
High Estrogen Levels Cause Greater Leg Muscle Fatigability in Eumenorrheic Young Women after 4 mA Transcranial Direct Current Stimulation



Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects
2.2. Study Design
2.3. Isokinetic/Isometric Strength Testing
2.4. Isokinetic Fatigue Task (FT)
2.5. Electromyography (EMG)
2.6. Transcranial Direct Current Stimulation (tDCS)
2.7. Blood Draw
2.8. Data Analysis
2.9. Statistical Analysis
3. Results
3.1. Subject Characteristics
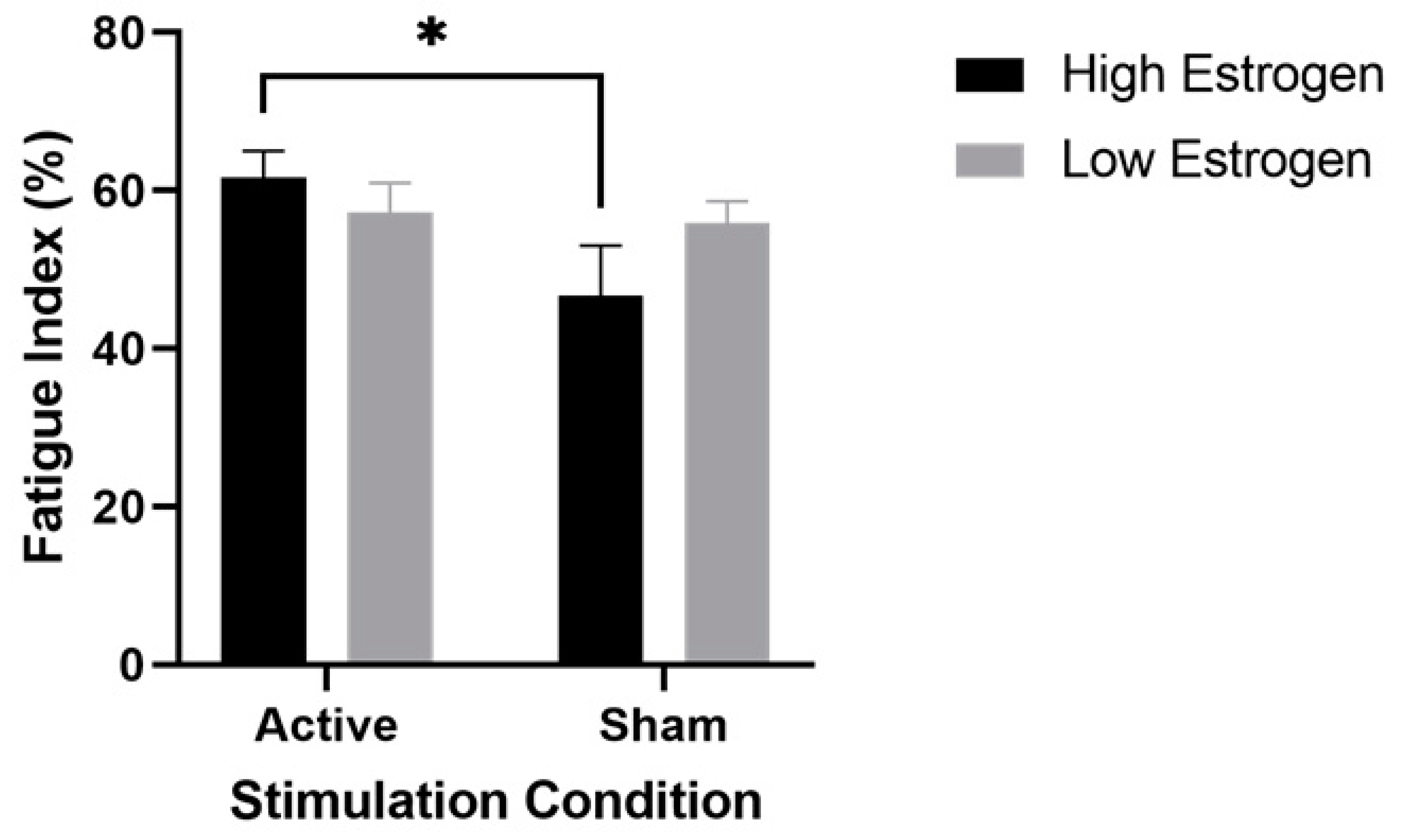
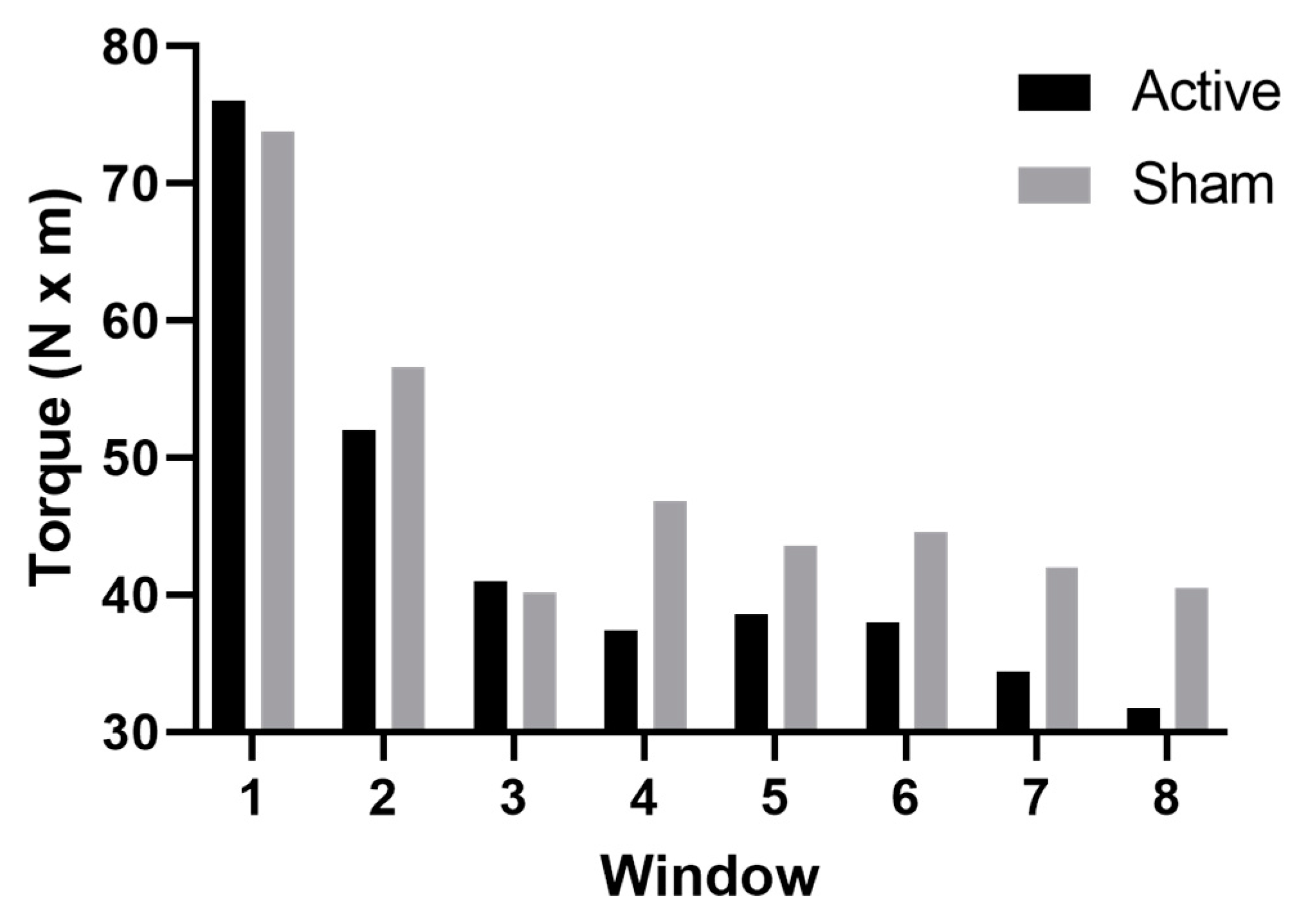
3.2. Muscle Fatigability
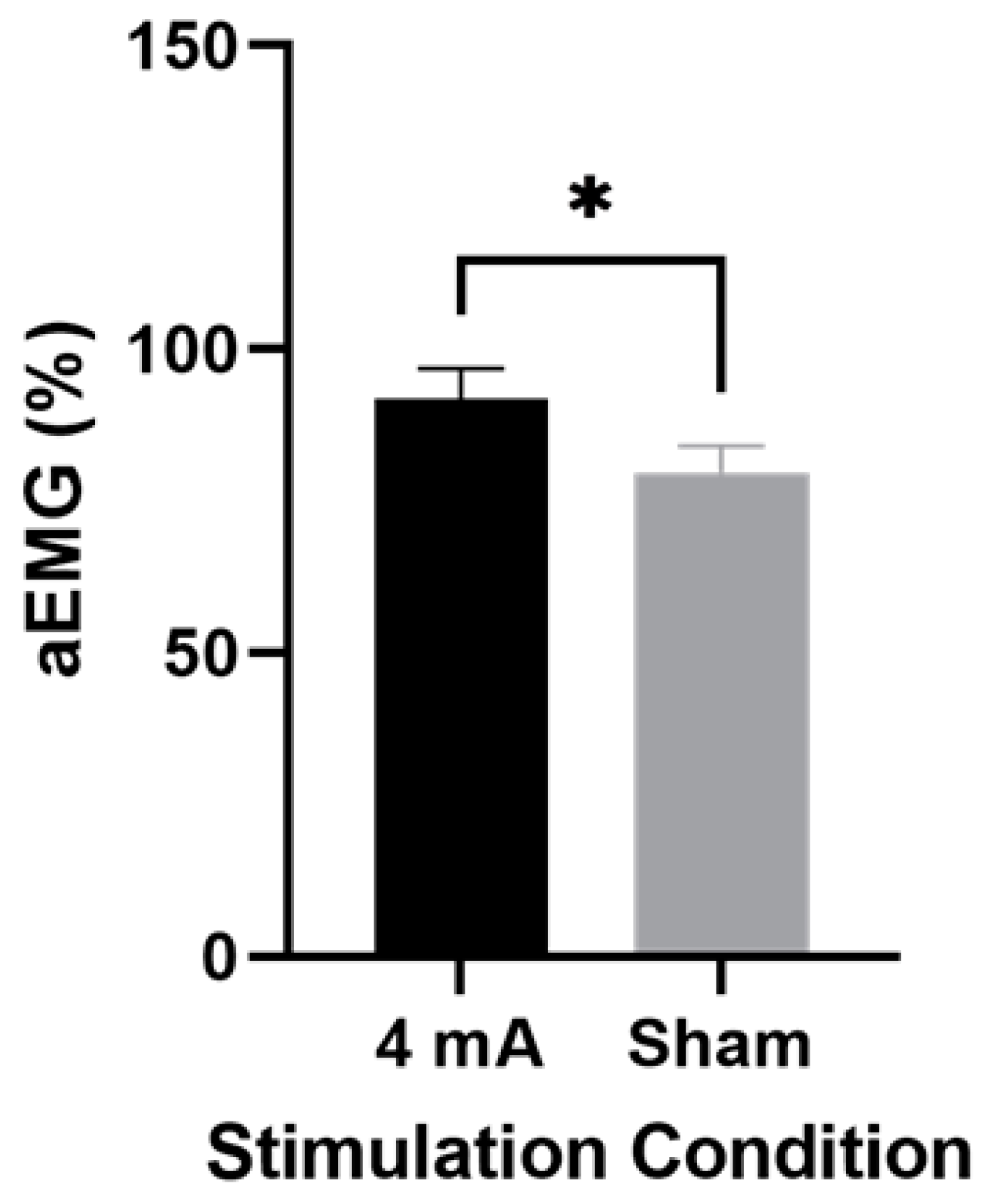
3.3. Muscle Activity
3.4. tDCS Tolerability and Blinding
4. Discussion
4.1. High Estrogen and Active Stimulation in Performance Fatigability
4.2. Muscle Activity in Active Stimulation vs. Sham
4.3. Crossover Effect
4.4. Limitations and Future Studies
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kuo, M.-F.; Paulus, W.; Nitsche, M.A. Sex differences in cortical neuroplasticity in humans. NeuroReport 2006, 17, 1703–1707. [Google Scholar] [CrossRef] [PubMed]
- Chaieb, L.; Antal, A.; Paulus, W. Gender-specific modulation of short-term neuroplasticity in the visual cortex induced by transcranial direct current stimulation. Vis. Neurosci. 2008, 25, 77–81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fregni, F.; Boggio, P.S.; Nitsche, M.; Bermpohl, F.; Antal, A.; Feredoes, E.; Marcolin, M.A.; Rigonatti, S.P.; Silva, M.T.; Paulus, W.; et al. Anodal transcranial direct current stimulation of prefrontal cortex enhances working memory. Exp. Brain Res. 2005, 166, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Inghilleri, M.; Conte, A.; Currà, A.; Frasca, V.; Lorenzano, C.; Berardelli, A. Ovarian hormones and cortical excitability. An rTMS study in humans. Clin. Neurophysiol. 2004, 115, 1063–1068. [Google Scholar] [CrossRef] [PubMed]
- Smith, M.J.; Adams, L.F.; Schmidt, P.J.; Rubinow, D.R.; Wassermann, E.M. Effects of ovarian hormones on human cortical excitability. Ann. Neurol. 2002, 51, 599–603. [Google Scholar] [CrossRef] [PubMed]
- Dedoncker, J.; Brunoni, A.R.; Baeken, C.; Vanderhasselt, M.A. A Systematic Review and Meta-Analysis of the Effects of Transcranial Direct Current Stimulation (tDCS) Over the Dorsolateral Prefrontal Cortex in Healthy and Neuropsychiatric Samples: Influence of Stimulation Parameters. Brain Stimul. 2016, 9, 501–517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rudroff, T.; Workman, C.D.; Fietsam, A.C.; Kamholz, J. Response Variability in Transcranial Direct Current Stimulation: Why Sex Matters. Front. Psychiatry 2020, 11, 585. [Google Scholar] [CrossRef]
