yang, X.-F.; Zhang, H.X.; Thio, L.L.; Rothman, S.M.; Weisenfeld, A.E.; Wong, M.; Yamada, K.A. Leptin inhibits 4-aminopyridine– and pentylenetetrazole-induced seizures and AMPAR-mediated synaptic transmission in rodents. J. Clin. Investig. 2008, 118, 272–280. [Google Scholar] [CrossRef] [PubMed]
- Chyra, M.; Roczniak, W.; Świętochowska, E.; Dudzińska, M.; Oświęcimska, J. The Effect of the Ketogenic Diet on Adiponectin, Omentin and Vaspin in Children with Drug-Resistant Epilepsy. Nutrients 2022, 14, 479. [Google Scholar] [CrossRef]
- Ritchie, M.E.; Belinda, P.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res. 2015, 43, e47. [Google Scholar] [CrossRef] [PubMed]
- Charoentong, P.; Finotello, F.; Angelova, M.; Mayer, C.; Efremova, M.; Rieder, D.; Hackl, H.; Trajanoski, Z. Pan-cancer Immunogenomic Analyses Reveal Genotype-Immunophenotype Relationships and Predictors of Response to Checkpoint Blockade. Cell Rep. 2017, 18, 248–262. [Google Scholar] [CrossRef] [PubMed]
- Rawat, C.; Kushwaha, S.; Srivastava, A.K.; Kukreti, R. Peripheral blood gene expression signatures associated with epilepsy and its etiologic classification. Genomics 2020, 112, 218–224. [Google Scholar] [CrossRef]
- Rawat, C.; Kutum, R.; Kukal, S.; Srivastava, A.; Dahiya, U.R.; Kushwaha, S.; Sharma, S.; Dash, D.; Saso, L.; Srivastava, A.K.; et al. Downregulation of peripheral PTGS2/COX-2 in response to valproate treatment in patients with epilepsy. Sci. Rep. 2020, 10, 2546. [Google Scholar] [CrossRef] [PubMed]
- Singh, G.; Sander, J.W. The global burden of epilepsy report: Implications for low- and middle-income countries. Epilepsy Behav. 2020, 105, 106949. [Google Scholar] [CrossRef]
- Bauer, J.; Becker, A.J.; Elyaman, W.; Peltola, J.; Rüegg, S.; Titulaer, M.J.; Varley, J.A.; Beghi, E. Innate and adaptive immunity in human epilepsies. Epilepsia 2017, 58 (Suppl. S3), 57–68. [Google Scholar] [CrossRef] [PubMed]
- Hamed, S.A. Leptin and insulin homeostasis in epilepsy: Relation to weight adverse conditions. Epilepsy Res. 2007, 75, 1–9. [Google Scholar] [CrossRef]
- Shanley, L.J.; O’Malley, D.; Irving, A.J.; Ashford, M.L.; Harvey, J. Leptin inhibits epileptiform-like activity in rat hippocampal neurones via PI 3-kinase-driven activation of BK channels. J. Physiol. 2002, 545, 933–944. [Google Scholar] [CrossRef] [PubMed]
- Fang, H.; Judd, R.L. Adiponectin Regulation and Function. Compr. Physiol. 2018, 8, 1031–1063. [Google Scholar] [PubMed]
- Schlaepfer, I.R.; Joshi, M. CPT1A-mediated Fat Oxidation, Mechanisms, and Therapeutic Potential. Endocrinology 2020, 161, bqz046. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Lenardo, M.J.; Baltimore, D. 30 Years of NF-kappaB: A Blossoming of Relevance to Human Pathobiology. Cell 2017, 168, 37–57. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Hu, X.; Hu, X.; Gao, F.; Li, M.; Cui, Y.; Wei, X.; Qin, Y.; Zhang, C.; Zhao, Y.; et al. MicroRNA-520c-3p targeting of RelA/p65 suppresses atherosclerotic plaque formation. Int. J. Biochem. Cell Biol. 2021, 131, 105873. [Google Scholar] [CrossRef]
- Chen, S.; Jiang, S.; Zheng, W.; Tu, B.; Liu, S.; Ruan, H.; Fan, C. RelA/p65 inhibition prevents tendon adhesion by modulating inflammation, cell proliferation, and apoptosis. Cell Death Dis. 2017, 8, e2710. [Google Scholar] [CrossRef]
- Pinkosky, S.L.; Scott, J.W.; Desjardins, E.M.; Smith, B.K.; Day, E.A.; Ford, R.J.; Langendorf, C.G.; Ling, N.X.Y.; Nero, T.L.; Loh, K.; et al. Long-chain fatty acyl-CoA esters regulate metabolism via allosteric control of AMPK beta1 isoforms. Nat. Metab. 2020, 2, 873–881. [Google Scholar] [CrossRef]
- Ni, Y.; Xu, Z.; Li, C.; Zhu, Y.; Liu, R.; Zhang, F.; Chang, H.; Li, M.; Sheng, L.; Li, Z.; et al. Therapeutic inhibition of miR-802 protects against obesity through AMPK-mediated regulation of hepatic lipid metabolism. Theranostics 2021, 11, 1079–1099. [Google Scholar] [CrossRef]
- Peng, I.C.; Chen, Z.; Sun, W.; Li, Y.S.; Marin, T.L.; Hsu, P.H.; Su, M.I.; Cui, X.; Pan, S.; Lytle, C.Y.; et al. Glucagon regulates ACC activity in adipocytes through the CAMKKbeta/AMPK pathway. Am. J. Physiol. Endocrinol. Metab. 2012, 302, E1560–E1568. [Google Scholar] [CrossRef]
