
Integrative Hedonic and Homeostatic Food Intake Regulation by the Central Nervous System: Insights from Neuroimaging


Abstract
:1. Introduction
2. Methods
3. Homeostatic Food Intake Regulation
3.1. Peripheral Metabolic Signals
3.1.1. Leptin
3.1.2. Ghrelin
3.1.3. Cholecystokinin
3.1.4. Glucagon-like Peptide-1
3.1.5. Peptide Y (PYY)
3.2. Visceroceptive Feedback
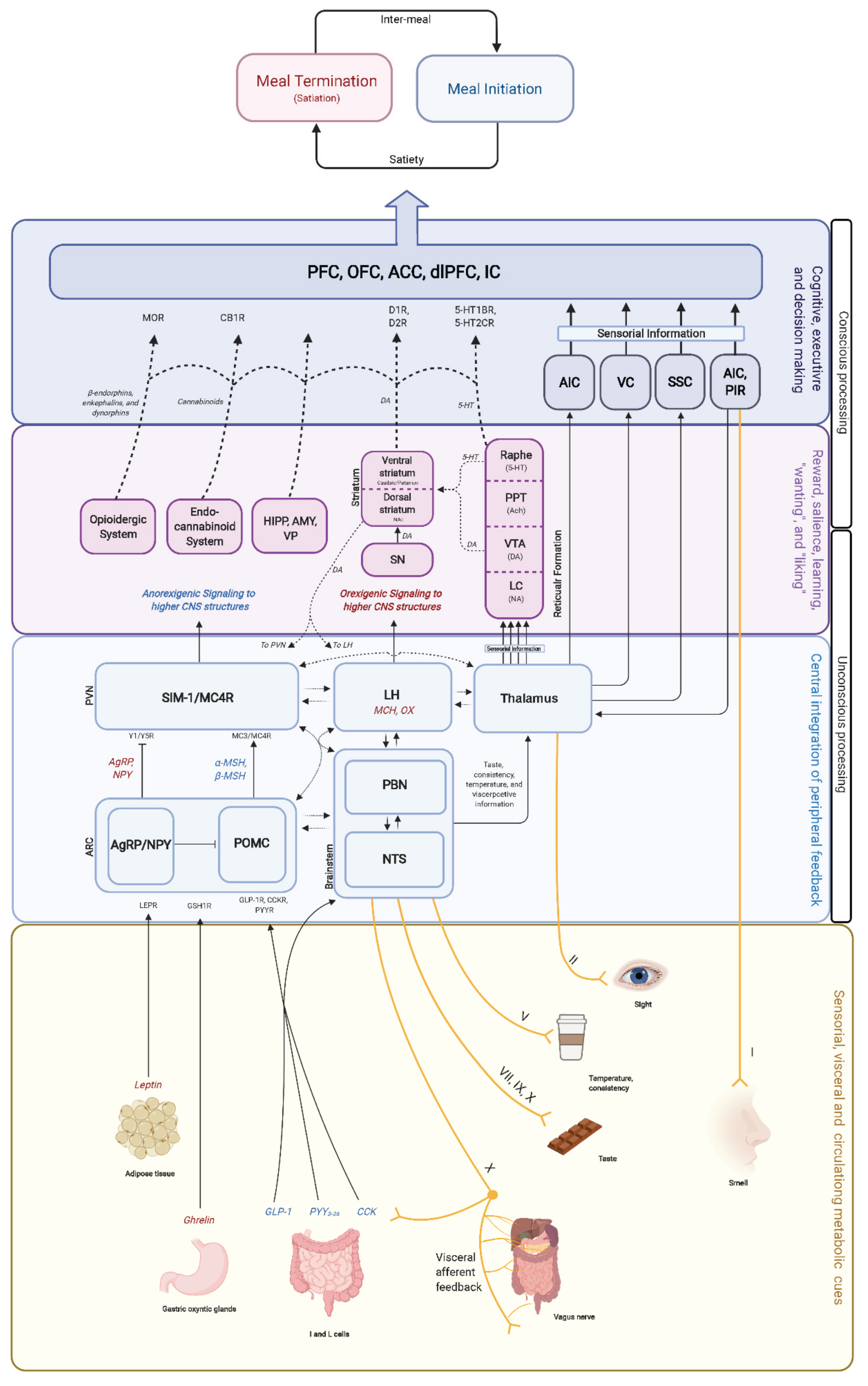
3.3. Central Nervous System Regulation
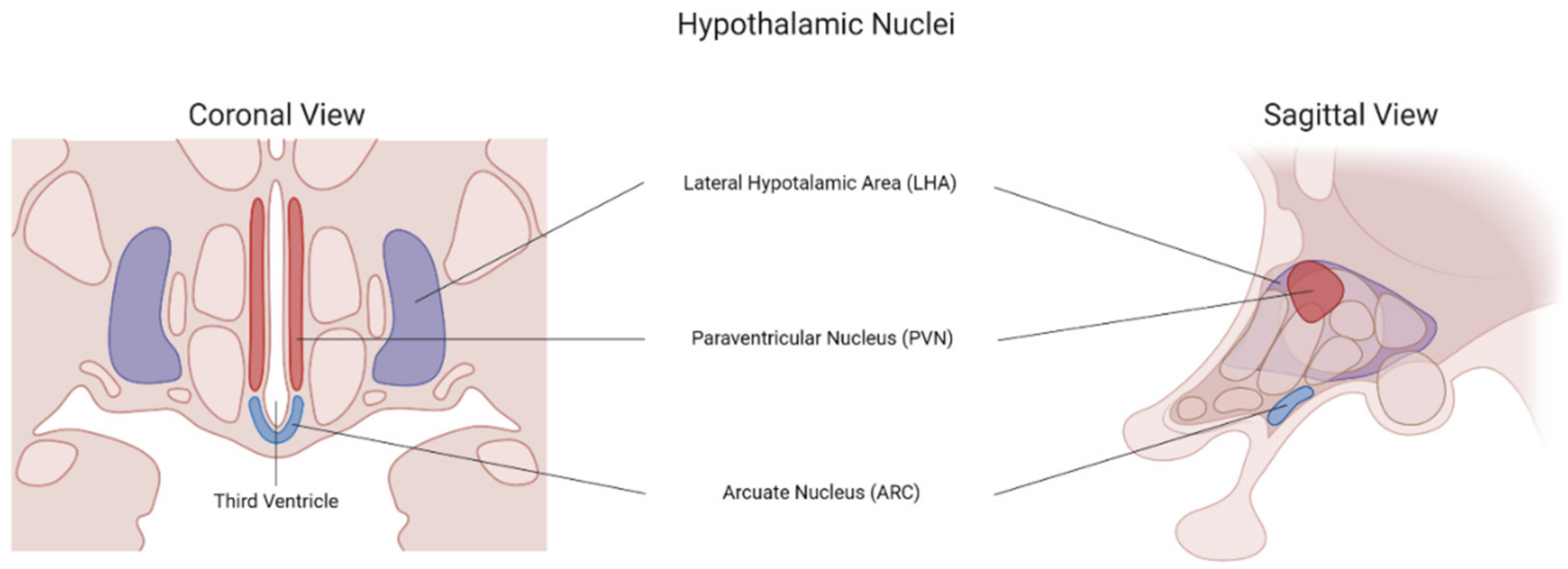
3.3.1. The Hypothalamic Leptin–Melanocortin Pathway
3.3.2. The Lateral Hypothalamic Hypocretin Pathway
3.3.3. Sensitive and Sensorial Information
3.4. Stages of Food Intake: Hunger, Satiety, and Satiation
4. Hedonic Food Intake Regulation
4.1. Neurotransmitters and Neuropeptides
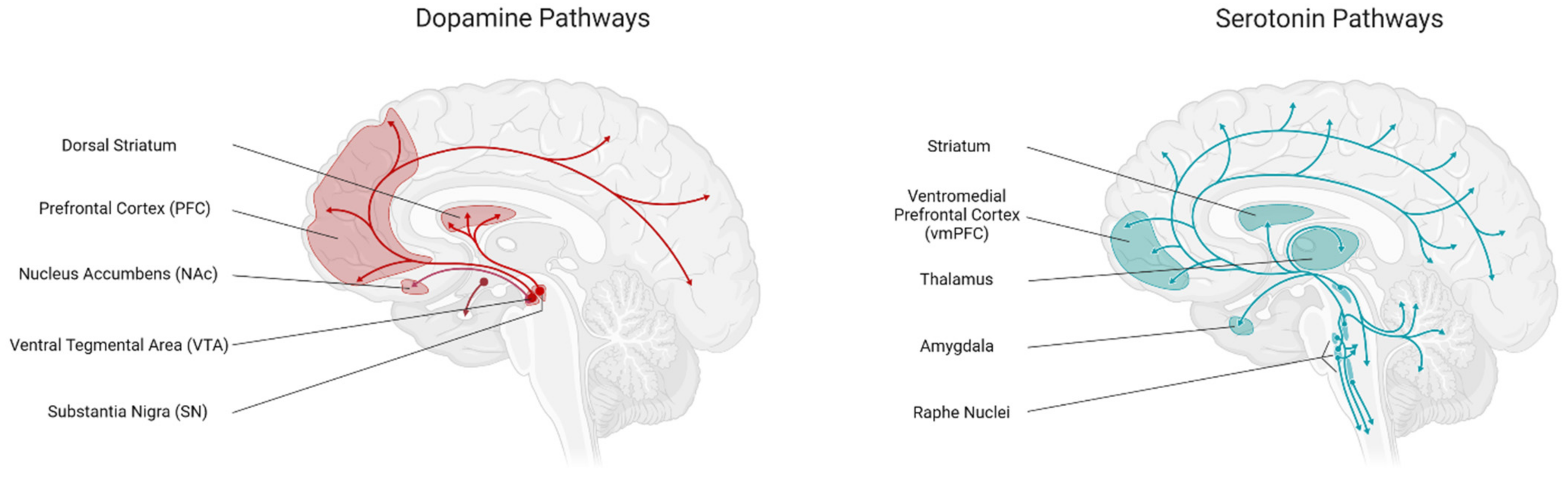
4.1.1. Dopamine
4.1.2. Serotonin (5-HT)
4.1.3. Endocannabinoid and Opioid Systems
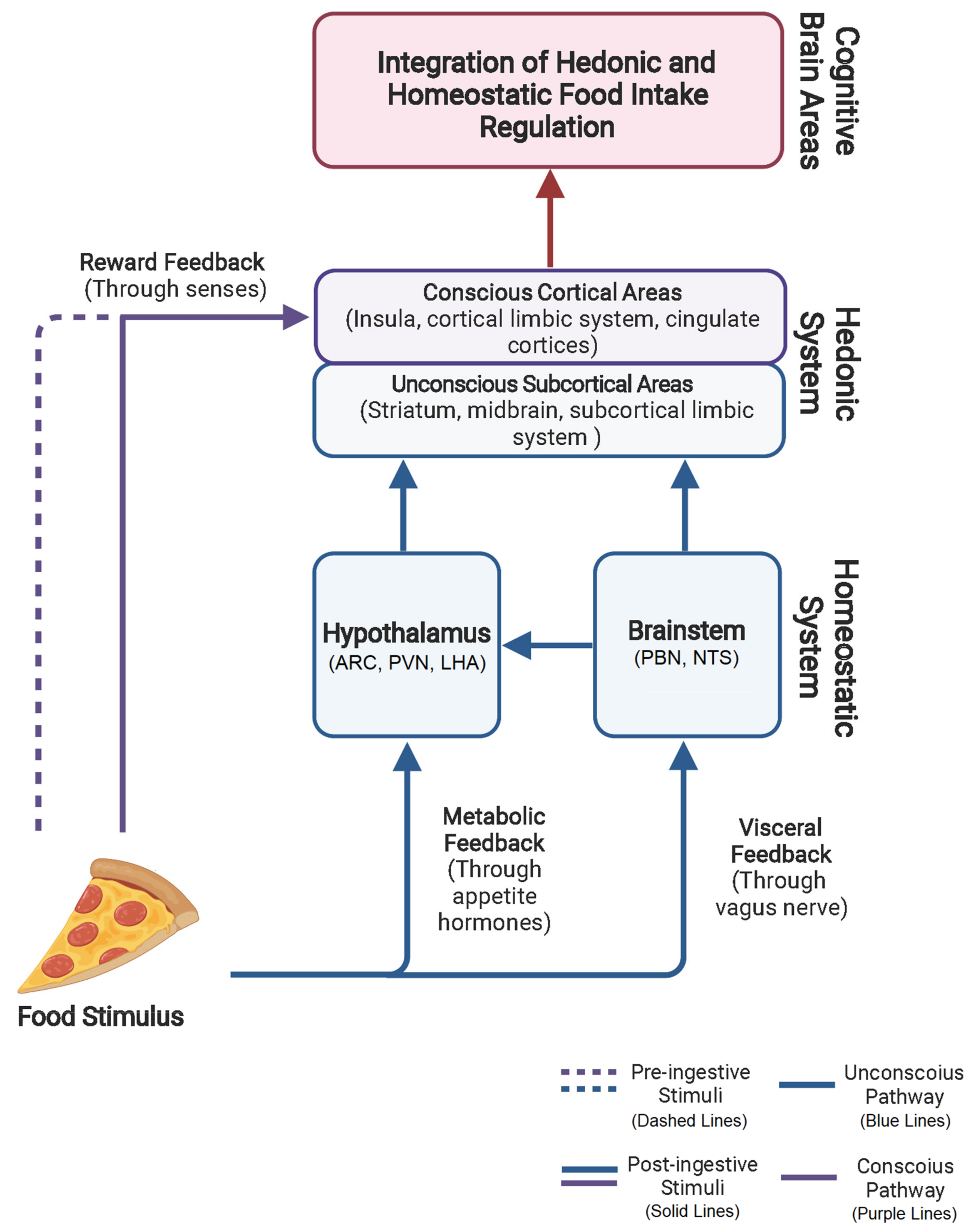
4.2. Homeostatic and Hedonic System Overlap and Interaction
5. Future Directives in Food Intake Neuroimaging
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Blundell, J.; De Graaf, C.; Hulshof, T.; Jebb, S.; Livingstone, B.; Lluch, A.; Mela, D.; Salah, S.; Schuring, E.; Van Der Knaap, H.; et al. Appetite control: Methodological aspects of the evaluation of foods. Obes. Rev. 2010, 11, 251–270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cifuentes, L.; Acosta, A. Homeostatic regulation of food intake. Clin. Res. Hepatol. Gastroenterol. 2021, 46, 101794. [Google Scholar] [CrossRef] [PubMed]
- Sclafani, A. Gut–brain nutrient signaling. Appetition vs. satiation. Appetite 2013, 71, 454–458. [Google Scholar]
- Simpson, K.A.; Martin, N.M.; Bloom, S.R. Hypothalamic regulation of food intake and clinical therapeutic applications. Arq. Bras. Endocrinol. Metabol. 2009, 53, 120–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Avena, N. Hedonic Eating: How the Pleasure of Food Affects Our Brains and Behavior; Oxford University Press: Oxford, UK, 2015. [Google Scholar]
- De Araujo, I.E.; Schatzker, M.; Small, D.M. Rethinking Food Reward. Annu. Rev. Psychol. 2020, 71, 139–164. [Google Scholar] [CrossRef]
- Laughlin, M.; Cooke, B.; Boutelle, K.; Savage, C.R.; Kravitz, A.; Small, D.; Arvanitakis, Z.; Stoeckel, A.M.L.E. Neuroimaging and modulation in obesity and diabetes research: 10th anniversary meeting. Int. J. Obes. 2021, 110, 43–52. [Google Scholar]
