
Altered Development of Prefrontal GABAergic Functions and Anxiety-like Behavior in Adolescent Offspring Induced by Prenatal Stress


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Maternal Restraint Stress
2.3. Elevated plus Maze
2.4. Tissue Preparation
2.5. Reagents
2.6. Western Blot Analysis
2.7. Immunohistochemistry
2.8. Image Analysis
2.9. Statistical Analysis
3. Results
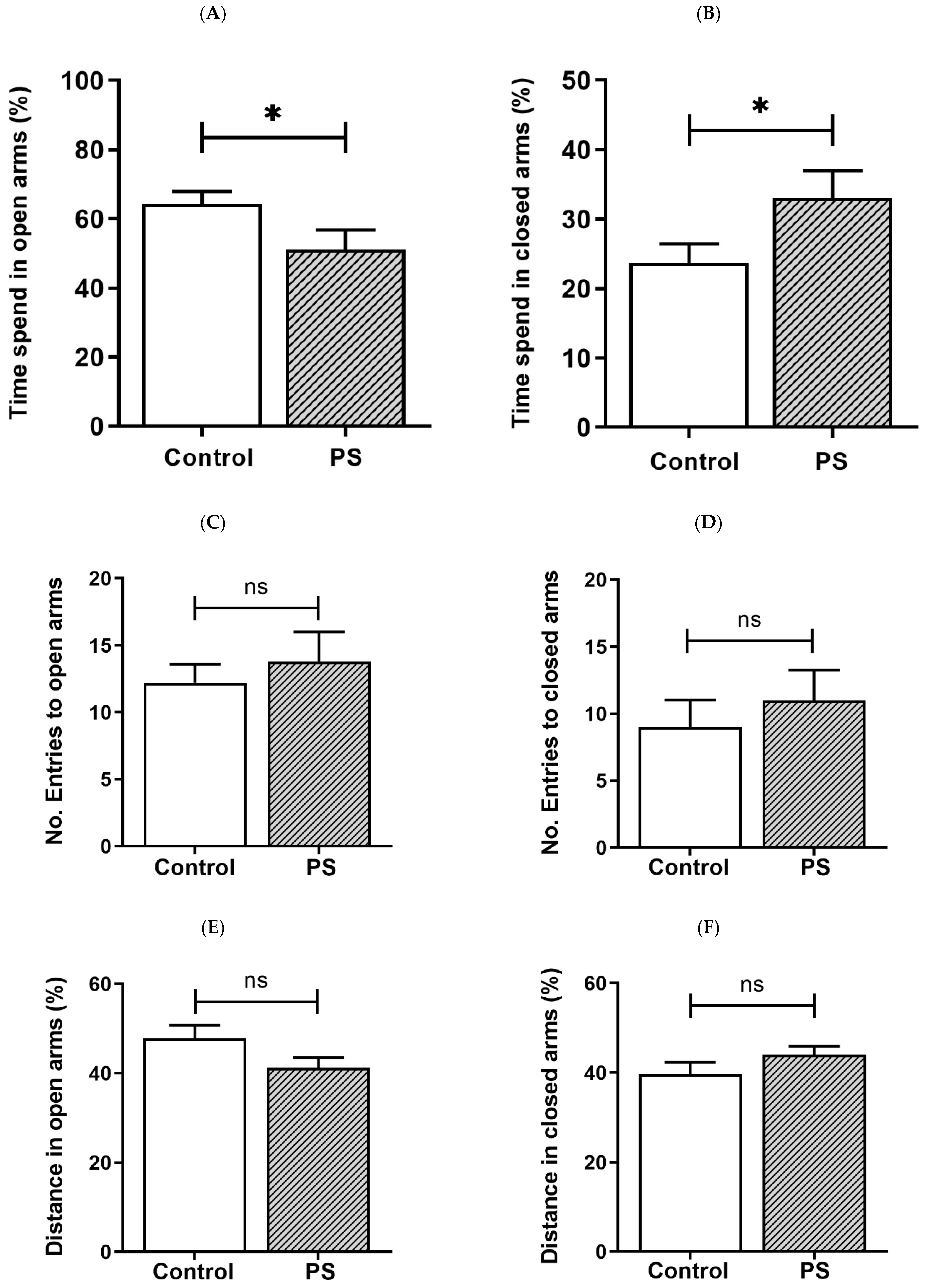
3.1. Anxiety-like Behavior in Adolescent Rats
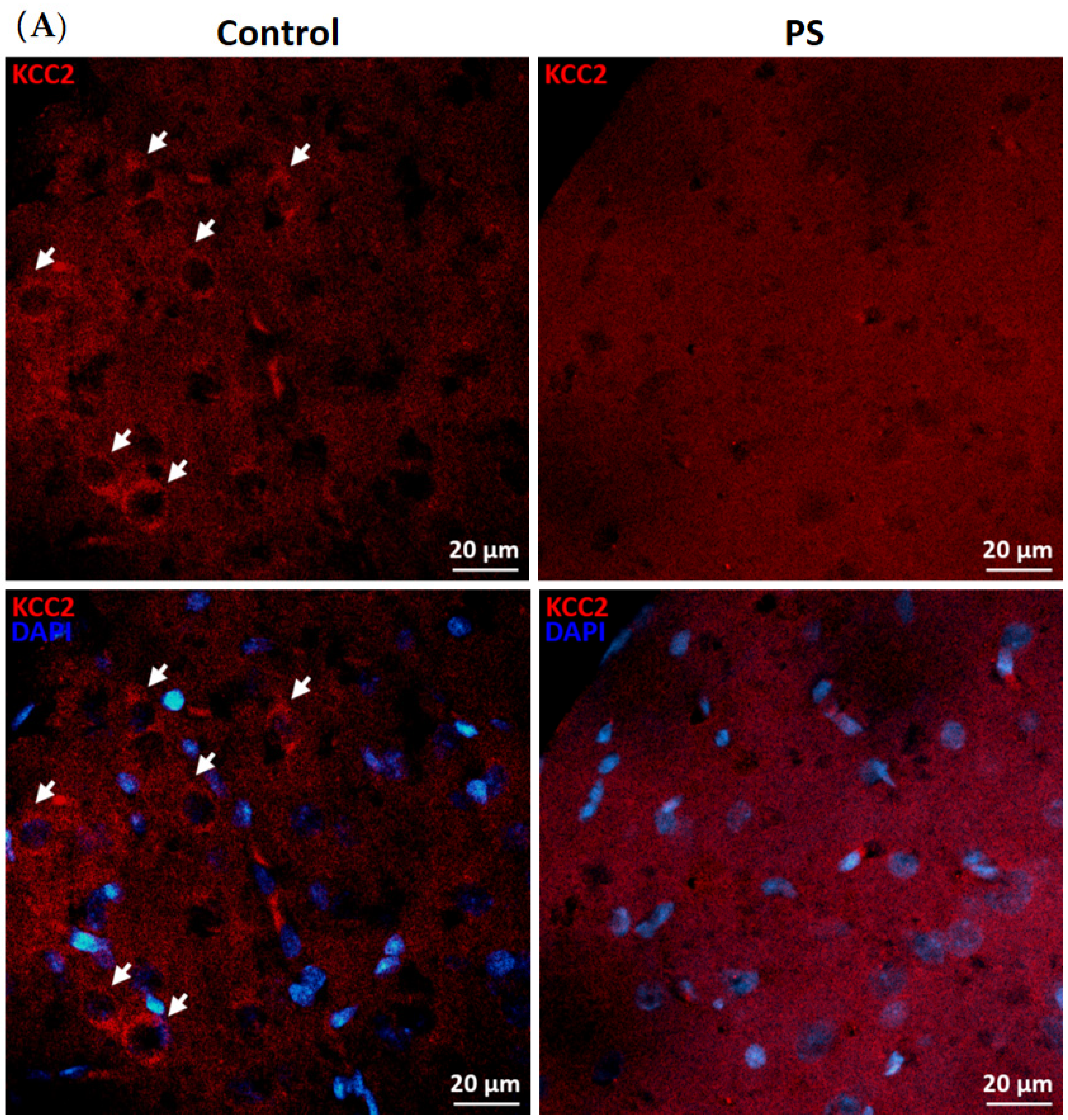
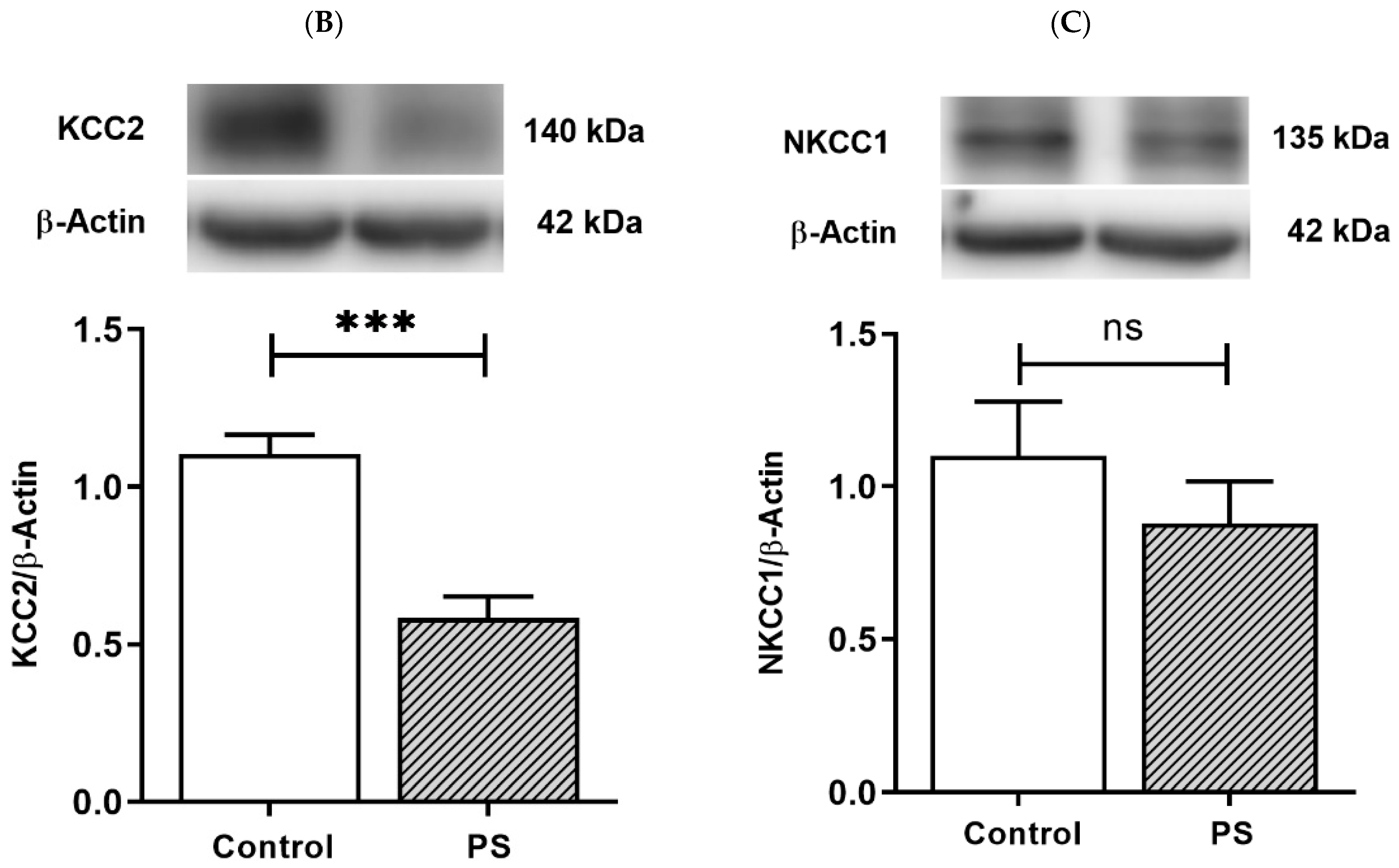
3.2. Developmental Shift of Cation-Chloride Cotransporters
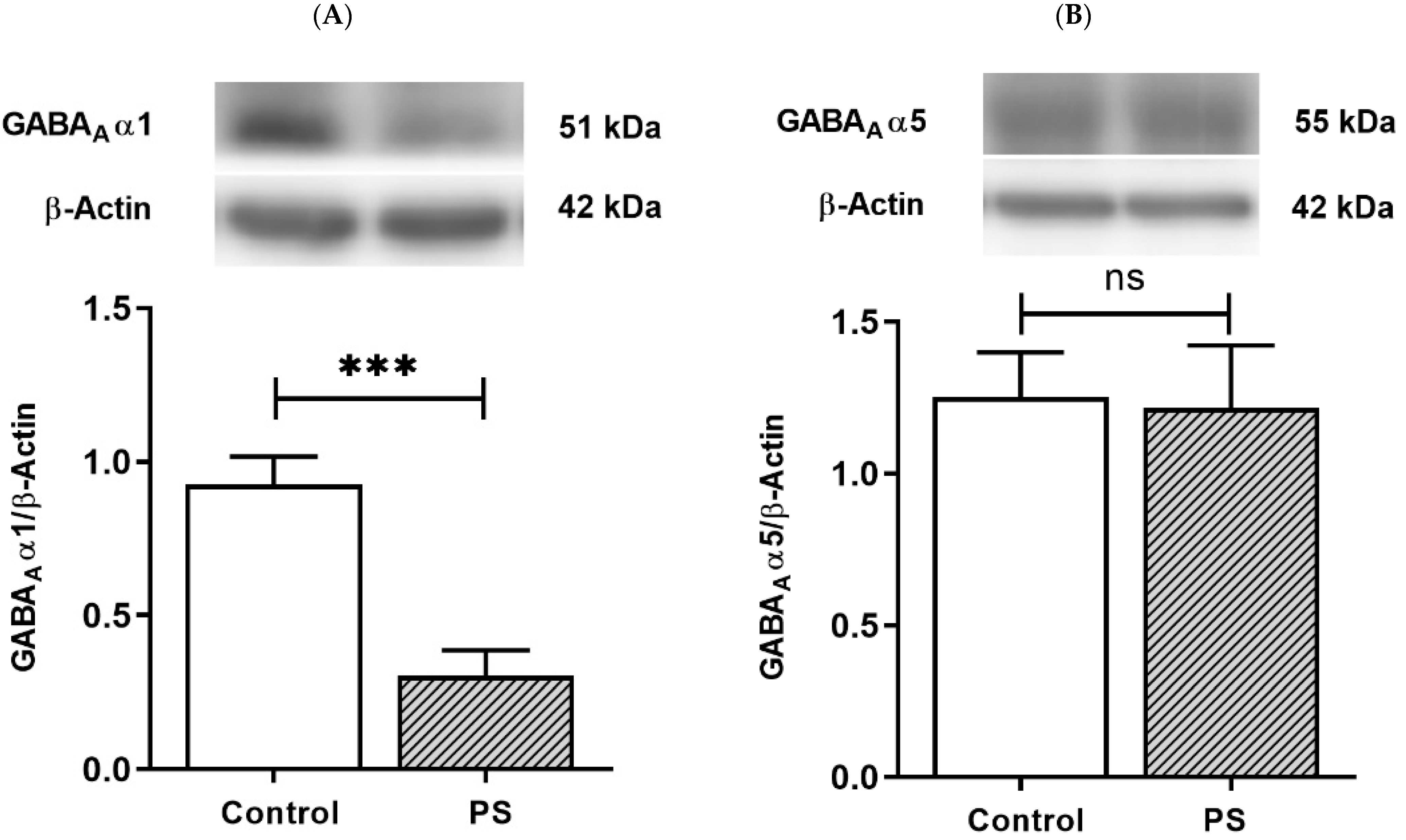
3.3. GABA Type A Receptors Functional Development
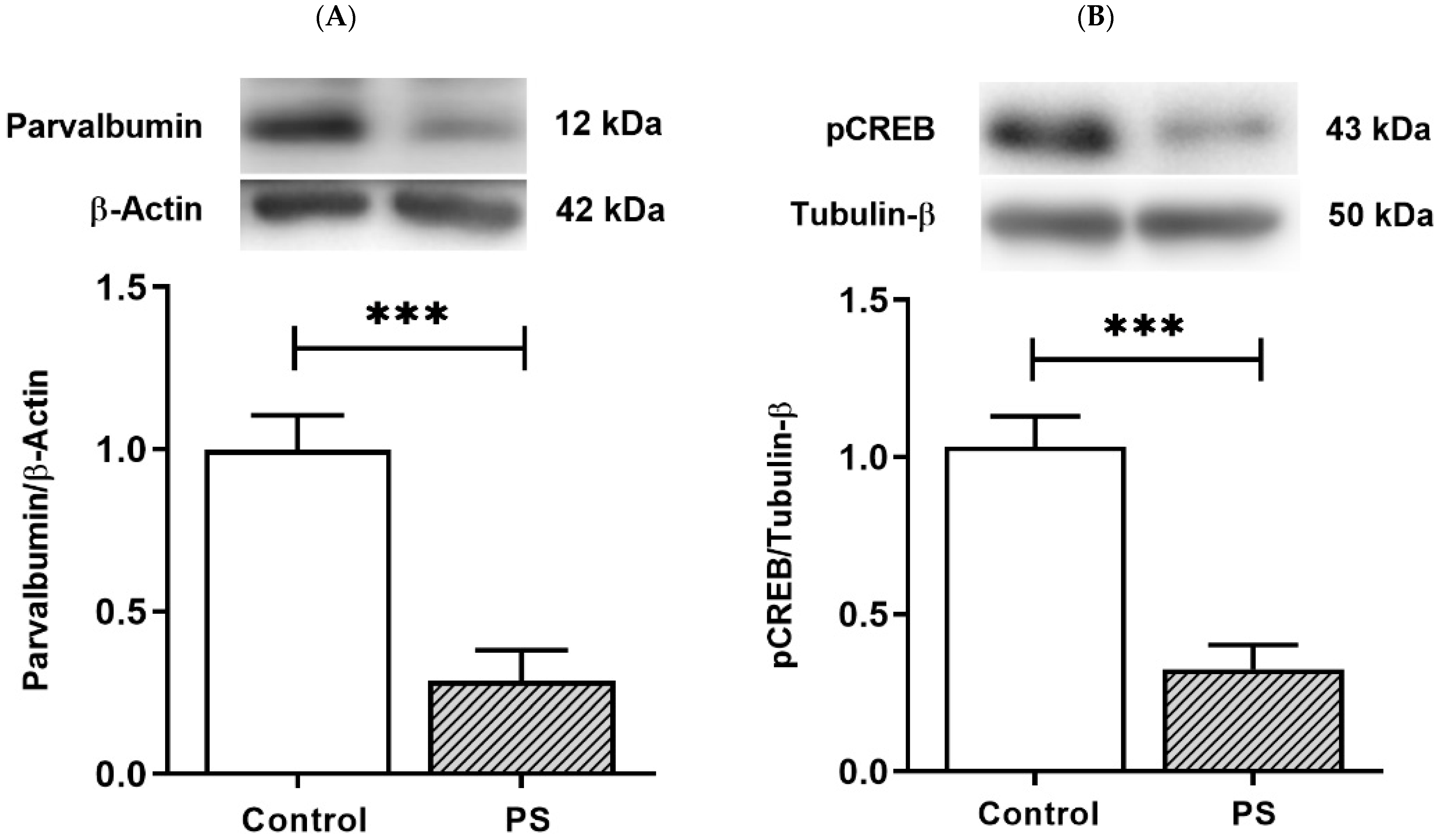
3.4. The Plasticity Regulation of GABAergic Interneurons
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Reynolds, R.M. Glucocorticoid excess and the developmental origins of disease: Two decades of testing the hypothesis—2012 Curt Richter Award Winner. Psychoneuroendocrinology 2013, 38, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Weinstock, M. Prenatal stressors in rodents: Effects on behavior. Neurobiol. Stress 2017, 6, 3–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bock, J.; Wainstock, T.; Braun, K.; Segal, M. Stress In Utero: Prenatal Programming of Brain Plasticity and Cognition. Biol. Psychiatry 2015, 78, 315–326. [Google Scholar] [CrossRef] [PubMed]
- Van den Bergh, B.R.H.; van den Heuvel, M.I.; Lahti, M.; Braeken, M.; de Rooij, S.R.; Entringer, S.; Hoyer, D.; Roseboom, T.; Räikkönen, K.; King, S.; et al. Prenatal developmental origins of behavior and mental health: The influence of maternal stress in pregnancy. Neurosci. Biobehav. Rev. 2020, 117, 26–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mills, K.L.; Goddings, A.L.; Clasen, L.S.; Giedd, J.N.; Blakemore, S.J. The Developmental Mismatch in Structural Brain Maturation during Adolescence. Dev. Neurosci. 2014, 36, 147–160. [Google Scholar] [CrossRef] [PubMed]
