
LncRNA-Profile-Based Screening of Extracellular Vesicles Released from Brain Endothelial Cells after Oxygen–Glucose Deprivation


Abstract
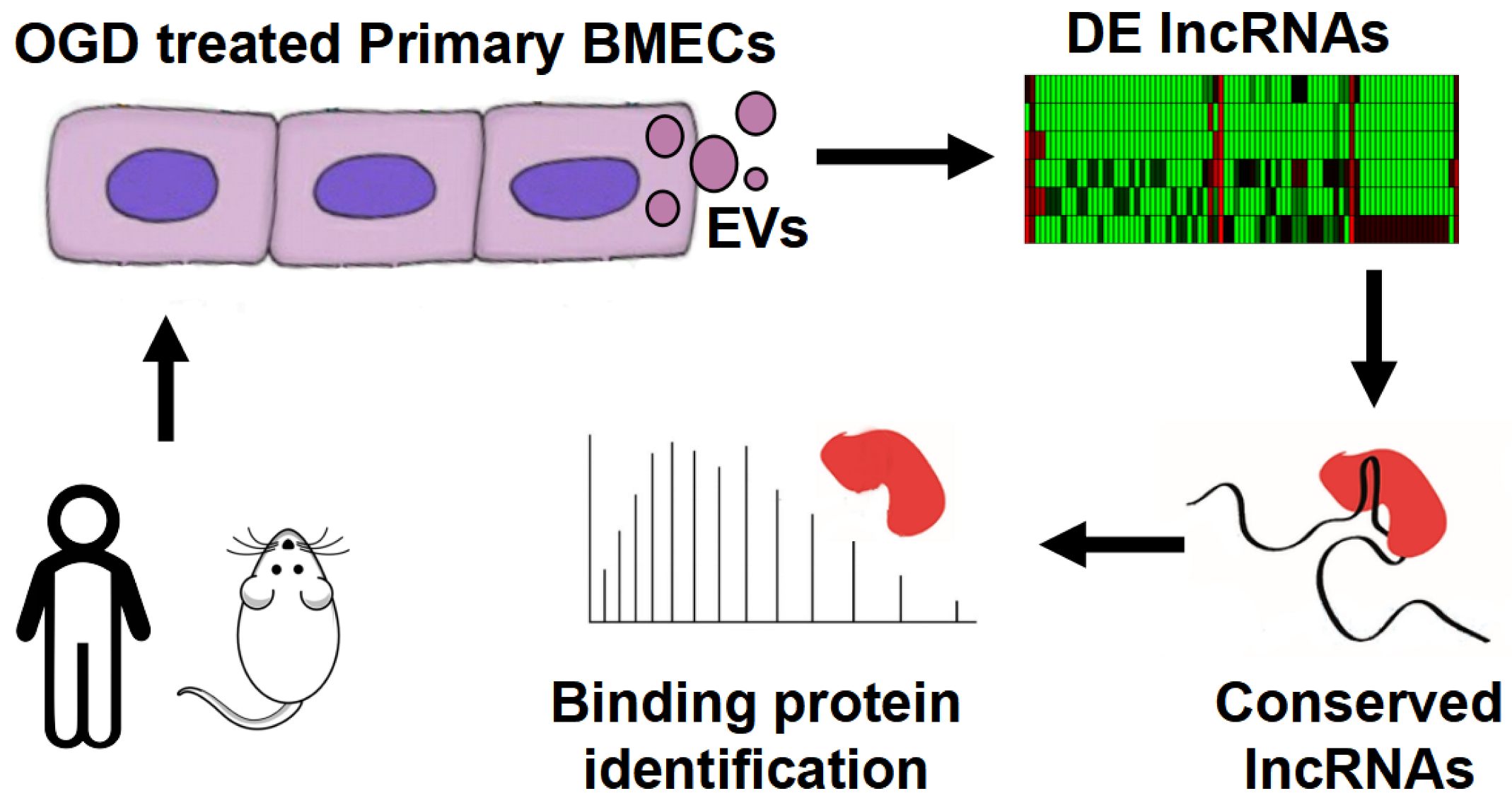
:1. Introduction
2. Methods
2.1. Cell Culture
2.2. EV Isolation
2.3. RNA Sequencing
2.4. Sequencing Data Analysis
2.5. RNA–Protein Pulldown
2.6. Statistical Analysis
3. Results
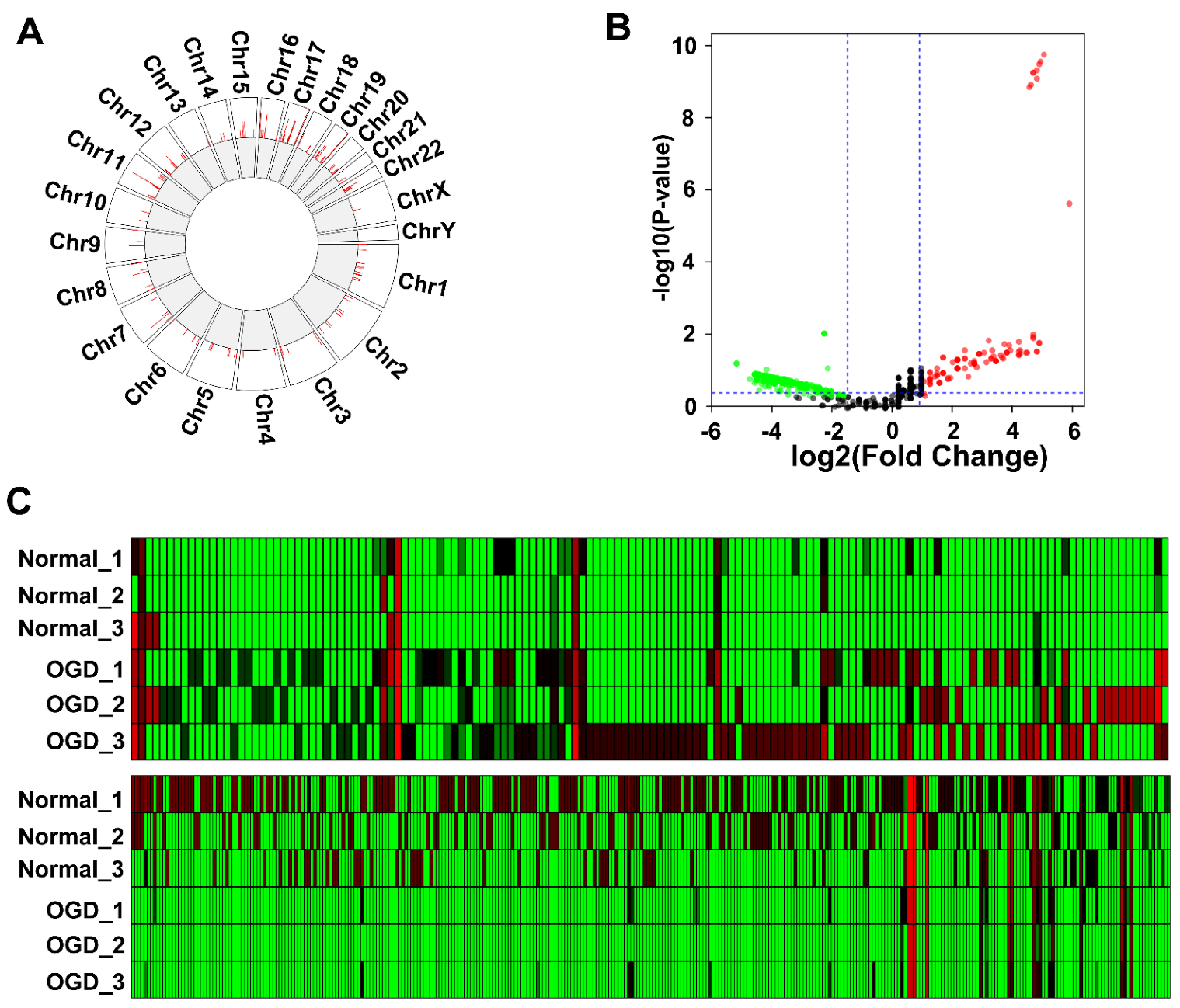
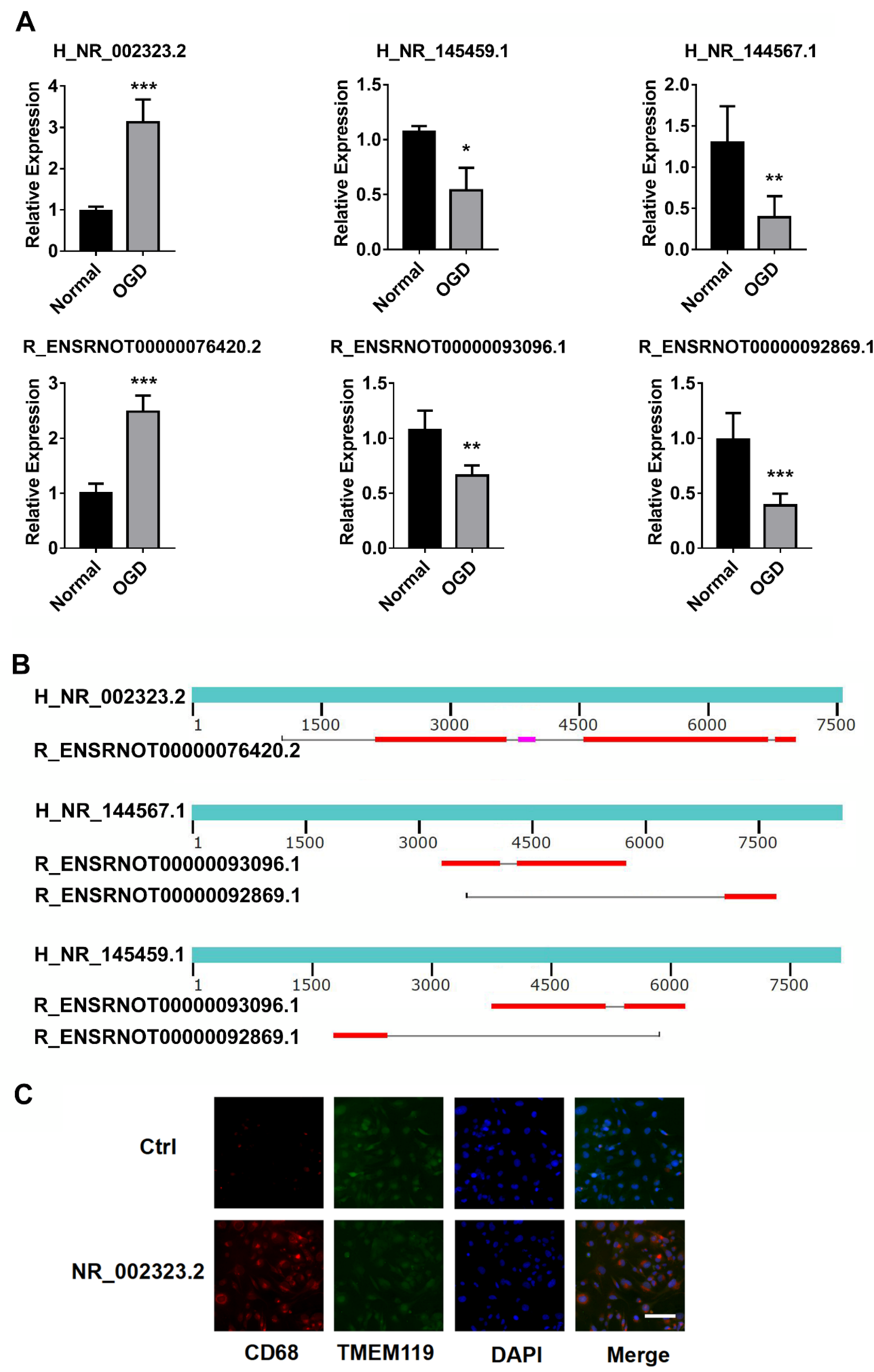
3.1. OGD Induces Differential Expression of LncRNAs in Human BMEC-Derived EVs
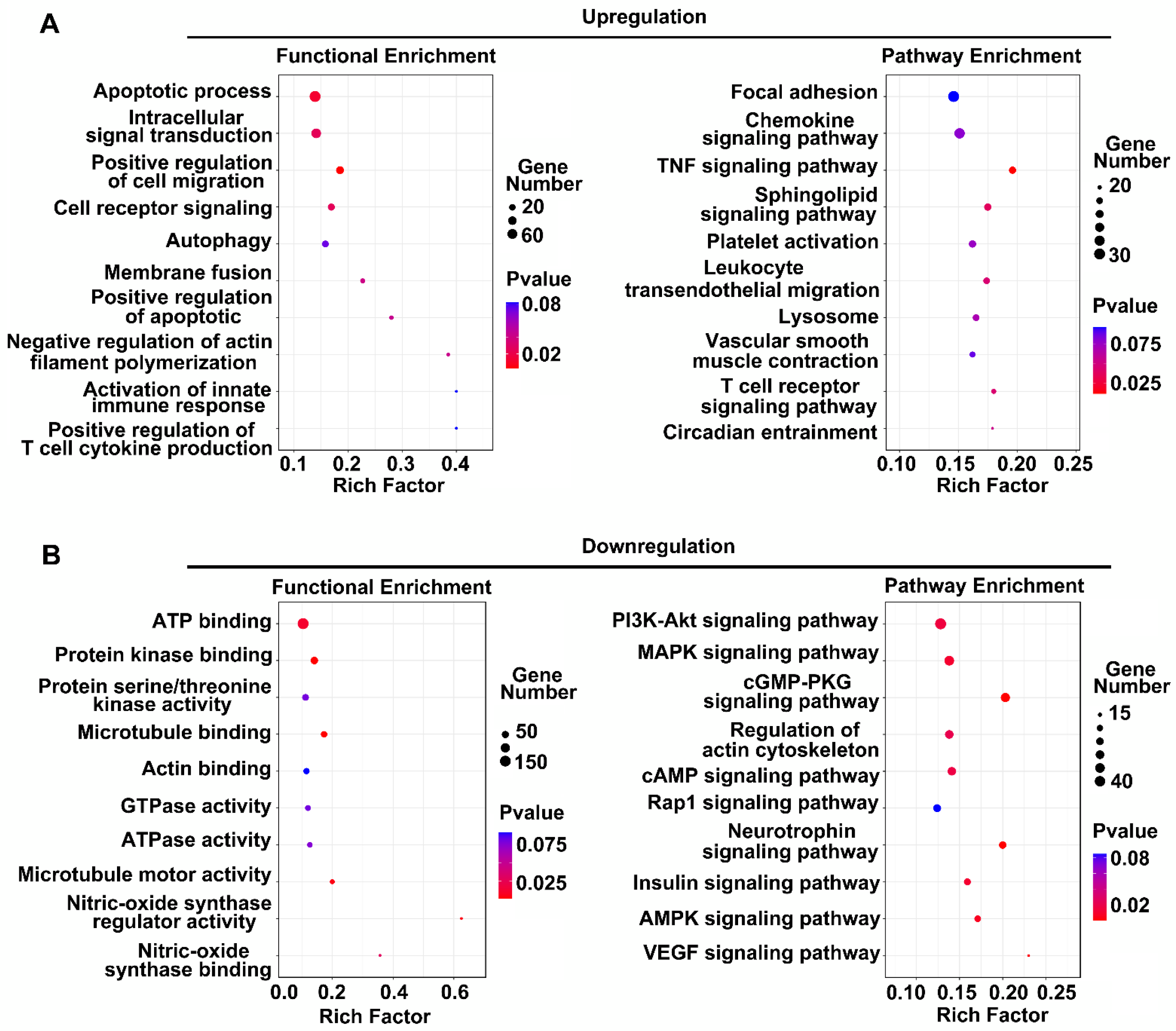
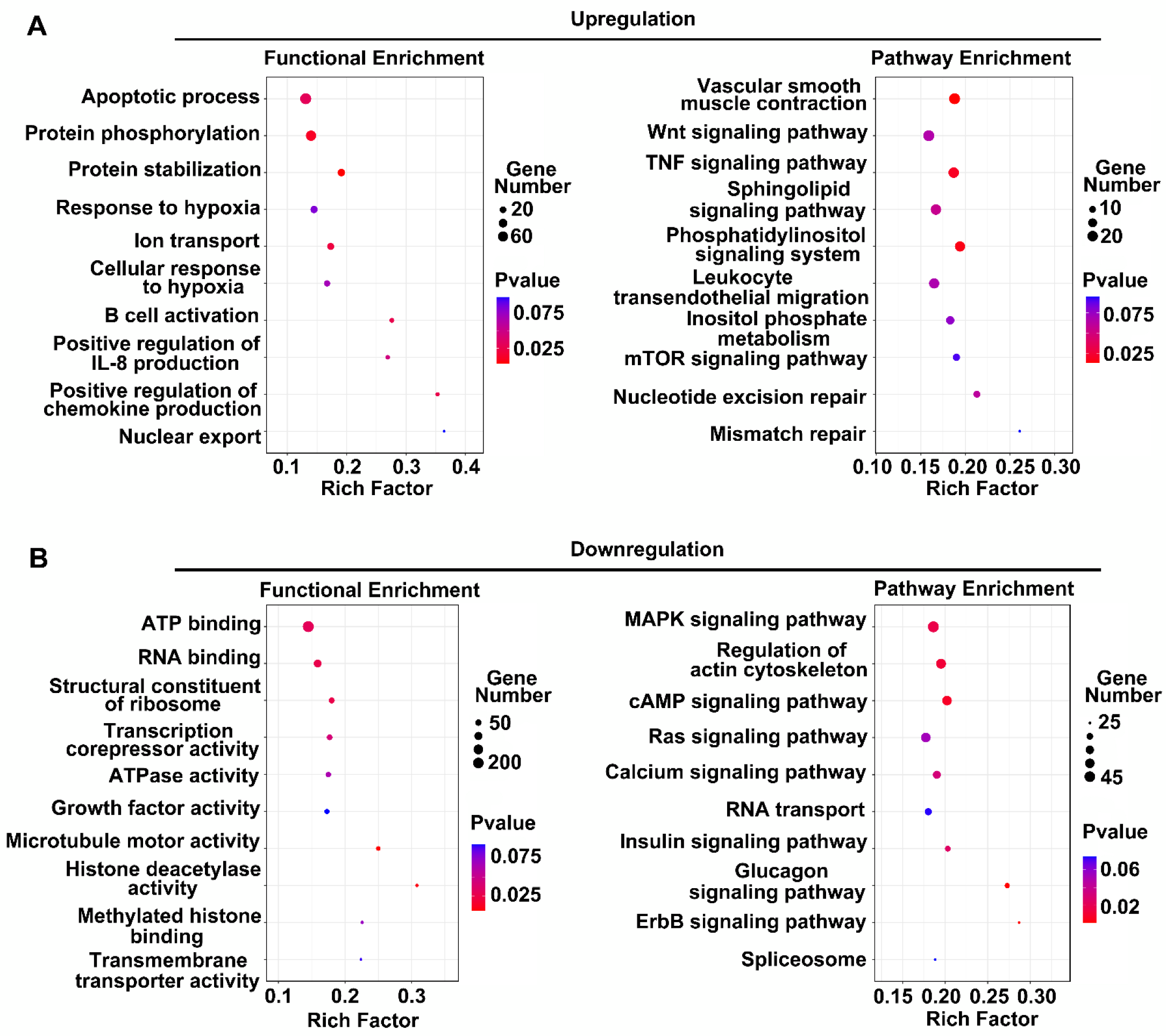
3.2. Functional Analysis of DE LncRNAs in Human BMEC-Derived EVs
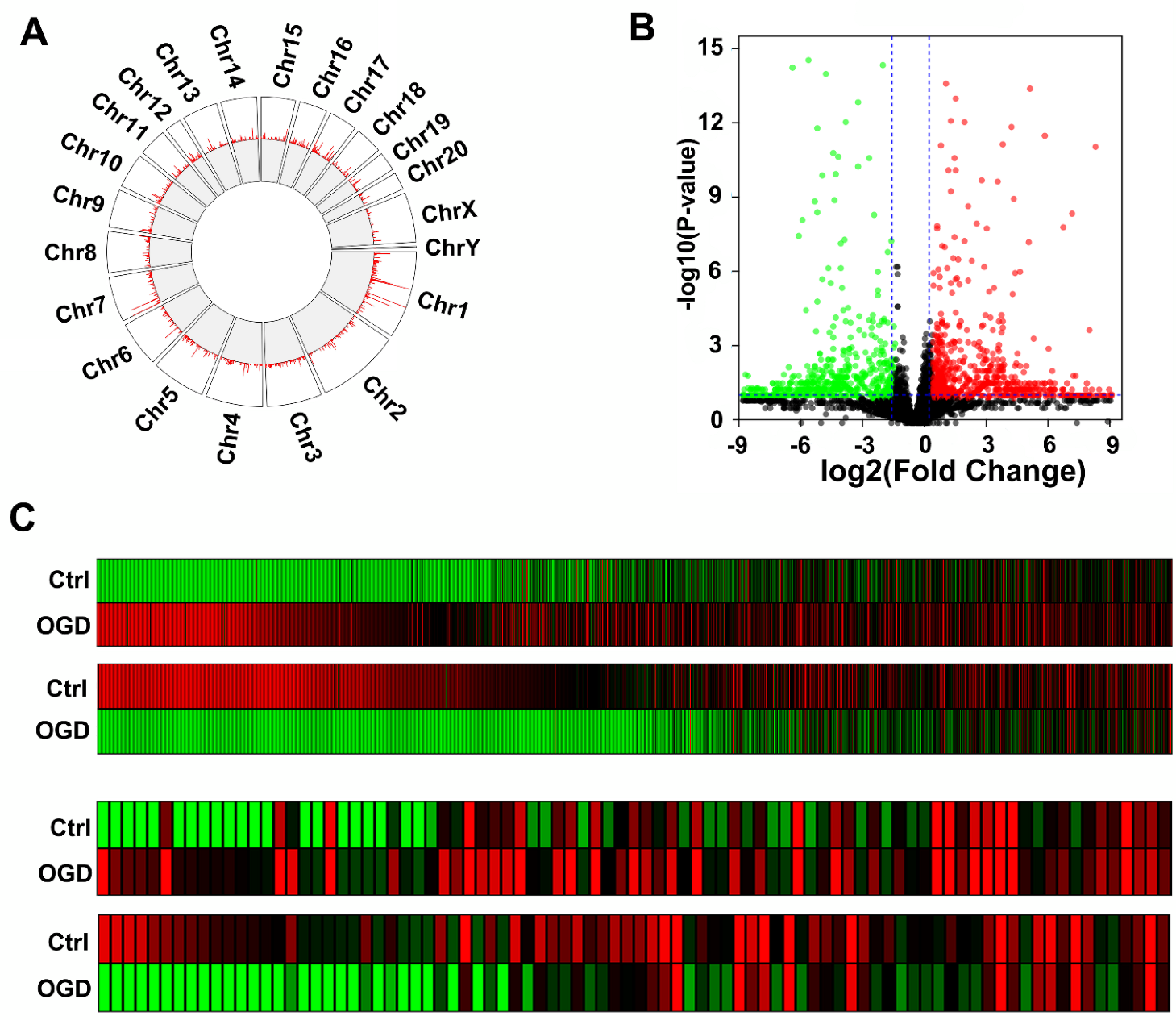
3.3. OGD Induces Differential Expression of LncRNAs in Rat BMEC-Derived EVs
3.4. Functional Analysis of DE LncRNAs in Rat BMEC-Derived EVs
3.5. Protein Binding Analysis of Conserved DE LncRNAs
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Moretti, A.; Ferrari, F.; Villa, R.F. Neuroprotection for ischaemic stroke: Current status and challenges. Pharmacol. Ther. 2015, 146, 23–34. [Google Scholar] [CrossRef] [PubMed]
- Pivoriūnas, A.; Verkhratsky, A. Astrocyte–Endotheliocyte Axis in the Regulation of the Blood–Brain Barrier. Neurochem. Res. 2021, 46, 2538–2550. [Google Scholar] [CrossRef] [PubMed]
- Yu, Q.-J.; Li, M.-C.; Tao, H.; Wang, X. Targeting brain microvascular endothelial cells: A therapeutic approach to neuroprotection against stroke. Neural Regen. Res. 2015, 10, 1882–1891. [Google Scholar] [CrossRef] [PubMed]