- Krashes, M.J.; Kravitz, A.V. Optogenetic and chemogenetic insights into the food addiction hypothesis. Front. Behav. Neurosci. 2014, 8, 57. [Google Scholar] [CrossRef] [Green Version]
- Whissell, P.D.; Tohyama, S.; Martin, L.J. The Use of DREADDs to Deconstruct Behavior. Front. Genet. 2016, 7, 70. [Google Scholar] [CrossRef] [Green Version]
- Urstadt, K.R.; Berridge, K.C. Optogenetic mapping of feeding and self-stimulation within the lateral hypothalamus of the rat. PLoS ONE 2020, 15, e0224301. [Google Scholar] [CrossRef] [Green Version]
- Anand, B.K.; Brobeck, J.R. Localization of a “Feeding Center” in the Hypothalamus of the Rat. Exp. Biol. Med. 1951, 77, 323–325. [Google Scholar] [CrossRef]
- Han, P. Advances in research on brain processing of food odors using different neuroimaging techniques. Curr. Opin. Food Sci. 2021, 42, 134–139. [Google Scholar] [CrossRef]
- Zanchi, D.; Depoorter, A.; Egloff, L.; Haller, S.; Mählmann, L.; Lang, U.E.; Drewe, J.; Beglinger, C.; Schmidt, A.; Borgwardt, S. The impact of gut hormones on the neural circuit of appetite and satiety: A systematic review. Neurosci. Biobehav. Rev. 2017, 80, 457–475. [Google Scholar] [CrossRef] [PubMed]
- Devoto, F.; Zapparoli, L.; Bonandrini, R.; Berlingeri, M.; Ferrulli, A.; Luzi, L.; Banfi, G.; Paulesu, E. Hungry brains: A meta-analytical review of brain activation imaging studies on food perception and appetite in obese individuals. Neurosci. Biobehav. Rev. 2018, 94, 271–285. [Google Scholar] [CrossRef] [PubMed]
- Cambridge University Press. Principles of Functional Magnetic Resonance Imaging. In Introduction to Functional Magnetic Resonance Imaging: Principles and Techniques; Buxton, R.B., Ed.; Cambridge University Press: Cambridge, UK, 2009; pp. 277–424. [Google Scholar]
- Detre, J.A.; Wang, J. Technical aspects and utility of fMRI using BOLD and ASL. Clin. Neurophysiol. 2002, 113, 621–634. [Google Scholar] [CrossRef] [PubMed]
- Cornier, M.-A.; Melanson, E.L.; Salzberg, A.K.; Bechtell, J.L.; Tregellas, J.R. The effects of exercise on the neuronal response to food cues. Physiol. Behav. 2012, 105, 1028–1034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masterson, T.D.; Kirwan, C.B.; Davidson, L.E.; Larson, M.; Keller, K.L.; Fearnbach, S.N.; Evans, A.; LeCheminant, J.D. Brain reactivity to visual food stimuli after moderate-intensity exercise in children. Brain Imaging Behav. 2018, 12, 1032–1041. [Google Scholar] [CrossRef] [PubMed]