- Caballero, A.; Granberg, R.; Tseng, K.Y. Mechanisms contributing to prefrontal cortex maturation during adolescence. Neurosci. Biobehav. Rev. 2016, 70, 4–12. [Google Scholar] [CrossRef] [Green Version]
- Sah, P. Interneurons in the Prefrontal Cortex: A Role in the Genesis of Anxiety in Adolescence? Biol. Psychiatry 2019, 86, 650–651. [Google Scholar] [CrossRef]
- Newman, E.; Thompson, W.K.; Bartsch, H.; Hagler, D.J.; Chen, C.-H.; Brown, T.T.; Kuperman, J.M.; McCabe, C.; Chung, Y.; Libiger, O.; et al. Anxiety is related to indices of cortical maturation in typically developing children and adolescents. Brain Struct. Funct. 2016, 221, 3013–3025. [Google Scholar] [CrossRef]
- Caballero, A.; Tseng, K.Y. GABAergic Function as a Limiting Factor for Prefrontal Maturation during Adolescence. Trends Neurosci. 2016, 39, 441–448. [Google Scholar] [CrossRef] [Green Version]
- Zimmermann, K.S.; Richardson, R.; Baker, K.D. Maturational Changes in Prefrontal and Amygdala Circuits in Adolescence: Implications for Understanding Fear Inhibition during a Vulnerable Period of Development. Brain Sci. 2019, 9, 65. [Google Scholar] [CrossRef] [Green Version]
- Tang, X.; Jaenisch, R.; Sur, M. The role of GABAergic signalling in neurodevelopmental disorders. Nat. Rev. Neurosci. 2021, 22, 290–307. [Google Scholar] [CrossRef] [PubMed]
- Ben-Ari, Y. Excitatory actions of gaba during development: The nature of the nurture. Nat. Rev. Neurosci. 2002, 3, 728–739. [Google Scholar] [CrossRef] [PubMed]
- Ben-Ari, Y. The GABA excitatory/inhibitory developmental sequence: A personal journey. Neuroscience 2014, 279, 187–219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ben-Ari, Y.; Khalilov, I.; Kahle, K.T.; Cherubini, E. The GABA Excitatory/Inhibitory Shift in Brain Maturation and Neurological Disorders. Neuroscientist 2012, 18, 467–486. [Google Scholar] [CrossRef]