- Galon, J.; Aksentijevich, I.; McDermott, M.F.; O’Shea, J.J.; Kastner, D.L. TNFRSF1A mutations and autoinflammatory syndromes. Curr. Opin. Immunol. 2000, 12, 479–486. [Google Scholar] [CrossRef]
- Tienari, P.J.; Hohlfeld, R. TNFRSF1A in multiple sclerosis: A tale of soluble receptors and signaling cascades. Neurology 2013, 81, 1886–1888. [Google Scholar] [CrossRef] [PubMed]
- Warfel, J.D.; Bermudez, E.M.; Mendoza, T.M.; Ghosh, S.; Zhang, J.; Elks, C.M.; Mynatt, R.; Vandanmagsar, B. Mitochondrial fat oxidation is essential for lipid-induced inflammation in skeletal muscle in mice. Sci. Rep. 2016, 6, 37941. [Google Scholar] [PubMed]
- De Heredia, F.P.; Gómez-Martínez, S.; Marcos, A. Obesity, inflammation and the immune system. Proc. Nutr. Soc. 2012, 71, 332–338. [Google Scholar] [PubMed] [Green Version]
- Feng, L.; Murugan, M.; Bosco, D.B.; Liu, Y.; Peng, J.; Worrell, G.A.; Wang, H.; Ta, L.E.; Richardson, J.; Shen, Y.; et al. Microglial proliferation and monocyte infiltration contribute to microgliosis following status epilepticus. Glia 2019, 67, 1434–1448. [Google Scholar]
- Varvel, N.H.; Neher, J.J.; Bosch, A.; Wang, W.; Ransohoff, R.M.; Miller, R.J.; Dingledine, R. Infiltrating monocytes promote brain inflammation and exacerbate neuronal damage after status epilepticus. Proc. Natl. Acad. Sci. USA 2016, 113, E5665–E5674. [Google Scholar]
- Alvarado, C.D.; Brewster, A.L. Hit by a Smooth CD8: T-Cell Attack on Hippocampal Neurons Triggers Limbic Encephalitis and Epilepsy. Epilepsy Curr. 2021, 21, 369–371. [Google Scholar]
- Helmstaedter, C.; Hansen, N.; Leelaarporn, P.; Schwing, K.; Oender, D.; Widman, G.; Racz, A.; Surges, R.; Becker, A.; Witt, J.-A. Specific B- and T-cell populations are associated with cognition in patients with epilepsy and antibody positive and negative suspected limbic encephalitis. J. Neurol. 2021, 268, 455–466. [Google Scholar]
- Granata, T.; Cross, H.; Theodore, W.; Avanzini, G. Immune-mediated epilepsies. Epilepsia 2011, 52 (Suppl. S3), 5–11. [Google Scholar]
- Tröscher, A.R.; Sakaraki, E.; Mair, K.M.; Köck, U.; Racz, A.; Borger, V.; Cloppenborg, T.; Becker, A.J.; Bien, C.G.; Bauer, J. T cell numbers correlate with neuronal loss rather than with seizure activity in medial temporal lobe epilepsy. Epilepsia 2021, 62, 1343–1353. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ID | Categories | Description |
| GO:0140375 | CC | immune receptor activity |
| GO:0038187 | CC | pattern recognition receptor activity |
| GO:0003953 | CC | NAD+ nucleosidase activity |
| GO:0019864 | CC | IgG binding |
| GO:0050135 | CC | NAD(P)+ nucleosidase activity |
| GO:0061809 | CC | NAD+ nucleotidase, cyclic ADP-ribose generating |
| GO:0045088 | BP | regulation of innate immune response |
| GO:0002831 | BP | regulation of response to biotic stimulus |
| GO:0002764 | BP | immune response-regulating signaling pathway |
| GO:0030217 | BP | T cell differentiation |
| GO:0045824 | BP | negative regulation of innate immune response |
| GO:0031349 | BP | positive regulation of defense response |
| GO:0042581 | MF | specific granule |
| GO:0070820 | MF | tertiary granule |
| GO:0030667 | MF | secretory granule membrane |
| GO:0009897 | MF | external side of plasma membrane |
| GO:0101002 | MF | ficolin-1-rich granule |
| GO:0030669 | MF | clathrin-coated endocytic vesicle membrane |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, J.; Sun, R.; Jin, D.; Yang, Q.; Yang, H.; Long, Y.; Li, L. Identification of Adipocytokine Pathway-Related Genes in Epilepsy and Its Effect on the Peripheral Immune Landscape. Brain Sci. 2022, 12, 1156. https://doi.org/10.3390/brainsci12091156
Chen J, Sun R, Jin D, Yang Q, Yang H, Long Y, Li L. Identification of Adipocytokine Pathway-Related Genes in Epilepsy and Its Effect on the Peripheral Immune Landscape. Brain Sciences. 2022; 12(9):1156. https://doi.org/10.3390/brainsci12091156
Chicago/Turabian StyleChen, Jinkun, Ruohan Sun, Di Jin, Quan Yang, He Yang, Yu Long, and Lixian Li. 2022. "Identification of Adipocytokine Pathway-Related Genes in Epilepsy and Its Effect on the Peripheral Immune Landscape" Brain Sciences 12, no. 9: 1156. https://doi.org/10.3390/brainsci12091156