- Evero, N.; Hackett, L.C.; Clark, R.D.; Phelan, S.; Hagobian, T.A. Aerobic exercise reduces neuronal responses in food reward brain regions. J. Appl. Physiol. 2012, 112, 1612–1619. [Google Scholar] [CrossRef] [Green Version]
- Crabtree, D.; Chambers, E.S.; Hardwick, R.M.; Blannin, A.K. The effects of high-intensity exercise on neural responses to images of food. Am. J. Clin. Nutr. 2013, 99, 258–267. [Google Scholar] [CrossRef] [Green Version]
- Fröhlich, A. Ein Fall von Tumor der Hypophysis Cerebri Ohne Akromegalie; Wiener Klinische Rundschau: Wien, Austria, 1901; Volume 15, pp. 833–836, 906–908. [Google Scholar]
- Bruch, H. The Fröhlich Syndrome: Report of The Original Case. Am. J. Dis. Child. 1993, 58, 1282–1289. [Google Scholar] [CrossRef]
- Hervey, G.R. The effects of lesions in the hypothalamus in parabiotic rats. J. Physiol. 1959, 145, 336–352. [Google Scholar] [CrossRef]
- Zhang, Y.; Proenca, R.; Maffei, M.; Barone, M.; Leopold, L.; Friedman, J.M. Positional cloning of the mouse obese gene and its human homologue. Nature 1994, 372, 425–432. [Google Scholar] [CrossRef] [PubMed]
- Friedman, J. 20 YEARS OF LEPTIN: Leptin at 20: An overview. J. Endocrinol. 2014, 223, T1–T8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeo, G.S.H.; Farooqi, S.; Aminian, S.; Halsall, D.J.; Stanhope, R.G.; O’Rahilly, S. A frameshift mutation in MC4R associated with dominantly inherited human obesity. Nat. Genet. 1998, 20, 111–112. [Google Scholar] [CrossRef] [PubMed]
- Farooqi, I.S.; Jebb, S.A.; Langmack, G.; Lawrence, E.; Cheetham, C.H.; Prentice, A.M.; Hughes, I.A.; McCamish, M.A.; O’Rahilly, S. Effects of recombinant leptin therapy in a child with congenital leptin deficiency. N. Engl. J. Med. 1999, 341, 879–884. [Google Scholar]
- Farooqi, I.S.; Keogh, J.M.; Yeo, G.S.; Lank, E.J.; Cheetham, T.; O’Rahilly, S. Clinical spectrum of obesity and mutations in the melanocortin 4 receptor gene. N. Engl. J. Med. 2003, 348, 1085–1095. [Google Scholar]
- Locke, A.E.; Kahali, B.; Berndt, S.I.; Justice, A.E.; Pers, T.H.; Day, F.R.; Powell, C.; Vedantam, S.; Buchkovich, M.L.; Yang, J.; et al. Genetic studies of body mass index yield new insights for obesity biology. Nature 2015, 518, 197–206. [Google Scholar] [CrossRef] [Green Version]
- Camilleri, M. Peripheral Mechanisms in Appetite Regulation. Gastroenterology 2015, 148, 1219–1233. [Google Scholar] [CrossRef] [Green Version]
- Saper, C.B.; Chou, T.C.; Elmquist, J.K. The Need to Feed: Homeostatic and Hedonic Control of Eating. Neuron 2002, 36, 199–211. [Google Scholar] [CrossRef] [Green Version]