- Bargiotas, P.; Krenz, A.; Hormuzdi, S.G.; Ridder, D.A.; Herb, A.; Barakat, W.; Penuela, S.; von Engelhardt, J.; Monyer, H.; Schwaninger, M. Pannexins in ischemia-induced neurodegeneration. Proc. Natl. Acad. Sci. USA 2011, 108, 20772–20777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.-W.; Shang, F.-F.; Liu, W.; Jia, C.; Luo, L.; Wang, H.-L.; Min, X.-L.; Xu, J.-H.; Ma, L.-Q.; Yang, X.-M. Novel circular RNAs expressed in brain microvascular endothelial cells after oxygen-glucose deprivation/recovery. Neural Regen. Res. 2019, 14, 2104–2111. [Google Scholar] [CrossRef]
- Raposo, G.; Stoorvogel, W. Extracellular vesicles: Exosomes, microvesicles, and friends. J. Cell Biol. 2013, 200, 373–383. [Google Scholar] [CrossRef] [Green Version]
- Yáñez-Mó, M.; Siljander, P.R.-M.; Andreu, Z.; Bedina Zavec, A.; Borràs, F.E.; Buzas, E.I.; Buzas, K.; Casal, E.; Cappello, F.; Carvalho, J.; et al. Biological properties of extracellular vesicles and their physiological functions. J. Extracell. Vesicles 2015, 4, 27066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Balkom, B.W.; de Jong, O.G.; Smits, M.; Brummelman, J.; den Ouden, K.; de Bree, P.M.; van Eijndhoven, M.A.J.; Pegtel, D.M.; Wurdinger, T. Endothelial cells require miR-214 to secrete exosomes that suppress senescence and induce angiogenesis in human and mouse endothelial cells. Blood 2013, 121, 3997–4006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dozio, V.; Sanchez, J.-C. Characterisation of extracellular vesicle-subsets derived from brain endothelial cells and analysis of their protein cargo modulation after TNF exposure. J. Extracell. Vesicles 2017, 6, 1302705. [Google Scholar] [CrossRef] [Green Version]
- Xu, H.; Ni, Y.-Q.; Liu, Y.-S. Mechanisms of Action of MiRNAs and LncRNAs in Extracellular Vesicle in Atherosclerosis. Front. Cardiovasc. Med. 2021, 8, 1301. [Google Scholar] [CrossRef]