- Knoflach, F.; Hernandez, M.-C.; Bertrand, D. GABAA receptor-mediated neurotransmission: Not so simple after all. Biochem. Pharmacol. 2016, 115, 10–17. [Google Scholar] [CrossRef]
- Duncan, C.E.; Webster, M.J.; Rothmond, D.A.; Bahn, S.; Elashoff, M.; Shannon Weickert, C. Prefrontal GABAA receptor α-subunit expression in normal postnatal human development and schizophrenia. J. Psychiatr. Res. 2010, 44, 673–681. [Google Scholar] [CrossRef]
- Datta, D.; Arion, D.; Lewis, D.A. Developmental Expression Patterns of GABAA Receptor Subunits in Layer 3 and 5 Pyramidal Cells of Monkey Prefrontal Cortex. Cereb. Cortex 2015, 25, 2295–2305. [Google Scholar] [CrossRef] [Green Version]
- Bosman, L.W.J.; Rosahl, T.W.; Brussaard, A.B. Neonatal development of the rat visual cortex: Synaptic function of GABAa receptor α subunits. J. Physiol. 2002, 545, 169–181. [Google Scholar] [CrossRef]
- Vicini, S.; Ferguson, C.; Prybylowski, K.; Kralic, J.; Morrow, A.L.; Homanics, G.E. GABAA Receptor α1 Subunit Deletion Prevents Developmental Changes of Inhibitory Synaptic Currents in Cerebellar Neurons. J. Neurosci. 2001, 21, 3009. [Google Scholar] [CrossRef] [Green Version]
- Caraiscos Valerie, B.; Elliott Erin, M.; You-Ten Kong, E.; Cheng Victor, Y.; Belelli, D.; Newell, J.G.; Jackson Michael, F.; Lambert Jeremy, J.; Rosahl Thomas, W.; Wafford Keith, A.; et al. Tonic inhibition in mouse hippocampal CA1 pyramidal neurons is mediated by α5 subunit-containing γ-aminobutyric acid type A receptors. Proc. Natl. Acad. Sci. USA 2004, 101, 3662–3667. [Google Scholar] [CrossRef] [Green Version]
- Farrant, M.; Nusser, Z. Variations on an inhibitory theme: Phasic and tonic activation of GABAA receptors. Nat. Rev. Neurosci. 2005, 6, 215–229. [Google Scholar] [CrossRef] [PubMed]
