
Gene Expression Changes of Murine Cortex Homeostasis in Response to Sleep Deprivation Hint Dysregulated Aging-like Transcriptional Responses

 ,
,  , ,
, , {kind=link}
{kind=link}
Abstract
:1. Introduction
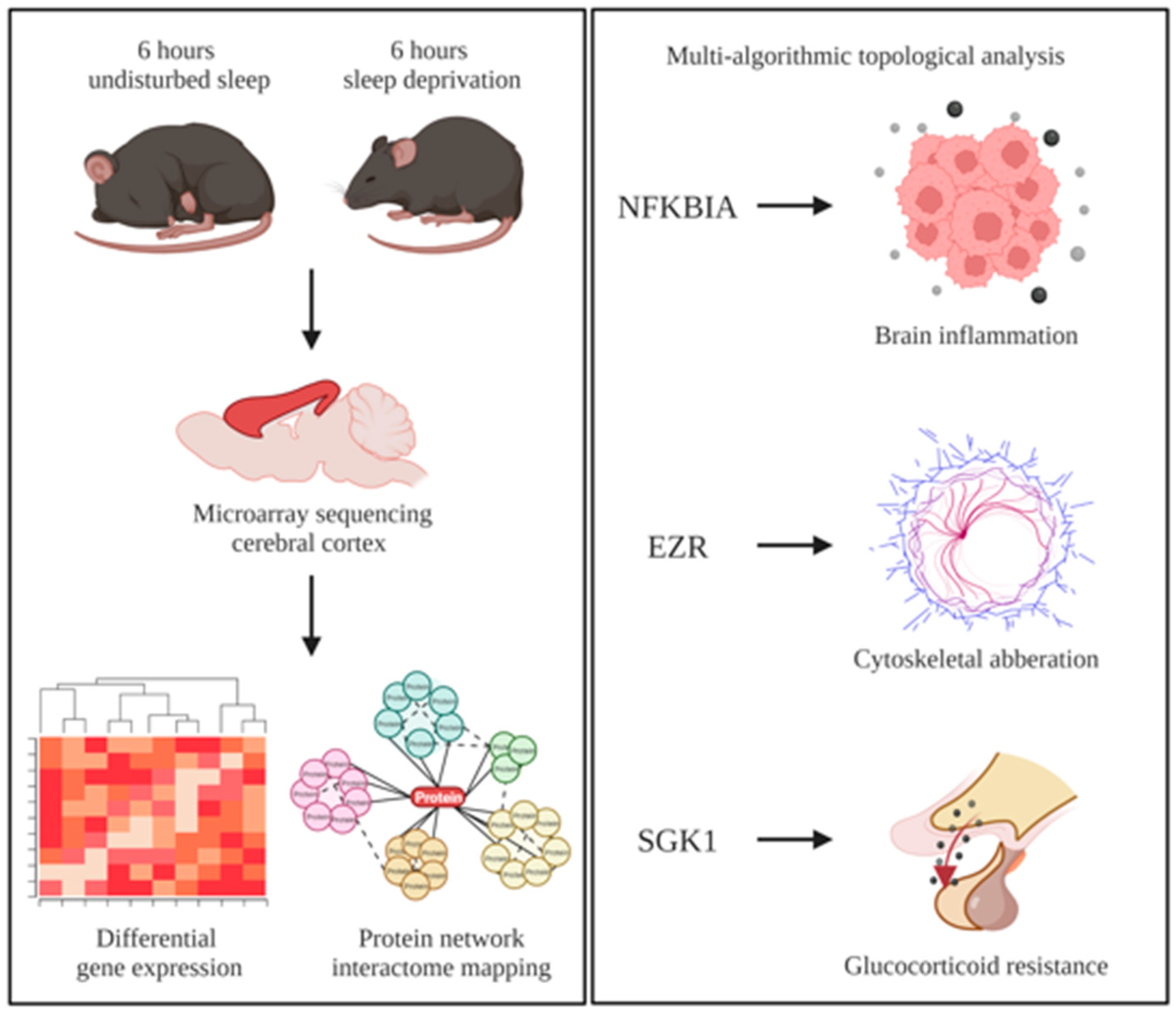
2. Methods
2.1. Collection of Microarray Datasets
2.2. Identification of Differentially Expressed Genes
2.3. Construction of Protein–Protein Interaction Networks
2.4. Identification of Regulatory Differentially Expressed Genes
3. Results
3.1. Overview of Microarray Datasets
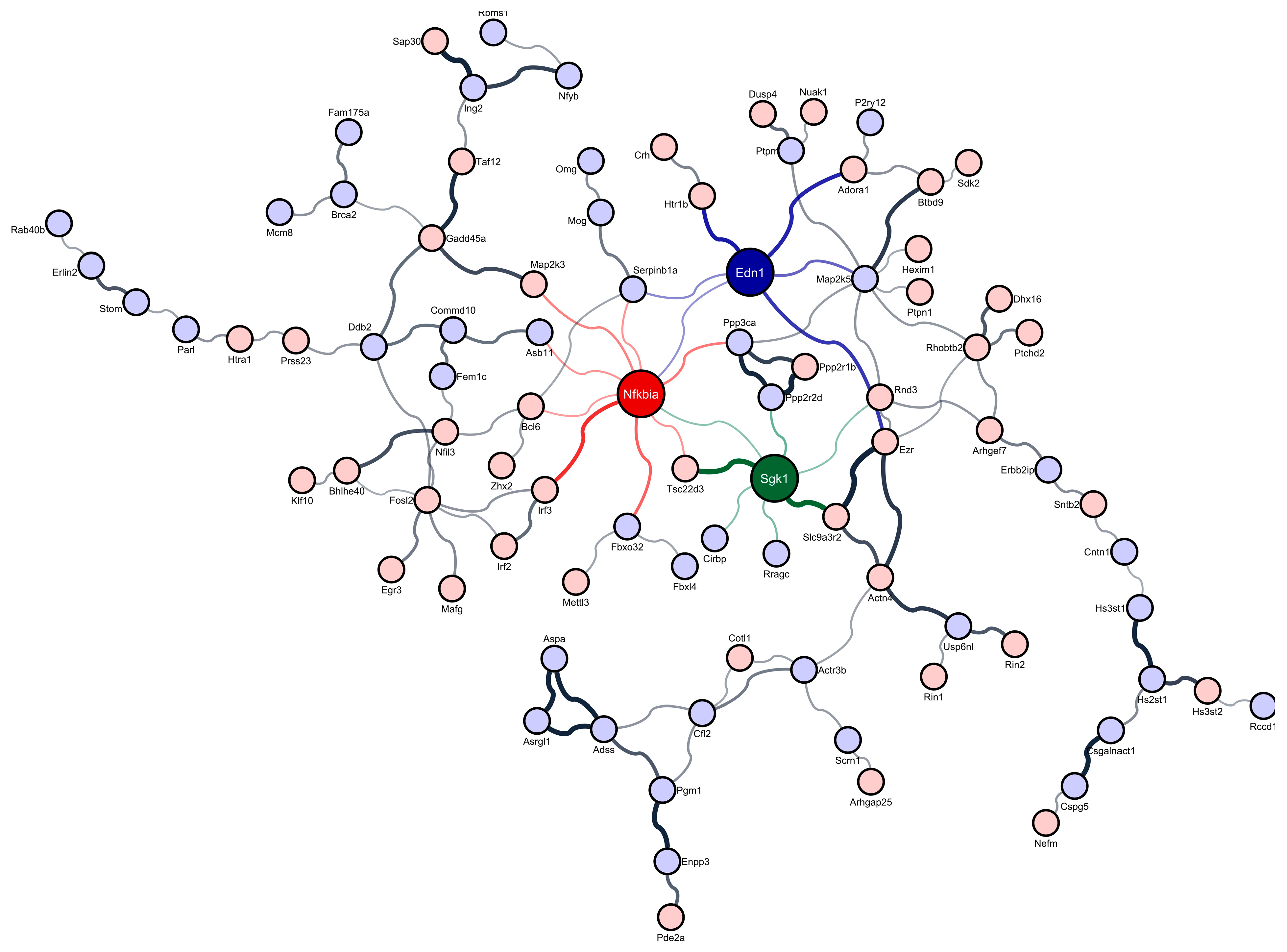
3.2. Regulatory Differentially Expressed Genes in Sleep-Deprived Murine Cortex
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dinges, D.F.; Pack, F.; Williams, K.; Gillen, K.A.; Powell, J.W.; Ott, G.E.; Aptowicz, C.; Pack, A.I. Cumulative sleepiness, mood disturbance, and psychomotor vigilance performance decrements during a week of sleep restricted to 4–5 hours per night. Sleep 1997, 20, 267–277. [Google Scholar]
- Mollicone, D.J.; Van Dongen, H.; Rogers, N.L.; Banks, S.; Dinges, D.F. Time of day effects on neurobehavioral performance during chronic sleep restriction. Aviat. Space Environ. Med. 2010, 81, 735–744. [Google Scholar] [CrossRef]
- Jiang, F.; VanDyke, R.D.; Zhang, J.; Li, F.; Gozal, D.; Shen, X. Effect of chronic sleep restriction on sleepiness and working memory in adolescents and young adults. J. Clin. Exp. Neuropsychol. 2011, 33, 892–900. [Google Scholar] [CrossRef]
- Heaney, J.; Phillips, A. Aging, Health Behaviors, and the Diurnal Rhythm and Awakening Response of Salivary Cortisol. Exp. Aging Res. 2012, 38, 295–314. [Google Scholar] [CrossRef] [Green Version]
- Lo, J.C.; Bennion, K.A.; Chee, M.W. Sleep restriction can attenuate prioritization benefits on declarative memory consolidation. J. Sleep Res. 2016, 25, 664–672. [Google Scholar] [CrossRef]
- Chen, L.; Huang, J.; Yang, L.; Zeng, X.-A.; Zhang, Y.; Wang, X.; Chen, M.; Li, X.; Zhang, Y.; Zhang, M. Sleep deprivation accelerates the progression of Alzheimer’s disease by influencing Aβ-related metabolism. Neurosci. Lett. 2017, 650, 146–152. [Google Scholar] [CrossRef]
- Simor, A.; Györffy, B.A.; Gulyássy, P.; Völgyi, K.; Tóth, V.; Todorov, M.I.; Kis, V.; Borhegyi, Z.; Szabó, Z.; Janáky, T. The short-and long-term proteomic effects of sleep deprivation on the cortical and thalamic synapses. Mol. Cell. Neurosci. 2017, 79, 64–80. [Google Scholar] [CrossRef] [Green Version]