- Born, L.J.; Harmon, J.W.; Jay, S.M. Therapeutic potential of extracellular vesicle-associated long noncoding RNA. Bioeng. Transl. Med. 2020, 5, e10172. [Google Scholar] [CrossRef] [PubMed]
- Hosen, M.R.; Li, Q.; Liu, Y.; Zietzer, A.; Maus, K.; Goody, P.; Uchida, S.; Latz, E.; Werner, N.; Nickenig, G.; et al. CAD increases the long noncoding RNA PUNISHER in small ex-tracellular vesicles and regulates endothelial cell function via vesicular shuttling. Molecular therapy Nucleic Acids 2021, 25, 388–405. [Google Scholar] [CrossRef]
- Statello, L.; Guo, C.-J.; Chen, L.-L.; Huarte, M. Gene regulation by long non-coding RNAs and its biological functions. Nat. Rev. Mol. Cell Biol. 2021, 22, 96–118, Correction in Nat. Rev. Mol. Cell Biol. 2021, 22, 159. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Bajic, V.B.; Zhang, Z. On the classification of long non-coding RNAs. RNA Biol. 2013, 10, 924–933. [Google Scholar] [CrossRef] [PubMed]
- Michalik, K.M.; You, X.; Manavski, Y.; Doddaballapur, A.; Zörnig, M.; Braun, T.; John, D.; Ponomareva, Y.; Chen, W.; Uchida, S.; et al. Long Noncoding RNA MALAT1 Regulates Endothelial Cell Function and Vessel Growth. Circ. Res. 2014, 114, 1389–1397. [Google Scholar] [CrossRef] [PubMed]
- Lo, E.H.; Dalkara, T.; Moskowitz, M.A. Mechanisms, challenges and opportunities in stroke. Nat. Rev. Neurosci. 2003, 4, 399–414. [Google Scholar] [CrossRef] [PubMed]
- Naoi, M.; Wu, Y.; Shamoto-Nagai, M.; Maruyama, W. Mitochondria in Neuroprotection by Phytochemicals: Bioactive Polyphenols Modulate Mitochondrial Apoptosis System, Function and Structure. Int. J. Mol. Sci. 2019, 20, 2451. [Google Scholar] [CrossRef] [Green Version]