- Clarke, I. Hypothalamus as an Endocrine Organ. Compr. Physiol. 2014, 5, 217–253. [Google Scholar] [CrossRef]
- Rodríguez, E.M.; Blázquez, J.L.; Guerra, M. The design of barriers in the hypothalamus allows the median eminence and the arcuate nucleus to enjoy private milieus: The former opens to the portal blood and the latter to the cerebrospinal fluid. Peptides 2010, 31, 757–776. [Google Scholar]
- Peruzzo, B.; Pastor, F.E.; Blázquez, J.L.; Schöbitz, K.; Peláez, B.; Amat, P.; Rodríguez, E.M. A second look at the barriers of the medial basal hypothalamus. Exp. Brain Res. 2000, 132, 10–26. [Google Scholar] [CrossRef] [PubMed]
- Steinert, R.E.; Feinle-Bisset, C.; Asarian, L.; Horowitz, M.; Beglinger, C.; Geary, N. Ghrelin, CCK, GLP-1, and PYY(3–36): Secretory Controls and Physiological Roles in Eating and Glycemia in Health, Obesity, and After RYGB. Physiol. Rev. 2017, 97, 411–463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abizaid, A.; Gao, Q.; Horvath, T.L. Thoughts for Food: Brain Mechanisms and Peripheral Energy Balance. Neuron 2006, 51, 691–702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwartz, G.J. Brainstem Integrative Function in the Central Nervous System Control of Food Intake. In Frontiers in Eating and Weight Regulation; S. Karger AG: Basel, Switzerland, 2010; Volume 63, pp. 141–151. [Google Scholar]
- Atkinson, T.J. Central and peripheral neuroendocrine peptides and signalling in appetite regulation: Considerations for obesity pharmacotherapy. Obes. Rev. 2008, 9, 108–120. [Google Scholar] [CrossRef] [PubMed]
- Blundell, J.E.; Halford, J.C.G. Regulation of nutrient supply: The brain and appetite control. Proc. Nutr. Soc. 1994, 53, 407–418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berthoud, H.-R. The vagus nerve, food intake and obesity. Regul. Pept. 2008, 149, 15–25. [Google Scholar] [CrossRef] [Green Version]
- Hussain, Z.; Khan, J.A. Food intake regulation by leptin: Mechanisms mediating gluconeogenesis and energy expenditure. Asian Pac. J. Trop. Med. 2017, 10, 940–944. [Google Scholar] [CrossRef]
- Flier, S.J.; Maratos-Flier, E. Leptin’s physiologic role: Does the emperor of energy balance have no clothes? Cell Metab. 2017, 26, 24–26. [Google Scholar]
- Collet, T.H.; Sonoyama, T.; Henning, E.; Keogh, J.M.; Ingram, B.; Kelway, S.; Guo, L.; Farooqi, I.S. A Metabolomic Signature of Acute Caloric Restriction. 2017. Available online: https://academic.oup.com/jcem/article/102/12/4486/4209888?login=true (accessed on 4 February 2022).