- Horvath, J.C.; Carter, O.; Forte, J.D. No significant effect of transcranial direct current stimulation (tDCS) found on simple motor reaction time comparing 15 different simulation protocols. Neuropsychologia 2016, 91, 544–552. [Google Scholar] [CrossRef]
- Horvath, J.C.; Vogrin, S.J.; Carter, O.; Cook, M.J.; Forte, J.D. Effects of a common transcranial direct current stimulation (tDCS) protocol on motor evoked potentials found to be highly variable within individuals over 9 testing sessions. Exp. Brain Res. 2016, 234, 2629–2642. [Google Scholar] [CrossRef]
- De Araújo, A.V.L.; Ribeiro, F.P.G.; Massetti, T.; Potter-Baker, K.A.; Cortes, M.; Plow, E.B.; Da Silva, T.D.; Tonks, J.; Anghinah, R.; Magalhães, F.H.; et al. Effectiveness of anodal transcranial direct current stimulation to improve muscle strength and motor functionality after incomplete spinal cord injury: A systematic review and meta-analysis. Spinal. Cord. 2020, 58, 635–646. [Google Scholar] [CrossRef]
- Baharlouei, H.; Saba, M.A.; Yazdi, M.J.S.; Jaberzadeh, S. The effect of transcranial direct current stimulation on balance in healthy young and older adults: A systematic review of the literature. Neurophysiol. Clin. 2020, 50, 119–131. [Google Scholar] [CrossRef] [PubMed]
- Benussi, A.; Pascual-Leone, A.; Borroni, B. Non-Invasive Cerebellar Stimulation in Neurodegenerative Ataxia: A Literature Review. Int. J. Mol. Sci. 2020, 21, 1948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lloyd, D.M.; Wittkopf, P.G.; Arendsen, L.J.; Jones, A.K.P. Is Transcranial Direct Current Stimulation (tDCS) Effective for the Treatment of Pain in Fibromyalgia? A Systematic Review and Meta-Analysis. J. Pain 2020, 21, 1085–1100. [Google Scholar] [CrossRef] [PubMed]
- Ramger, B.C.; Bader, K.A.; Davies, S.P.; Stewart, D.A.; Ledbetter, L.S.; Simon, C.B.; Feld, J.A. Effects of Non-Invasive Brain Stimulation on Clinical Pain Intensity and Experimental Pain Sensitivity Among Individuals with Central Post-Stroke Pain: A Systematic Review. J. Pain Res. 2019, 12, 3319–3329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiethoff, S.; Hamada, M.; Rothwell, J.C. Variability in Response to Transcranial Direct Current Stimulation of the Motor Cortex. Brain Stimul. 2014, 7, 468–475. [Google Scholar] [CrossRef] [PubMed]