- Shi, C.; Jin, J.; Wang, X.; Song, T.; Li, G.; Li, K.; Ma, J. Sevoflurane attenuates brain damage through inhibiting autophagy and apoptosis in cerebral ischemia-reperfusion rats. Mol. Med. Rep. 2019, 21, 123–130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, S.; Wang, M.; Yang, H.; Mao, L.; He, Q.; Jin, H.; Ye, Z.-M.; Luo, X.-Y.; Xia, Y.-P.; Hu, B. LncRNA TUG1 sponges microRNA-9 to promote neurons apoptosis by up-regulated Bcl2l11 under ischemia. Biochem. Biophys. Res. Commun. 2017, 485, 167–173. [Google Scholar] [CrossRef]
- Tian, C.; Li, Z.; Zhang, L.; Dai, D.; Huang, Q.; Liu, J.; Hong, B. lncRNA NR_120420 promotes SH-SY5Y cells apoptosis by regulating NF-kappaB after oxygen and glucose deprivation. Gene 2020, 728, 144285. [Google Scholar] [CrossRef]
- Gu, C.; Li, L.; Huang, Y.; Qian, D.; Liu, W.; Zhang, C.; Luo, Y.; Zhou, Z.; Kong, F.; Zhao, X.; et al. Salidroside Ameliorates Mitochondria-Dependent Neuronal Apoptosis after Spinal Cord Ischemia-Reperfusion Injury Partially through Inhibiting Oxidative Stress and Promoting Mitophagy. Oxidative Med. Cell. Longev. 2020, 2020, 3549704. [Google Scholar] [CrossRef] [PubMed]
- Ma, W.; Zhang, W.; Cui, B.; Gao, J.; Liu, Q.; Yao, M.; Ning, H.; Xing, L. Functional delivery of lncRNA TUG1 by endothelial progenitor cells derived extracellular vesicles confers anti-inflammatory macrophage polarization in sepsis via impairing miR-9-5p-targeted SIRT1 in-hibition. Cell Death Dis. 2021, 12, 1056. [Google Scholar] [CrossRef] [PubMed]
- Lei, L.; Mou, Q. Exosomal taurine up-regulated 1 promotes angiogenesis and endothelial cell proliferation in cervical cancer. Cancer Biol. Ther. 2020, 21, 717–725. [Google Scholar] [CrossRef]
- Wen, H.; Li, L.; Zhan, L.; Zuo, Y.; Li, K.; Qiu, M.; Li, H.; Sun, W.; Xu, E. Hypoxic postconditioning promotes mitophagy against transient global cerebral ischemia via PINK1/Parkin-induced mitochondrial ubiquitination in adult rats. Cell Death Dis. 2021, 12, 630. [Google Scholar] [CrossRef] [PubMed]