- Ba, L.; Huang, L.; He, Z.; Deng, S.; Xie, Y.; Zhang, M.; Jacob, C.; Antonecchia, E.; Liu, Y.; Xiao, W. Does Chronic Sleep Fragmentation Lead to Alzheimer’s Disease in Young Wild-Type Mice? Front. Aging Neurosci. 2021, 13, 759983. [Google Scholar] [CrossRef]
- Naidoo, N.; Giang, W.; Galante, R.J.; Pack, A.I. Sleep deprivation induces the unfolded protein response in mouse cerebral cortex. J. Neurochem. 2005, 92, 1150–1157. [Google Scholar] [CrossRef]
- Grønli, J.; Soulé, J.; Bramham, C.R. Sleep and protein synthesis-dependent synaptic plasticity: Impacts of sleep loss and stress. Front. Behav. Neurosci. 2014, 7, 224. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Lai, C.S.W.; Bai, Y.; Li, W.; Zhao, R.; Yang, G.; Frank, M.G.; Gan, W.-B. REM sleep promotes experience-dependent dendritic spine elimination in the mouse cortex. Nat. Commun. 2020, 11, 4819. [Google Scholar] [CrossRef]
- Tudor, J.C.; Davis, E.J.; Peixoto, L.; Wimmer, M.E.; van Tilborg, E.; Park, A.J.; Poplawski, S.G.; Chung, C.W.; Havekes, R.; Huang, J. Sleep deprivation impairs memory by attenuating mTORC1-dependent protein synthesis. Sci. Signal. 2016, 9, ra41. [Google Scholar] [CrossRef] [Green Version]
- Kam, K.; Kang, M.; Eren, C.Y.; Pettibone, W.D.; Bowling, H.; Taveras, S.; Ly, A.; Chen, R.K.; Berryman, N.V.; Klann, E. Interactions between sleep disruption, motor learning, and p70 S6 kinase 1 signaling. Sleep 2020, 43, zsz244. [Google Scholar] [CrossRef]
- Alhola, P.; Polo-Kantola, P. Sleep deprivation: Impact on cognitive performance. Neuropsychiatr. Dis. Treat. 2007, 3, 553–567. [Google Scholar]
- Mander, B.A.; Winer, J.R.; Walker, M.P. Sleep and human aging. Neuron 2017, 94, 19–36. [Google Scholar] [CrossRef] [Green Version]
- Millstein, J.; Winrow, C.J.; Kasarskis, A.; Owens, J.R.; Zhou, L.; Summa, K.C.; Fitzpatrick, K.; Zhang, B.; Vitaterna, M.H.; Schadt, E.E. Identification of causal genes, networks, and transcriptional regulators of REM sleep and wake. Sleep 2011, 34, 1469–1477. [Google Scholar] [CrossRef] [Green Version]
- Gaine, M.E.; Bahl, E.; Chatterjee, S.; Michaelson, J.J.; Abel, T.; Lyons, L.C. Altered hippocampal transcriptome dynamics following sleep deprivation. Mol. Brain 2021, 14, 125. [Google Scholar] [CrossRef]
- Krause, A.J.; Simon, E.B.; Mander, B.A.; Greer, S.M.; Saletin, J.M.; Goldstein-Piekarski, A.N.; Walker, M.P. The sleep-deprived human brain. Nat. Rev. Neurosci. 2017, 18, 404–418. [Google Scholar] [CrossRef]