- Balland, E.; Dam, J.; Langlet, F.; Caron, E.; Steculorum, S.; Messina, A.; Rasika, S.; Falluel-Morel, A.; Anouar, Y.; Dehouck, B.; et al. Hypothalamic Tanycytes Are an ERK-Gated Conduit for Leptin into the Brain. Cell Metab. 2014, 19, 293–301. [Google Scholar] [CrossRef] [Green Version]
- Elias, C.F.; Aschkenasi, C.; Lee, C.; Kelly, J.; Ahima, R.S.; Bjorbæk, C.; Flier, J.S.; Saper, C.B.; Elmquist, J.K. Leptin Differentially Regulates NPY and POMC Neurons Projecting to the Lateral Hypothalamic Area. Neuron 1999, 23, 775–786. [Google Scholar] [CrossRef] [Green Version]
- Cowley, M.A.; Smart, J.L.; Rubinstein, M.; Cerdán, M.G.; Diano, S.; Horvath, T.L.; Cone, R.D.; Low, M.J. Leptin activates anorexigenic POMC neurons through a neural network in the arcuate nucleus. Nature 2001, 411, 480–484. [Google Scholar] [CrossRef] [PubMed]
- Spanswick, D.; Smith, M.; Groppi, V.E.; Logan, S.D.; Ashford, M. Leptin inhibits hypothalamic neurons by activation of ATP-sensitive potassium channels. Nature 1997, 390, 521–525. [Google Scholar] [CrossRef] [PubMed]
- Farooqi, I.S.; Bullmore, E.; Keogh, J.; Gillard, J.; O’Rahilly, S.; Fletcher, P.C. Leptin Regulates Striatal Regions and Human Eating Behavior. Science 2007, 317, 1355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baicy, K.; London, E.D.; Monterosso, J.; Wong, M.-L.; Delibasi, T.; Sharma, A.; Licinio, J. Leptin replacement alters brain response to food cues in genetically leptin-deficient adults. Proc. Natl. Acad. Sci. USA 2007, 104, 18276–18279. [Google Scholar] [CrossRef] [Green Version]
- Berman, S.M.; Paz-Filho, G.; Wong, M.-L.; Kohno, M.; Licinio, J.; London, E.D. Effects of Leptin Deficiency and Replacement on Cerebellar Response to Food-Related Cues. Cerebellum 2012, 12, 59–67. [Google Scholar] [CrossRef] [Green Version]
- Mendoza, J.; Pévet, P.; Felder-Schmittbuhl, M.-P.; Bailly, Y.; Challet, E. The Cerebellum Harbors a Circadian Oscillator Involved in Food Anticipation. J. Neurosci. 2010, 30, 1894–1904. [Google Scholar] [CrossRef]
- Vollmert, C.; Grosshans, M.; Vollstädt-Klein, S.; Tost, H.; Leber, S.; Bach, P.; Bühler, M.; Von Der Goltz, C.; Mutschler, J.; Loeber, S.; et al. Association of Leptin With Food Cue–Induced Activation in Human Reward Pathways. Arch. Gen. Psychiatry 2012, 69, 529–537. [Google Scholar] [CrossRef] [Green Version]