- Dawson, T.M.; Dawson, V.L. Mitochondrial Mechanisms of Neuronal Cell Death: Potential Therapeutics. Annu. Rev. Pharmacol. Toxicol. 2017, 57, 437–454. [Google Scholar] [CrossRef] [PubMed]
- Sang, L.; Ju, H.-Q.; Yang, Z.; Ge, Q.; Zhang, Z.; Liu, F.; Yang, L.; Gong, H.; Shi, C.; Qu, L.; et al. Mitochondrial long non-coding RNA GAS5 tunes TCA metabolism in response to nutrient stress. Nat. Metab. 2021, 3, 90–106. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Li, S.; Zhang, Y.; Wang, M.; Li, X.; Liu, S.; Xu, D.; Bao, Y.; Jia, P.; Wu, N.; et al. The lncRNA Malat1 regulates microvascular function after myocardial infarction in mice via miR-26b-5p/Mfn1 axis-mediated mitochondrial dynamics. Redox Biol. 2021, 41, 101910. [Google Scholar] [CrossRef]
- Wang, H.; Zhang, Y.; Guan, X.; Li, X.; Zhao, Z.; Gao, Y.; Zhang, X.; Chen, R. An Integrated Transcriptomics and Proteomics Analysis Implicates lncRNA MALAT1 in the Regulation of Lipid Metabolism. Mol. Cell. Proteom. 2021, 20, 100141. [Google Scholar] [CrossRef] [PubMed]
- Zong, X.; Nakagawa, S.; Freier, S.M.; Fei, J.; Ha, T.; Prasanth, S.G.; Prasanth, K.V. Natural antisense RNA promotes 3′ end processing and maturation of MALAT1 lncRNA. Nucleic Acids Res. 2016, 44, 2898–2908. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pichon, X.; Wilson, L.A.; Stoneley, M.; Bastide, A.; King, H.A.; Somers, J.; Willis, A.E. RNA Binding Protein/RNA Element Interactions and the Control of Translation. Curr. Protein Pept. Sci. 2012, 13, 294–304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferre, F.; Colantoni, A.; Helmer-Citterich, M. Revealing protein-lncRNA interaction. Brief. Bioinform. 