- Sexton, C.E.; Storsve, A.B.; Walhovd, K.B.; Johansen-Berg, H.; Fjell, A.M. Poor sleep quality is associated with increased cortical atrophy in community-dwelling adults. Neurology 2014, 83, 967–973. [Google Scholar]
- Toro-Domínguez, D.; Martorell-Marugán, J.; López-Domínguez, R.; García-Moreno, A.; González-Rumayor, V.; Alarcón-Riquelme, M.E.; Carmona-Sáez, P. ImaGEO: Integrative gene expression meta-analysis from GEO database. Bioinformatics 2019, 35, 880–882. [Google Scholar] [CrossRef]
- Giannos, P.; Triantafyllidis, K.K.; Giannos, G.; Kechagias, K.S. SPP1 in infliximab resistant ulcerative colitis and associated colorectal cancer: An analysis of differentially expressed genes. Eur. J. Gastroenterol. Hepatol. 2022, 34, 598–606. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P. STRING v11: Protein–protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef] [Green Version]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Chin, C.-H.; Chen, S.-H.; Wu, H.-H.; Ho, C.-W.; Ko, M.-T.; Lin, C.-Y. cytoHubba: Identifying hub objects and sub-networks from complex interactome. BMC Syst. Biol. 2014, 8, S11. [Google Scholar] [CrossRef] [Green Version]
- Giannos, P.; Prokopidis, K. Gene Expression Profiles of the Aging Rat Hippocampus Imply Altered Immunoglobulin Dynamics. Front. Neurosci. 2022, 16, 915907. [Google Scholar] [CrossRef]
- Mackiewicz, M.; Shockley, K.R.; Romer, M.A.; Galante, R.J.; Zimmerman, J.E.; Naidoo, N.; Baldwin, D.A.; Jensen, S.T.; Churchill, G.A.; Pack, A.I. Macromolecule biosynthesis: A key function of sleep. Physiol. Genom. 2007, 31, 441–457. [Google Scholar] [CrossRef]
- Gerstner, J.R.; Koberstein, J.N.; Watson, A.J.; Zapero, N.; Risso, D.; Speed, T.P.; Frank, M.G.; Peixoto, L. Removal of unwanted variation reveals novel patterns of gene expression linked to sleep homeostasis in murine cortex. BMC Genom. 2016, 17, 377–387. [Google Scholar] [CrossRef] [Green Version]
- Hinard, V.; Mikhail, C.; Pradervand, S.; Curie, T.; Houtkooper, R.H.; Auwerx, J.; Franken, P.; Tafti, M. Key electrophysiological, molecular, and metabolic signatures of sleep and wakefulness revealed in primary cortical cultures. J. Neurosci. 2012, 32, 12506–12517. [Google Scholar] [CrossRef] [Green Version]
- Albensi, B.C. What is nuclear factor kappa B (NF-κB) doing in and to the mitochondrion? Front. Cell Dev. Biol. 2019, 7, 154. [Google Scholar] [CrossRef]
- Sharman, E.H.; Bondy, S.C.; Sharman, K.G.; Lahiri, D.; Cotman, C.W.; Perreau, V.M. Effects of melatonin and age on gene expression in mouse CNS using microarray analysis. Neurochem. Int. 2007, 50, 336–344. [Google Scholar] [CrossRef] [Green Version]