- Rosenbaum, M.; Sy, M.; Pavlovich, K.; Leibel, R.L.; Hirsch, J. Leptin reverses weight loss–induced changes in regional neural activity responses to visual food stimuli. J. Clin. Investig. 2008, 118, 2583–2591. [Google Scholar] [CrossRef] [Green Version]
- Date, Y.; Kojima, M.; Hosoda, H.; Sawaguchi, A.; Mondal, M.S.; Suganuma, T.; Matsukura, S.; Kangawa, K.; Nakazato, M. Ghrelin, a novel growth hormone-releasing acylated peptide, is synthesized in a distinct endocrine cell type in the gastrointestinal tracts of rats and humans. Endocrinology 2000, 141, 4255–4261. [Google Scholar]
- Muller, T.D.; Nogueiras, R.; Andermann, M.L.; Andrews, Z.B.; Anker, S.D.; Argente, J.; Batterham, R.L.; Benoit, S.C.; Bowers, C.Y.; Broglio, F.; et al. Ghrelin. Mol. Metab. 2015, 4, 437–460. [Google Scholar]
- Akamizu, T.; Takaya, K.; Irako, T.; Hosoda, H.; Teramukai, S.; Matsuyama, A.; Tada, H.; Miura, K.; Shimizu, A.; Fukushima, M.; et al. Pharmacokinetics, safety, and endocrine and appetite effects of ghrelin administration in young healthy subjects. Eur. J. Endocrinol. 2004, 150, 447–455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willesen, M.G.; Kristensen, P.; Rømer, J. Co-Localization of Growth Hormone Secretagogue Receptor and NPY mRNA in the Arcuate Nucleus of the Rat. Neuroendocrinology 1999, 70, 306–316. [Google Scholar] [CrossRef] [PubMed]
- Skibicka, K.P.; Shirazi, R.H.; Rabasa-Papio, C.; Alvarez-Crespo, M.; Neuber, C.; Vogel, H.; Dickson, S.L. Divergent circuitry underlying food reward and intake effects of ghrelin: Dopaminergic VTA-accumbens projection mediates ghrelin’s effect on food reward but not food intake. Neuropharmacology 2013, 73, 274–283. [Google Scholar]
- Albarran-Zeckler, R.G.; Sun, Y.; Smith, R.G. Physiological roles revealed by ghrelin and ghrelin receptor deficient mice. Peptides 2011, 32, 2229–2235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chin, S.-H.; Kahathuduwa, C.N.; Stearns, M.B.; Davis, T.; Binks, M. Is hunger important to model in fMRI visual food-cue reactivity paradigms in adults with obesity and how should this be done? Appetite 2018, 120, 388–397. [Google Scholar] [CrossRef]
- Prechtl de Hernandez, C. Modulation of Food Reward Systems by Fasting and Ghrelin: Human Functional MRI Studies; Imperial College London: London, UK, 2011. [Google Scholar]
- Malik, S.; McGlone, F.; Bedrossian, D.; Dagher, A. Ghrelin Modulates Brain Activity in Areas that Control Appetitive Behavior. Cell Metab. 2008, 7, 400–409. [Google Scholar] [CrossRef] [Green Version]