2016, 17, 106–116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gardner, L.B. Hypoxic Inhibition of Nonsense-Mediated RNA Decay Regulates Gene Expression and the Integrated Stress Response. Mol. Cell. Biol. 2008, 28, 3729–3741. [Google Scholar] [CrossRef] [Green Version]
- Jia, J.; Furlan, A.; Gonzalez-Hilarion, S.; Leroy, C.; Gruenert, D.C.; Tulasne, D.; Lejeune, F. Caspases shutdown nonsense-mediated mRNA decay during apoptosis. Cell Death Differ. 2015, 22, 1754–1763. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Wu, W.; Han, Q.; Wang, Y.; Li, C.; Zhang, P.; Xu, H. New Insights into the Interplay between Non-Coding RNAs and RNA-Binding Protein HnRNPK in Regulating Cellular Functions. Cells 2019, 8, 62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zietzer, A.; Hosen, M.R.; Wang, H.; Goody, P.R.; Sylvester, M.; Latz, E.; Nickenig, G.; Werner, N.; Jansen, F. The RNA-binding protein hnRNPU regulates the sorting of microRNA-30c-5p into large extracellular vesicles. J. Extracell. Vesicles 2020, 9, 1786967. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.-C.; Ou, A.C.; Park, J.; Yu, F.; Yu, B.; Lee, A.; Yang, G.; Zhou, A.; Benz, E.J. RBFOX2 Promotes Protein 4.1R Exon 16 Selection via U1 snRNP Recruitment. Mol. Cell. Biol. 2012, 32, 513–526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, J.; Verma, S.K.; Jaworski, E.; Mohan, S.; Nagasawa, C.K.; Rayavara, K.; Sooter, A.; Miller, S.N.; Holcomb, R.J.; Powell, M.J.; et al. RBFOX2 is critical for maintaining alternative polyadenylation patterns and mitochondrial health in rat myoblasts. Cell Rep. 2021, 37. [Google Scholar] [CrossRef] [PubMed]