- Smith, M.J.; Keel, J.C.; Greenberg, B.D.; Adams, L.F.; Schmidt, P.J.; Rubinow, D.A.; Wassermann, E.M. Menstrual cycle effects on cortical excitability. Neurology 1999, 53, 2069. [Google Scholar] [CrossRef]
- Pellicciari, M.C.; Brignani, D.; Miniussi, C. Excitability modulation of the motor system induced by transcranial direct current stimulation: A multimodal approach. Neuroimage 2013, 83, 569–580. [Google Scholar] [CrossRef] [Green Version]
- Quiles, V.; Ferrero, L.; Iáñez, E.; Ortiz, M.; Azorín, J.M. Review of tDCS Configurations for Stimulation of the Lower-Limb Area of Motor Cortex and Cerebellum. Brain Sci. 2022, 12, 248. [Google Scholar] [CrossRef]
- Workman, C.D.; Kamholz, J.; Rudroff, T. Increased leg muscle fatigability during 2 mA and 4 mA transcranial direct current stimulation over the left motor cortex. Exp. Brain Res. 2020, 238, 333–343. [Google Scholar] [CrossRef]
- Workman, C.D.; Fietsam, A.C.; Rudroff, T. Transcranial Direct Current Stimulation at 4 mA Induces Greater Leg Muscle Fatigability in Women Compared to Men. Brain Sci. 2020, 10, 244. [Google Scholar] [CrossRef] [Green Version]
- Russell, M.; Goodman, T.; Wang, Q.; Groshong, B.; Lyeth, B.G. Gender Differences in Current Received during Transcranial Electrical Stimulation. Front. Psychiatry 2014, 5, 104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhattacharjee, S.; Kashyap, R.; Goodwill, A.M.; O’Brien, B.A.; Rapp, B.; Oishi, K.; Desmond, J.E.; Chen, S.H.A. Sex difference in tDCS current mediated by changes in cortical anatomy: A study across young, middle and older adults. Brain Stimul. 2022, 15, 125–140. [Google Scholar] [CrossRef] [PubMed]
- Rudroff, T.; Kindred, J.H.; Ketelhut, N.B. Fatigue in Multiple Sclerosis: Misconceptions and Future Research Directions. Front. Neurol. 2016, 7, 122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nitsche, M.A.; Paulus, W. Excitability changes induced in the human motor cortex by weak transcranial direct current stimulation. J. Physiol. 2000, 527, 633–639. [Google Scholar] [CrossRef]
- Krishnan, C.; Ranganathan, R.; Kantak, S.S.; Dhaher, Y.Y.; Rymer, W.Z. Anodal transcranial direct current stimulation alters elbow flexor muscle recruitment strategies. Brain Stimul. 2014, 7, 443–450. [Google Scholar] [CrossRef]