- Goldstone, A.P.; Prechtl, C.G.; Scholtz, S.; Miras, A.; Chhina, N.; Durighel, G.; Deliran, S.S.; Beckmann, C.F.; Ghatei, M.A.; Ashby, D.R.; et al. Ghrelin mimics fasting to enhance human hedonic, orbitofrontal cortex, and hippocampal responses to food. Am. J. Clin. Nutr. 2014, 99, 1319–1330. [Google Scholar] [CrossRef]
- Jones, R.B.; McKie, S.; Astbury, N.; Little, T.; Tivey, S.; Lassman, D.J.; McLaughlin, J.; Luckman, S.; Williams, S.; Dockray, G.J.; et al. Functional neuroimaging demonstrates that ghrelin inhibits the central nervous system response to ingested lipid. Gut 2012, 61, 1543–1551. [Google Scholar] [CrossRef]
- Karra, E.; O’Daly, O.G.; Choudhury, A.I.; Yousseif, A.; Millership, S.; Neary, M.T.; Scott, W.R.; Chandarana, K.; Manning, S.; Hess, M.E.; et al. A link between FTO, ghrelin, and impaired brain food-cue responsivity. J. Clin. Investig. 2013, 123, 3539–3551. [Google Scholar] [CrossRef] [Green Version]
- Little, T.; Horowitz, M.; Feinle-Bisset, C. Role of cholecystokinin in appetite control and body weight regulation. Obes. Rev. 2005, 6, 297–306. [Google Scholar] [CrossRef]
- Eldeghaidy, S.; Marciani, L.; Hort, J.; Hollowood, T.; Singh, G.; Bush, D.; Foster, T.; Taylor, A.J.; Busch, J.; Spiller, R.C.; et al. Prior Consumption of a Fat Meal in Healthy Adults Modulates the Brain’s Response to Fat. J. Nutr. 2016, 146, 2187–2198. [Google Scholar] [PubMed] [Green Version]
- Lassman, D.J.; McKie, S.; Gregory, L.J.; Lal, S.; D’Amato, M.; Steele, I.; Varro, A.; Dockray, G.J.; Williams, S.; Thompson, D.G. Defining the Role of Cholecystokinin in the Lipid-Induced Human Brain Activation Matrix. Gastroenterology 2010, 138, 1514–1524. [Google Scholar] [CrossRef] [PubMed]
- Psichas, A.; Reimann, F.; Gribble, F.M. Gut chemosensing mechanisms. J. Clin. Investig. 2015, 125, 908–917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campbell, J.E.; Drucker, D.J. Pharmacology, Physiology, and Mechanisms of Incretin Hormone Action. Cell Metab. 2013, 17, 819–837. [Google Scholar] [CrossRef] [Green Version]
- Parkinson, J.R.; Chaudhri, O.B.; Kuo, Y.-T.; Field, B.C.; Herlihy, A.H.; Dhillo, W.S.; Ghatei, M.A.; Bloom, S.R.; Bell, J.D. Differential patterns of neuronal activation in the brainstem and hypothalamus following peripheral injection of GLP-1, oxyntomodulin and lithium chloride in mice detected by manganese-enhanced magnetic resonance imaging (MEMRI). NeuroImage 2009, 44, 1022–1031. [Google Scholar] [CrossRef]