- Ng, K.Y.; Leong, M.K.; Liang, H.; Paxinos, G. Melatonin receptors: Distribution in mammalian brain and their respective putative functions. Brain Struct. Funct. 2017, 222, 2921–2939. [Google Scholar] [CrossRef]
- Muxel, S.M.; Pires-Lapa, M.A.; Monteiro, A.W.A.; Cecon, E.; Tamura, E.K.; Floeter-Winter, L.M.; Markus, R.P. NF-κB drives the synthesis of melatonin in RAW 264.7 macrophages by inducing the transcription of the arylalkylamine-N-acetyltransferase (AA-NAT) gene. PLoS ONE 2012, 7, e52010. [Google Scholar] [CrossRef] [Green Version]
- Irwin, M.R.; Wang, M.; Ribeiro, D.; Cho, H.J.; Olmstead, R.; Breen, E.C.; Martinez-Maza, O.; Cole, S. Sleep loss activates cellular inflammatory signaling. Biol. Psychiatry 2008, 64, 538–540. [Google Scholar] [CrossRef] [Green Version]
- Fleischman, D.A.; Arfanakis, K.; Kelly, J.F.; Rajendran, N.; Buchman, A.S.; Morris, M.C.; Barnes, L.L.; Bennett, D.A. Regional cortical thinning and systemic inflammation in older persons without dementia. J. Am. Geriatr. Soc. 2010, 58, 1823. [Google Scholar] [CrossRef]
- Kaur, S.S.; Gonzales, M.M.; Eagan, D.E.; Goudarzi, K.; Tanaka, H.; Haley, A.P. Inflammation as a mediator of the relationship between cortical thickness and metabolic syndrome. Brain Imaging Behav. 2015, 9, 737–743. [Google Scholar] [CrossRef] [Green Version]
- Arena, A.; Zimmer, T.S.; van Scheppingen, J.; Korotkov, A.; Anink, J.J.; Mühlebner, A.; Jansen, F.E.; van Hecke, W.; Spliet, W.G.; van Rijen, P.C. Oxidative stress and inflammation in a spectrum of epileptogenic cortical malformations: Molecular insights into their interdependence. Brain Pathol. 2019, 29, 351–365. [Google Scholar] [CrossRef]
- Prats-Soteras, X.; Jurado, M.; Ottino-González, J.; García-García, I.; Segura, B.; Caldú, X.; Sánchez-Garre, C.; Miró, N.; Tor, C.; Sender-Palacios, M. Inflammatory agents partially explain associations between cortical thickness, surface area, and body mass in adolescents and young adulthood. Int. J. Obes. 2020, 44, 1487–1496. [Google Scholar] [CrossRef]
- De Pablos, R.; Villaran, R.; Argüelles, S.; Herrera, A.; Venero, J.; Ayala, A.; Cano, J.; Machado, A. Stress increases vulnerability to inflammation in the rat prefrontal cortex. J. Neurosci. 2006, 26, 5709–5719. [Google Scholar] [CrossRef]
- Dahlke, C.; Saberi, D.; Ott, B.; Brand-Saberi, B.; Schmitt-John, T.; Theiss, C. Inflammation and neuronal death in the motor cortex of the wobbler mouse, an ALS animal model. J. Neuroinflamm. 2015, 12, 215. [Google Scholar] [CrossRef] [Green Version]