- Alix-Fages, C.; Romero-Arenas, S.; Castro-Alonso, M.; Colomer-Poveda, D.; Río-Rodriguez, D.; Jerez-Martínez, A.; Fernandez-Del-Olmo, M.; Márquez, G. Short-Term Effects of Anodal Transcranial Direct Current Stimulation on Endurance and Maximal Force Production. A Systematic Review and Meta-Analysis. J. Clin. Med. 2019, 8, 536. [Google Scholar] [CrossRef] [Green Version]
- Workman, C.D.; Fietsam, A.C.; Rudroff, T. Different Effects of 2 mA and 4 mA Transcranial Direct Current Stimulation on Muscle Activity and Torque in a Maximal Isokinetic Fatigue Task. Front. Hum. Neurosci. 2020, 14, 240. [Google Scholar] [CrossRef]
- Jang, H.; Lee, J.Y.; Lee, K.I.; Park, K.M. Are there differences in brain morphology according to handedness? Brain Behav. 2017, 7, e00730. [Google Scholar] [CrossRef]
- Pate, R.R. Physical Activity and Public Health. JAMA 1995, 273, 402. [Google Scholar] [CrossRef]
- Dasilva, A.F.; Volz, M.S.; Bikson, M.; Fregni, F. Electrode Positioning and Montage in Transcranial Direct Current Stimulation. J. Vis. Exp. 2011, 23, e2744. [Google Scholar] [CrossRef] [Green Version]
- Allen, A.M.; McRae-Clark, A.L.; Carlson, S.; Saladin, M.E.; Gray, K.M.; Wetherington, C.L.; McKee, S.A.; Allen, S.S. Determining menstrual phase in human biobehavioral research: A review with recommendations. Exp. Clin. Psychopharmacol. 2016, 24, 1. [Google Scholar] [CrossRef] [PubMed]
- Fietsam, A.C.; Deters, J.R.; Workman, C.D.; Rudroff, T. Personal Protective Equipment Alters Leg Muscle Fatigability Independent of Transcranial Direct Current Stimulation: A Comparison with Pre-COVID-19 Pandemic Results. Brain Sci. 2021, 11, 962. [Google Scholar] [CrossRef] [PubMed]
- Workman, C.; Kamholz, J.; Rudroff, T. The Tolerability and Efficacy of 4 mA Transcranial Direct Current Stimulation on Leg Muscle Fatigability. Brain Sci. 2019, 10, 12. [Google Scholar] [CrossRef] [Green Version]
- Foerster, Á.S.; Rezaee, Z.; Paulus, W.; Nitsche, M.A.; Dutta, A. Effects of Cathode Location and the Size of Anode on Anodal Transcranial Direct Current Stimulation Over the Leg Motor Area in Healthy Humans. Front. Neurosci. 2018, 12, 433. [Google Scholar] [CrossRef]
- Jeffery, D.T.; Norton, J.A.; Roy, F.D.; Gorassini, M.A. Effects of transcranial direct current stimulation on the excitability of the leg motor cortex. Exp. Brain Res. 2007, 182, 281–287. [Google Scholar] [CrossRef]