- Graham, B.H.; Waymire, K.G.; Cottrell, B.; Trounce, I.A.; MacGregor, G.R.; Wallace, D.C. A mouse model for mitochondrial myopathy and cardiomyopathy resulting from a deficiency in the heart/muscle isoform of the adenine nucleotide translocator. Nat. Genet. 1997, 16, 226–234. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| H_LncRNA | Conservation% | R_LncRNA | Conservation% |
|---|---|---|---|
| NR_002323.2 | 62.42 | ENSRNOT00000076420.2 | 91.94 |
| NR_110493.1 | 62.35 | ENSRNOT00000076420.2 | 91.66 |
| NR_110492.1 | 61.07 | ENSRNOT00000076420.2 | 90.61 |
| NR_145459.1 | 26.83 | ENSRNOT00000093096.1 | 92.72 |
| NR_144567.1 | 25.5 | ENSRNOT00000093096.1 | 92.72 |
| NR_002819.4 | 24.82 | ENSRNOT00000093096.1 | 92.72 |
| NR_144568.1 | 23.73 | ENSRNOT00000093096.1 | 83.83 |
| ENST00000537157.1 | 20.88 | ENSRNOT00000090171.1 | 10.28 |
| NR_002809.2 | 13.15 | ENSRNOT00000090171.1 | 20.55 |
| NR_026794.1 | 8.32 | ENSRNOT00000087244.1 | 45.89 |
| NR_026794.1 | 8.32 | ENSRNOT00000081933.1 | 38.66 |
| NR_144568.1 | 7.72 | ENSRNOT00000092949.1 | 92.23 |
| ENST00000567556.1 | 7.66 | ENSRNOT00000035590.6 | 1.72 |
| NR_144567.1 | 7.5 | ENSRNOT00000092949.1 | 92.23 |
| NR_145459.1 | 7.34 | ENSRNOT00000092869.1 | 84.06 |
| NR_002819.4 | 7.3 | ENSRNOT00000092949.1 | 92.23 |
| NR_144568.1 | 7.18 | ENSRNOT00000092869.1 | 84.06 |
| NR_144567.1 | 6.97 | ENSRNOT00000092869.1 | 84.06 |
| NR_002819.4 | 6.79 | ENSRNOT00000092869.1 | 84.06 |
| NR_109854.1 | 6.08 | ENSRNOT00000090651.1 | 11.1 |
| NR_109853.1 | 5.96 | ENSRNOT00000090651.1 | 11.1 |
| NR_145459.1 | 5.1 | ENSRNOT00000092949.1 | 59.57 |
| NR_015431.2 | 4.38 | ENSRNOT00000090651.1 | 11.1 |
| NR_109852.1 | 4.24 | ENSRNOT00000090651.1 | 11.1 |
| NR_109851.1 | 4.21 | ENSRNOT00000090651.1 | 11.1 |
| NR_109850.1 | 4.21 | ENSRNOT00000090651.1 | 11.1 |