- Veerawatananan, B.; Surakul, P.; Chutabhakdikul, N. Maternal restraint stress delays maturation of cation-chloride cotransporters and GABAA receptor subunits in the hippocampus of rat pups at puberty. Neurobiol. Stress 2016, 3, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berry, A.; Panetta, P.; Luoni, A.; Bellisario, V.; Capoccia, S.; Riva, M.A.; Cirulli, F. Decreased Bdnf expression and reduced social behavior in periadolescent rats following prenatal stress. Dev. Psychobiol. 2015, 57, 365–373. [Google Scholar] [CrossRef] [PubMed]
- Ehrlich, D.E.; Neigh, G.N.; Bourke, C.H.; Nemeth, C.L.; Hazra, R.; Ryan, S.J.; Rowson, S.; Jairam, N.; Sholar, C.A.; Rainnie, D.G.; et al. Prenatal stress, regardless of concurrent escitalopram treatment, alters behavior and amygdala gene expression of adolescent female rats. Neuropharmacology 2015, 97, 251–258. [Google Scholar] [CrossRef] [Green Version]
- Woodward, E.M.; Coutellier, L. Age- and sex-specific effects of stress on parvalbumin interneurons in preclinical models: Relevance to sex differences in clinical neuropsychiatric and neurodevelopmental disorders. Neurosci. Biobehav. Rev. 2021, 131, 1228–1242. [Google Scholar] [CrossRef]
- Lussier, S.J.; Stevens, H.E. Delays in GABAergic interneuron development and behavioral inhibition after prenatal stress. Dev. Neurobiol. 2016, 76, 1078–1091. [Google Scholar] [CrossRef] [PubMed]
- Cohen, S.M.; Ma, H.; Kuchibhotla, K.V.; Watson, B.O.; Buzsáki, G.; Froemke, R.C.; Tsien, R.W. Excitation-Transcription Coupling in Parvalbumin-Positive Interneurons Employs a Novel CaM Kinase-Dependent Pathway Distinct from Excitatory Neurons. Neuron 2016, 90, 292–307. [Google Scholar] [CrossRef] [Green Version]
- Roshan-Milani, S.; Seyyedabadi, B.; Saboory, E.; Parsamanesh, N.; Mehranfard, N. Prenatal stress and increased susceptibility to anxiety-like behaviors: Role of neuroinflammation and balance between GABAergic and glutamatergic transmission. Stress 2021, 24, 481–495. [Google Scholar] [CrossRef]