- Santarnecchi, E.; Feurra, M.; Barneschi, F.; Acampa, M.; Bianco, G.; Cioncoloni, D.; Rossi, A.; Rossi, S. Time Course of Corticospinal Excitability and Autonomic Function Interplay during and Following Monopolar tDCS. Front. Psychiatry 2014, 5, 86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Workman, C.D.; Fietsam, A.C.; Rudroff, T. Tolerability and Blinding of Transcranial Direct Current Stimulation in People with Parkinson’s Disease: A Critical Review. Brain Sci. 2020, 10, 467. [Google Scholar] [CrossRef] [PubMed]
- Janse De Jonge, X.; Thompson, B.; Han, A. Methodological Recommendations for Menstrual Cycle Research in Sports and Exercise. Med. Sci. Sports Exerc. 2019, 51, 2610–2617. [Google Scholar] [CrossRef]
- Elliott-Sale, K.J.; Minahan, C.L.; de Jonge, X.; Ackerman, K.E.; Sipila, S.; Constantini, N.W.; Lebrun, C.M.; Hackney, A.C. Methodological Considerations for Studies in Sport and Exercise Science with Women as Participants: A Working Guide for Standards of Practice for Research on Women. Sports Med. 2021, 51, 843–861. [Google Scholar] [CrossRef]
- Thorstensson, A.; Karlsson, J. Fatiguability and fibre composition of human skeletal muscle. Acta Physiol. Scand. 1976, 98, 318–322. [Google Scholar] [CrossRef]
- Lambert, C.P.; Archer, R.L.; Evans, W.J. Muscle strength and fatigue during isokinetic exercise in individuals with multiple sclerosis. Med. Sci. Sports Exerc. 2001, 33, 1613–1619. [Google Scholar] [CrossRef] [PubMed]
- Clos, P.; Mater, A.; Legrand, H.; Poirier, G.; Ballay, Y.; Martin, A.; Lepers, R. Corticospinal Excitability Is Lower During Eccentric Than Concentric Cycling in Men. Front. Physiol. 2022, 13, 854824. [Google Scholar] [CrossRef] [PubMed]
- Tse, D.H.; Kwok, W.Y.; So, B.C. Investigation of Underwater Shoulder Muscle Activity during Manikin-Carrying in Young Elite Lifesaving Athletes. Sensors 2022, 22, 2143. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.S.; Woolley, C.S. Cellular and molecular effects of steroid hormones on CNS excitability. Clevel. Clin. J. Med. 2004, 71, S4. [Google Scholar] [CrossRef] [Green Version]
- Barth, C.; Villringer, A.; Sacher, J. Sex hormones affect neurotransmitters and shape the adult female brain during hormonal transition periods. Front. Neurosci. 2015, 9, 37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, C.H.; Iordanishvili, E.; Shah, N.J.; Binkofski, F. Magnetic resonance spectroscopy with transcranial direct current stimulation to explore the underlying biochemical and physiological mechanism of the human brain: A systematic review. Hum. Brain Mapp. 2021, 42, 2642–2671. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Chung, S.W.; Rogasch, N.C.; Thomson, C.J.; Worsley, R.N.; Kulkarni, J.; Thomson, R.H.; Fitzgerald, P.B.; Segrave, R.A. The influence of endogenous estrogen on transcranial direct current stimulation: A preliminary study. Eur. J. Neurosci. 2018, 48, 2001–2012. [Google Scholar] [CrossRef]