- Williams, J.A.; Sathyanarayanan, S.; Hendricks, J.C.; Sehgal, A. Interaction between sleep and the immune response in Drosophila: A role for the NFκB relish. Sleep 2007, 30, 389–400. [Google Scholar] [CrossRef]
- Amato, R.; D’Antona, L.; Porciatti, G.; Agosti, V.; Menniti, M.; Rinaldo, C.; Costa, N.; Bellacchio, E.; Mattarocci, S.; Fuiano, G. Sgk1 activates MDM2-dependent p53 degradation and affects cell proliferation, survival, and differentiation. J. Mol. Med. 2009, 87, 1221–1239. [Google Scholar] [CrossRef]
- Sarabdjitsingh, R.A.; Isenia, S.; Polman, A.; Mijalkovic, J.; Lachize, S.; Datson, N.; de Kloet, E.R.; Meijer, O.C. Disrupted corticosterone pulsatile patterns attenuate responsiveness to glucocorticoid signaling in rat brain. Endocrinology 2010, 151, 1177–1186. [Google Scholar] [CrossRef]
- Ma, Y.L.; Tsai, M.C.; Hsu, W.L.; Lee, E.H. SGK protein kinase facilitates the expression of long-term potentiation in hippocampal neurons. Learn. Mem. 2006, 13, 114–118. [Google Scholar] [CrossRef] [Green Version]
- Miyata, S.; Yoshikawa, K.; Taniguchi, M.; Ishikawa, T.; Tanaka, T.; Shimizu, S.; Tohyama, M. Sgk1 regulates desmoglein 1 expression levels in oligodendrocytes in the mouse corpus callosum after chronic stress exposure. Biochem. Biophys. Res. Commun. 2015, 464, 76–82. [Google Scholar] [CrossRef]
- Tai, D.J.; Su, C.-C.; Ma, Y.-L.; Lee, E.H. SGK1 phosphorylation of IB kinase and p300 up-regulates NF-B activity and increases N-methyl-D-aspartate receptor NR2A and NR2B expression. J. Biol. Chem. 2009, 284, 4073–4089. [Google Scholar] [CrossRef] [Green Version]
- Kino, T. Stress, glucocorticoid hormones, and hippocampal neural progenitor cells: Implications to mood disorders. Front. Physiol. 2015, 6, 230. [Google Scholar] [CrossRef] [Green Version]
- Sato, H.; Horikawa, Y.; Iizuka, K.; Sakurai, N.; Tanaka, T.; Shihara, N.; Oshima, A.; Takeda, J.; Mikuni, M. Large-scale analysis of glucocorticoid target genes in rat hypothalamus. J. Neurochem. 2008, 106, 805–814. [Google Scholar] [CrossRef]
- Anacker, C.; Cattaneo, A.; Musaelyan, K.; Zunszain, P.A.; Horowitz, M.; Molteni, R.; Luoni, A.; Calabrese, F.; Tansey, K.; Gennarelli, M. Role for the kinase SGK1 in stress, depression, and glucocorticoid effects on hippocampal neurogenesis. Proc. Natl. Acad. Sci. USA 2013, 110, 8708–8713. [Google Scholar] [CrossRef] [Green Version]