| NR_109855.1 | 3.45 | ENSRNOT00000090651.1 | 4.29 |
| NR_102327.1 | 1.43 | ENSRNOT00000086884.1 | 3.16 |
| NR_024053.2 | 0.7 | ENSRNOT00000086884.1 | 3.16 |
| NR_024052.2 | 0.67 | ENSRNOT00000086884.1 | 3.16 |
| NR_102326.1 | 0.54 | ENSRNOT00000086884.1 | 3.16 |
| NR_125909.1 | 0.14 | ENSRNOT00000081606.1 | 1.63 |
| Probe | Accession | Name | MW [kDa] | Score | Coverage% | Peptides |
|---|---|---|---|---|---|---|
| NR_002323.2 | Q92900 | RENT1 | 124.3 | 100.23 | 3.84 | 3 |
| Q08170 | SRSF4 | 56.7 | 160.7 | 5.18 | 2 | |
| Q6NTF9 | RHBD2 | 39.2 | 378.08 | 13.64 | 5 | |
| NR_145459.1 | Q00839 | HNRPU | 88.9 | 128.85 | 2.95 | 2 |
| Q9NR90 | DAZ3 | 54.9 | 175.25 | 10.18 | 3 | |
| P55795 | HNRH2 | 49.2 | 107.67 | 5.74 | 2 | |
| NR_144567.1 | O43251 | RFOX2 | 41.3 | 291.68 | 37.11 | 8 |
| A6NDY0 | EPAB2 | 30.3 | 106.23 | 6.04 | 2 | |
| Q9UJJ7 | RUSD1 | 34.7 | 217.54 | 25.65 | 4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, X.; Zou, H.; Lyu, Q.; Tang, Y.; Xiong, W.; Shang, F.-F. LncRNA-Profile-Based Screening of Extracellular Vesicles Released from Brain Endothelial Cells after Oxygen–Glucose Deprivation. Brain Sci. 2022, 12, 1027. https://doi.org/10.3390/brainsci12081027
He X, Zou H, Lyu Q, Tang Y, Xiong W, Shang F-F. LncRNA-Profile-Based Screening of Extracellular Vesicles Released from Brain Endothelial Cells after Oxygen–Glucose Deprivation. Brain Sciences. 2022; 12(8):1027. https://doi.org/10.3390/brainsci12081027
Chicago/Turabian StyleHe, Xiang, Hecun Zou, Qiang Lyu, Yujing Tang, Wenkui Xiong, and Fei-Fei Shang. 2022. "LncRNA-Profile-Based Screening of Extracellular Vesicles Released from Brain Endothelial Cells after Oxygen–Glucose Deprivation" Brain Sciences 12, no. 8: 1027. https://doi.org/10.3390/brainsci12081027