- Chung, S.W.; Thomson, C.J.; Lee, S.; Worsley, R.N.; Rogasch, N.C.; Kulkarni, J.; Thomson, R.H.; Fitzgerald, P.B.; Segrave, R.A. The influence of endogenous estrogen on high-frequency prefrontal transcranial magnetic stimulation. Brain Stimul. 2019, 12, 1271–1279. [Google Scholar] [CrossRef]
- Scalise, A.; Pittaro-Cadore, I.; Russo, V.; Fontana, A.; Simeoni, S.; Gigli, G.L. Absence of delayed facilitation of cortical excitability at early stage of multiple sclerosis: Evidence of altered cortical plasticity? Clin. Neurophysiol. 2013, 124, e143. [Google Scholar] [CrossRef]
- Kishore, A.; Joseph, T.; Velayudhan, B.; Popa, T.; Meunier, S. Early, severe and bilateral loss of LTP and LTD-like plasticity in motor cortex (M1) in de novo Parkinson’s disease. Clin. Neurophysiol. 2012, 123, 822–828. [Google Scholar] [CrossRef]
- Caramia, M.D.; Palmieri, M.G.; Desiato, M.T.; Boffa, L.; Galizia, P.; Rossini, P.M.; Centonze, D.; Bernardi, G. Brain excitability changes in the relapsing and remitting phases of multiple sclerosis: A study with transcranial magnetic stimulation. Clin. Neurophysiol. 2004, 115, 956–965. [Google Scholar] [CrossRef]
- Ayache, S.S.; Créange, A.; Farhat, W.H.; Zouari, H.G.; Lesage, C.; Palm, U.; Abdellaoui, M.; Lefaucheur, J.P. Cortical excitability changes over time in progressive multiple sclerosis. Funct. Neurol. 2015, 30, 257–263. [Google Scholar] [CrossRef]
- Workman, C.D.; Fietsam, A.C.; Uc, E.Y.; Rudroff, T. Cerebellar Transcranial Direct Current Stimulation in People with Parkinson’s Disease: A Pilot Study. Brain Sci. 2020, 10, 96. [Google Scholar] [CrossRef] [Green Version]
- Sarwar, R.; Niclos, B.B.; Rutherford, O.M. Changes in muscle strength, relaxation rate and fatiguability during the human menstrual cycle. J. Physiol. 1996, 493, 267–272. [Google Scholar] [CrossRef] [PubMed]
- Ekenros, L.; Papoutsi, Z.; Friden, C.; Dahlman Wright, K.; Linden Hirschberg, A. Expression of sex steroid hormone receptors in human skeletal muscle during the menstrual cycle. Acta Physiol. 2017, 219, 486–493. [Google Scholar] [CrossRef] [PubMed]