- Chutabhakdikul, N.; Surakul, P. Prenatal stress increased Snk Polo-like kinase 2, SCF β-TrCP ubiquitin ligase and ubiquitination of SPAR in the hippocampus of the offspring at adulthood. Int. J. Dev. Neurosci. 2013, 31, 560–567. [Google Scholar] [CrossRef]
- Freide, E.; Weinstock, M. The effects of prenatal exposure to predictable or unpredictable stress on early development in the rat. Dev. Psychobiol. 1984, 17, 651–660. [Google Scholar] [CrossRef]
- Walf, A.A.; Frye, C.A. The use of the elevated plus maze as an assay of anxiety-related behavior in rodents. Nat. Protoc. 2007, 2, 322–328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ari, C.; D’Agostino, D.P.; Diamond, D.M.; Kindy, M.; Park, C.; Kovács, Z. Elevated Plus Maze Test Combined with Video Tracking Software to Investigate the Anxiolytic Effect of Exogenous Ketogenic Supplements. JoVE 2019, 143, e58396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holliday, E.; Gould, T.J. Nicotine, adolescence, and stress: A review of how stress can modulate the negative consequences of adolescent nicotine abuse. Neurosci. Biobehav. Rev. 2016, 65, 173–184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eiland, L.; Romeo, R.D. Stress and the developing adolescent brain. Neuroscience 2013, 249, 162–171. [Google Scholar] [CrossRef] [Green Version]
- Crestani, C.C. Adolescent vulnerability to cardiovascular consequences of chronic emotional stress: Review and perspectives for future research. Neurosci. Biobehav. Rev. 2017, 74, 466–475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le Magueresse, C.; Monyer, H. GABAergic Interneurons Shape the Functional Maturation of the Cortex. Neuron 2013, 77, 388–405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cass, D.K.; Flores-Barrera, E.; Thomases, D.R.; Vital, W.F.; Caballero, A.; Tseng, K.Y. CB1 