- Jin, J.; Kim, S.-N.; Liu, X.; Zhang, H.; Zhang, C.; Seo, J.-S.; Kim, Y.; Sun, T. miR-17-92 cluster regulates adult hippocampal neurogenesis, anxiety, and depression. Cell Rep. 2016, 16, 1653–1663. [Google Scholar] [CrossRef] [Green Version]
- Licznerski, P.; Duric, V.; Banasr, M.; Alavian, K.N.; Ota, K.T.; Kang, H.J.; Jonas, E.A.; Ursano, R.; Krystal, J.H.; Duman, R.S. Decreased SGK1 expression and function contributes to behavioral deficits induced by traumatic stress. PLoS Biol. 2015, 13, e1002282. [Google Scholar] [CrossRef] [Green Version]
- Webster, M.K.; Goya, L.; Ge, Y.; Maiyar, A.; Firestone, G. Characterization of sgk, a novel member of the serine/threonine protein kinase gene family which is transcriptionally induced by glucocorticoids and serum. Mol. Cell. Biol. 1993, 13, 2031–2040. [Google Scholar]
- Lang, F.; Artunc, F.; Vallon, V. The physiological impact of the serum-and glucocorticoid-inducible kinase SGK1. Curr. Opin. Nephrol. Hypertens. 2009, 18, 439. [Google Scholar] [CrossRef]
- Conti, B.; Maier, R.; Barr, A.; Morale, M.; Lu, X.; Sanna, P.; Bilbe, G.; Hoyer, D.; Bartfai, T. Region-specific transcriptional changes following the three antidepressant treatments electro convulsive therapy, sleep deprivation and fluoxetine. Mol. Psychiatry 2007, 12, 167–189. [Google Scholar] [CrossRef]
- Elliott, A.S.; Huber, J.D.; O’Callaghan, J.P.; Rosen, C.L.; Miller, D.B. A review of sleep deprivation studies evaluating the brain transcriptome. SpringerPlus 2014, 3, 728. [Google Scholar] [CrossRef] [Green Version]
- Maret, S.; Dorsaz, S.; Gurcel, L.; Pradervand, S.; Petit, B.; Pfister, C.; Hagenbuchle, O.; O’Hara, B.F.; Franken, P.; Tafti, M. Homer1a is a core brain molecular correlate of sleep loss. Proc. Natl. Acad. Sci. USA 2007, 104, 20090–20095. [Google Scholar] [CrossRef] [Green Version]
- Cirelli, C.; Tononi, G. Gene expression in the brain across the sleep–waking cycle. Brain Res. 2000, 885, 303–321. [Google Scholar] [CrossRef]
- Cirelli, C. Invited Review: How sleep deprivation affects gene expression in the brain: A review of recent findings. J. Appl. Physiol. 2002, 92, 394–400. [Google Scholar] [CrossRef] [Green Version]
- Cirelli, C.; Faraguna, U.; Tononi, G. Changes in brain gene expression after long-term sleep deprivation. J. Neurochem. 2006, 98, 1632–1645. [Google Scholar] [CrossRef]
- Hwang, D.Y.; Golby, A.J. The brain basis for episodic memory: Insights from functional MRI, intracranial EEG, and patients with epilepsy. Epilepsy Behav. 2006, 8, 115–126. [Google Scholar] [CrossRef]