- Hunter, S.K. The Relevance of Sex Differences in Performance Fatigability. Med. Sci. Sports Exerc. 2016, 48, 2247–2256. [Google Scholar] [CrossRef] [Green Version]
- Ansdell, P.; Brownstein, C.G.; Skarabot, J.; Hicks, K.M.; Simoes, D.C.M.; Thomas, K.; Howatson, G.; Hunter, S.K.; Goodall, S. Menstrual cycle-associated modulations in neuromuscular function and fatigability of the knee extensors in eumenorrheic women. J. Appl. Physiol. 2019, 126, 1701–1712. [Google Scholar] [CrossRef]
- Jonge, X.A.K.J.; Boot, C.R.L.; Thom, J.M.; Ruell, P.A.; Thompson, M.W. The influence of menstrual cycle phase on skeletal muscle contractile characteristics in humans. J. Physiol. 2001, 530, 161–166. [Google Scholar] [CrossRef]
- Weidauer, L.; Zwart, M.B.; Clapper, J.; Albert, J.; Vukovich, M.; Specker, B. Neuromuscular performance changes throughout the menstrual cycle in physically active females. J. Musculoskelet Neuronal. Interact. 2020, 20, 314–324. [Google Scholar] [CrossRef] [Green Version]
- Mondini, V.; Mangia, A.L.; Cappello, A. Single-session tDCS over the dominant hemisphere affects contralateral spectral EEG power, but does not enhance neurofeedback-guided event-related desynchronization of the non-dominant hemisphere’s sensorimotor rhythm. PLoS ONE 2018, 13, e0193004. [Google Scholar] [CrossRef] [Green Version]
- Park, C.H.; Chang, W.H.; Park, J.Y.; Shin, Y.I.; Kim, S.T.; Kim, Y.H. Transcranial direct current stimulation increases resting state interhemispheric connectivity. Neurosci. Lett. 2013, 539, 7–10. [Google Scholar] [CrossRef] [PubMed]
- Waters, S.; Wiestler, T.; Diedrichsen, J. Cooperation Not Competition: Bihemispheric tDCS and fMRI Show Role for Ipsilateral Hemisphere in Motor Learning. J. Neurosci. 2017, 37, 7500–7512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Paz, R.H.; Serrano-Muñoz, D.; Pérez-Nombela, S.; Bravo-Esteban, E.; Avendaño-Coy, J.; Gómez-Soriano, J. Combining transcranial direct-current stimulation with gait training in patients with neurological disorders: A systematic review. J. NeuroEng. Rehabil. 2019, 16, 114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaushalya, F.S.; Romero-Arenas, S.; García-Ramos, A.; Colomer-Poveda, D.; Marquez, G. Acute effects of Transcranial Direct Current Stimulation on Cycling and Running Performance. A Systematic Review and Meta-Analysis. Eur. J. Sport Sci. 2020, 22, 1–32. [Google Scholar] [CrossRef]