cannabinoid receptor stimulation during adolescence impairs the maturation of GABA function in the adult rat prefrontal cortex. Mol. Psychiatry 2014, 19, 536–543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomases, D.R.; Cass, D.K.; Tseng, K.Y. Periadolescent Exposure to the NMDA Receptor Antagonist MK-801 Impairs the Functional Maturation of Local GABAergic Circuits in the Adult Prefrontal Cortex. J. Neurosci. 2013, 33, 26. [Google Scholar] [CrossRef] [PubMed]
- Nuss, P. Anxiety disorders and GABA neurotransmission: A disturbance of modulation. Neuropsychiatr. Dis. Treat. 2015, 11, 165–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Etkin, A. Functional Neuroanatomy of Anxiety: A Neural Circuit Perspective. In Behavioral Neurobiology of Anxiety and Its Treatment; Stein, M.B., Steckler, T., Eds.; Springer: Berlin/Heidelberg, Germany, 2010; pp. 251–277. [Google Scholar]
- Hashimoto, T.; Nguyen, Q.L.; Rotaru, D.; Keenan, T.; Arion, D.; Beneyto, M.; Gonzalez-Burgos, G.; Lewis, D.A. Protracted developmental trajectories of GABAA receptor alpha1 and alpha2 subunit expression in primate prefrontal cortex. Biol. Psychiatry 2009, 65, 1015–1023. [Google Scholar] [CrossRef] [Green Version]
- Caballero, A.; Thomases, D.R.; Flores-Barrera, E.; Cass, D.K.; Tseng, K.Y. Emergence of GABAergic-dependent regulation of input-specific plasticity in the adult rat prefrontal cortex during adolescence. Psychopharmacology 2014, 231, 1789–1796. [Google Scholar] [CrossRef] [Green Version]
- Bosman, L.W.J.; Heinen, K.; Spijker, S.; Brussaard, A.B. Mice Lacking the Major Adult GABAA Receptor Subtype Have Normal Number of Synapses, But Retain Juvenile IPSC Kinetics Until Adulthood. J. Neurophysiol. 2005, 94, 338–346. [Google Scholar] [CrossRef] [Green Version]
- Caballero, A.; Flores-Barrera, E.; Cass, D.K.; Tseng, K.Y. Differential regulation of parvalbumin and calretinin interneurons in the prefrontal cortex during adolescence. Brain Struct. Funct. 2014, 219, 395–406. [Google Scholar] [CrossRef] [Green Version]