- Drummond, S.P.; Brown, G.G.; Stricker, J.L.; Buxton, R.B.; Wong, E.C.; Gillin, J.C. Sleep deprivation-induced reduction in cortical functional response to serial subtraction. Neuroreport 1999, 10, 3745–3748. [Google Scholar] [CrossRef] [Green Version]
- Kelly, A.C.; Hester, R.; Murphy, K.; Javitt, D.C.; Foxe, J.J.; Garavan, H. Prefrontal-subcortical dissociations underlying inhibitory control revealed by event-related fMRI. Eur. J. Neurosci. 2004, 19, 3105–3112. [Google Scholar] [CrossRef]
- Thomas, M.; Sing, H.; Belenky, G.; Holcomb, H.; Mayberg, H.; Dannals, R.; Wagner Jr, H.; Thorne, D.; Popp, K.; Rowland, L. Neural basis of alertness and cognitive performance impairments during sleepiness. I. Effects of 24 h of sleep deprivation on waking human regional brain activity. J. Sleep Res. 2000, 9, 335–352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamasaki, H.; LaBar, K.S.; McCarthy, G. Dissociable prefrontal brain systems for attention and emotion. In Social Neuroscience; Psychology Press: London, UK, 2013; pp. 43–51. [Google Scholar]
- Levy, M.J.; Boulle, F.; Steinbusch, H.W.; van den Hove, D.L.; Kenis, G.; Lanfumey, L. Neurotrophic factors and neuroplasticity pathways in the pathophysiology and treatment of depression. Psychopharmacology 2018, 235, 2195–2220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dattilo, V.; Amato, R.; Perrotti, N.; Gennarelli, M. The emerging role of SGK1 (serum-and glucocorticoid-regulated kinase 1) in major depressive disorder: Hypothesis and mechanisms. Front. Genet. 2020, 11, 826. [Google Scholar] [CrossRef]
- Lavialle, M.; Aumann, G.; Anlauf, E.; Pröls, F.; Arpin, M.; Derouiche, A. Structural plasticity of perisynaptic astrocyte processes involves ezrin and metabotropic glutamate receptors. Proc. Natl. Acad. Sci. USA 2011, 108, 12915–12919. [Google Scholar] [CrossRef] [Green Version]
- Fehon, R.G.; McClatchey, A.I.; Bretscher, A. Organizing the cell cortex: The role of ERM proteins. Nat. Rev. Mol. Cell Biol. 2010, 11, 276–287. [Google Scholar] [CrossRef] [Green Version]
- Rouven Brückner, B.; Pietuch, A.; Nehls, S.; Rother, J.; Janshoff, A. Ezrin is a major regulator of membrane tension in epithelial cells. Sci. Rep. 2015, 5, 14700. [Google Scholar] [CrossRef] [Green Version]
- Moon, Y.; Kim, J.Y.; Choi, S.Y.; Cho, H.M.; Kim, H.; Sun, W. Expression of ezrin in subventricular zone neural stem cells and their progeny in adult and developing mice. Histochem. Cell Biol. 2013, 139, 403–413. [Google Scholar] [CrossRef]