- Sudbrack-Oliveira, P.; Barbosa, M.Z.; Thome-Souza, S.; Razza, L.B.; Gallucci-Neto, J.; da Costa Lane Valiengo, L.; Brunoni, A.R. Transcranial direct current stimulation (tDCS) in the management of epilepsy: A systematic review. Seizure 2021, 86, 85–95. [Google Scholar] [CrossRef]
- Guerra, A.; López-Alonso, V.; Cheeran, B.; Suppa, A. Variability in non-invasive brain stimulation studies: Reasons and results. Neurosci. Lett. 2020, 719, 133330. [Google Scholar] [CrossRef] [Green Version]
- Russell, M.J.; Goodman, T.A.; Visse, J.M.; Beckett, L.; Saito, N.; Lyeth, B.G.; Recanzone, G.H. Sex and Electrode Configuration in Transcranial Electrical Stimulation. Front. Psychiatry 2017, 8, 147. [Google Scholar] [CrossRef] [Green Version]
- Parks, R.B.; Hetzel, S.J.; Brooks, M.A. Iron Deficiency and Anemia among Collegiate Athletes: A Retrospective Chart Review. Med. Sci. Sports Exerc. 2017, 49, 1711–1715. [Google Scholar] [CrossRef]
- Benson, C.S.; Shah, A.; Stanworth, S.J.; Frise, C.J.; Spiby, H.; Lax, S.J.; Murray, J.; Klein, A.A. The effect of iron deficiency and anaemia on women’s health. Anaesthesia 2021, 76, 84–95. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Average Estrogen Level (pg/mL) | Itching | Burning | Tingling | Prickling | |
|---|---|---|---|---|---|
| Active (High) | 157.7 ± 101.5 | (3) 4.3 ± 1.5 | (3) 5.3 ± 2.1 | (3) 3.3 ± 2.1 | (1) 5 |
| Active (Low) | 31.9 ± 10.9 | X | (3) 5.3 ± 2.3 | (3) 4.7 ± 1.2 | (2) 5.5 ± 2.1 |
| Sham (High) | 137.2 ± 114.0 | (3) 4.5 ± 2.1 | (4) 5.3 ± 2.1 | (3) 4.3 ± 1.5 | (4) 4.0 ± 2.6 |
| Sham (Low) | 42.9 ± 25.1 | (5) 3.4 ± 1.7 | (4) 4.75 ± 1.9 | (2) 3.4 ± 1.7 | (2) 4.0 ± 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Deters, J.R.; Fietsam, A.C.; Workman, C.D.; Rudroff, T. High Estrogen Levels Cause Greater Leg Muscle Fatigability in Eumenorrheic Young Women after 4 mA Transcranial Direct Current Stimulation. Brain Sci. 2022, 12, 506. https://doi.org/10.3390/brainsci12040506
Deters JR, Fietsam AC, Workman CD, Rudroff T. High Estrogen Levels Cause Greater Leg Muscle Fatigability in Eumenorrheic Young Women after 4 mA Transcranial Direct Current Stimulation. Brain Sciences. 2022; 12(4):506. https://doi.org/10.3390/brainsci12040506
Chicago/Turabian StyleDeters, Justin R., Alexandra C. Fietsam, Craig D. Workman, and Thorsten Rudroff. 2022. "High Estrogen Levels Cause Greater Leg Muscle Fatigability in Eumenorrheic Young Women after 4 mA Transcranial Direct Current Stimulation" Brain Sciences 12, no. 4: 506. https://doi.org/10.3390/brainsci12040506