- Stevens, H.E.; Su, T.; Yanagawa, Y.; Vaccarino, F.M. Prenatal stress delays inhibitory neuron progenitor migration in the developing neocortex. Psychoneuroendocrinology 2013, 38, 509–521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gumusoglu, S.B.; Fine, R.S.; Murray, S.J.; Bittle, J.L.; Stevens, H.E. The role of IL-6 in neurodevelopment after prenatal stress. Brain Behav. Immun. 2017, 65, 274–283. [Google Scholar] [CrossRef] [PubMed]
- Canetta, S.; Bolkan, S.; Padilla-Coreano, N.; Song, L.J.; Sahn, R.; Harrison, N.L.; Gordon, J.A.; Brown, A.; Kellendonk, C. Maternal immune activation leads to selective functional deficits in offspring parvalbumin interneurons. Mol. Psychiatry 2016, 21, 956–968. [Google Scholar] [CrossRef] [PubMed]
- Eyles, D.W.; McGrath, J.J.; Reynolds, G.P. Neuronal calcium-binding proteins and schizophrenia. Schizophr. Res. 2002, 57, 27–34. [Google Scholar] [CrossRef]
- Ferguson, B.R.; Gao, W.-J. PV Interneurons: Critical Regulators of E/I Balance for Prefrontal Cortex-Dependent Behavior and Psychiatric Disorders. Front. Neural Circuits 2018, 12, 37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McNally, J.M.; McCarley, R.W. Gamma band oscillations: A key to understanding schizophrenia symptoms and neural circuit abnormalities. Curr. Opin. Psychiatry 2016, 29, 202–210. [Google Scholar] [CrossRef] [Green Version]
- Kiran, C.; Chaudhury, S. Prevalence of comorbid anxiety disorders in schizophrenia. Ind. Psychiatry J. 2016, 25, 35–40. [Google Scholar] [CrossRef]
- Fine, R.; Zhang, J.; Stevens, H.E. Prenatal stress and inhibitory neuron systems: Implications for neuropsychiatric disorders. Mol. Psychiatry 2014, 19, 641–651. [Google Scholar] [CrossRef] [PubMed]