- Lööv, C.; Nadadhur, A.G.; Hillered, L.; Clausen, F.; Erlandsson, A. Extracellular ezrin: A novel biomarker for traumatic brain injury. J. Neurotrauma 2015, 32, 244–251. [Google Scholar] [CrossRef]
- Vega, I.E.; Umstead, A.; Wygant, C.M.; Beck, J.S.; Counts, S.E. Ezrin Expression is Increased During Disease Progression in a Tauopathy Mouse Model and Alzheimer’s Disease. Curr. Alzheimer Res. 2018, 15, 1086–1095. [Google Scholar] [CrossRef]
- Tang, L.; Wang, X.; Wu, J.; Zhang, Z.; Wu, S.; Su, T.; Lin, Z.; Chen, X.; Liao, X.; Bai, T. Sleep deprivation induces dry eye through inhibition of PPARα expression in corneal epithelium. Investig. Ophthalmol. Vis. Sci. 2018, 59, 5494–5508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bellesi, M.; de Vivo, L.; Tononi, G.; Cirelli, C. Effects of sleep and wake on astrocytes: Clues from molecular and ultrastructural studies. BMC Biol. 2015, 13, 66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minakawa, E.N.; Wada, K.; Nagai, Y. Sleep disturbance as a potential modifiable risk factor for Alzheimer’s disease. Int. J. Mol. Sci. 2019, 20, 803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shamim, S.A.; Warriach, Z.I.; Tariq, M.A.; Rana, K.F.; Malik, B.H. Insomnia: Risk factor for neurodegenerative diseases. Cureus 2019, 11, e6004. [Google Scholar] [CrossRef] [Green Version]
- Pawlyk, A.C.; Ferber, M.; Shah, A.; Pack, A.I.; Naidoo, N. Proteomic analysis of the effects and interactions of sleep deprivation and aging in mouse cerebral cortex. J. Neurochem. 2007, 103, 2301–2313. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Giannos, P.; Prokopidis, K.; Forbes, S.C.; Celoch, K.; Candow, D.G.; Tartar, J.L. Gene Expression Changes of Murine Cortex Homeostasis in Response to Sleep Deprivation Hint Dysregulated Aging-like Transcriptional Responses. Brain Sci. 2022, 12, 825. https://doi.org/10.3390/brainsci12070825
Giannos P, Prokopidis K, Forbes SC, Celoch K, Candow DG, Tartar JL. Gene Expression Changes of Murine Cortex Homeostasis in Response to Sleep Deprivation Hint Dysregulated Aging-like Transcriptional Responses. Brain Sciences. 2022; 12(7):825. https://doi.org/10.3390/brainsci12070825
Chicago/Turabian StyleGiannos, Panagiotis, Konstantinos Prokopidis, Scott C. Forbes, Kamil Celoch, Darren G. Candow, and Jaime L. Tartar. 2022. "Gene Expression Changes of Murine Cortex Homeostasis in Response to Sleep Deprivation Hint Dysregulated Aging-like Transcriptional Responses" Brain Sciences 12, no. 7: 825. https://doi.org/10.3390/brainsci12070825