- Schulte, J.T.; Wierenga, C.J.; Bruining, H. Chloride transporters and GABA polarity in developmental, neurological and psychiatric conditions. Neurosci. Biobehav. Rev. 2018, 90, 260–271. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Lovinger, D.; Delpire, E. Cortical Neurons Lacking KCC2 Expression Show Impaired Regulation of Intracellular Chloride. J. Neurophysiol. 2005, 93, 1557–1568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tornberg, J.; Voikar, V.; Savilahti, H.; Rauvala, H.; Airaksinen, M.S. Behavioural phenotypes of hypomorphic KCC2-deficient mice. Eur. J. Neurosci. 2005, 21, 1327–1337. [Google Scholar] [CrossRef] [PubMed]
- Chamma, I.; Chevy, Q.; Poncer, J.C.; Levi, S. Role of the neuronal K-Cl co-transporter KCC2 in inhibitory and excitatory neurotransmission. Front. Cell. Neurosci. 2012, 6, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roussa, E.; Speer, J.M.; Chudotvorova, I.; Khakipoor, S.; Smirnov, S.; Rivera, C.; Krieglstein, K. The membrane trafficking and functionality of the K+-Cl− co-transporter KCC2 is regulated by TGF-β2. J. Cell Sci. 2016, 129, 3485–3498. [Google Scholar] [CrossRef] [PubMed] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Suwaluk, A.; Chutabhakdikul, N. Altered Development of Prefrontal GABAergic Functions and Anxiety-like Behavior in Adolescent Offspring Induced by Prenatal Stress. Brain Sci. 2022, 12, 1015. https://doi.org/10.3390/brainsci12081015
Suwaluk A, Chutabhakdikul N. Altered Development of Prefrontal GABAergic Functions and Anxiety-like Behavior in Adolescent Offspring Induced by Prenatal Stress. Brain Sciences. 2022; 12(8):1015. https://doi.org/10.3390/brainsci12081015
Chicago/Turabian StyleSuwaluk, Arbthip, and Nuanchan Chutabhakdikul. 2022. "Altered Development of Prefrontal GABAergic Functions and Anxiety-like Behavior in Adolescent Offspring Induced by Prenatal Stress" Brain Sciences 12, no. 8: 1015. https://doi.org/10.3390/brainsci12081015