- Holst, J.J. The Physiology of Glucagon-like Peptide. Physiol. Rev. 2007, 87, 1409–1439. [Google Scholar] [CrossRef]
- Van Bloemendaal, L.; IJzerman, R.G.; Ten Kulve, J.S.; Barkhof, F.; Konrad, R.J.; Drent, M.L.; Veltman, D.J.; Diamant, M. GLP-1 Receptor Activation Modulates Appetite- and Reward-Related Brain Areas in Humans. Diabetes 2014, 63, 4186. [Google Scholar]
- Seeley, R.; Blake, K.; Rushing, P.A.; Benoit, S.; Eng, J.; Woods, S.C.; D’Alessio, D. The Role of CNS Glucagon-Like Peptide-1 (7-36) Amide Receptors in Mediating the Visceral Illness Effects of Lithium Chloride. J. Neurosci. 2000, 20, 1616–1621. [Google Scholar] [CrossRef]
- Yamamoto, H.; Lee, C.E.; Marcus, J.N.; Williams, T.D.; Overton, J.M.; Lopez, M.E.; Hollenberg, A.N.; Baggio, L.; Saper, C.B.; Drucker, D.J.; et al. Glucagon-like peptide-1 receptor stimulation increases blood pressure and heart rate and activates autonomic regulatory neurons. J. Clin. Investig. 2002, 110, 43–52. [Google Scholar]
- Thiele, T.E.; Van Dijk, G.; Campfield, L.A.; Smith, F.J.; Burn, P.; Woods, S.C.; Bernstein, I.L.; Seeley, R. Central infusion of GLP-1, but not leptin, produces conditioned taste aversions in rats. Am. J. Physiol. Integr. Comp. Physiol. 1997, 272, R726–R730. [Google Scholar] [CrossRef] [Green Version]
- De Silva, A.; Salem, V.; Long, C.J.; Makwana, A.; Newbould, R.D.; Rabiner, E.A.; Ghatei, M.A.; Bloom, S.R.; Matthews, P.M.; Beaver, J.D.; et al. The Gut Hormones PYY3-36 and GLP-17-36 amide Reduce Food Intake and Modulate Brain Activity in Appetite Centers in Humans. Cell Metab. 2011, 14, 700–706. [Google Scholar] [CrossRef] [Green Version]
- Schlögl, H.; Kabisch, S.; Horstmann, A.; Lohmann, G.; Müller, K.; Lepsien, J.; Busse-Voigt, F.; Kratzsch, J.; Pleger, B.; Villringer, A.; et al. Exenatide-Induced Reduction in Energy Intake Is Associated With Increase in Hypothalamic Connectivity. Diabetes Care 2013, 36, 1933–1940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kulve, J.S.T.; Veltman, D.J.; Van Bloemendaal, L.; Barkhof, F.; Deacon, C.; Holst, J.J.; Konrad, R.J.; Sloan, J.H.; Drent, M.L.; Diamant, M.; et al. Endogenous GLP-1 mediates postprandial reductions in activation in central reward and satiety areas in patients with type 2 diabetes. Diabetologia 2015, 58, 2688–2698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farr, O.M.; Sofopoulos, M.; Tsoukas, M.; Dincer, F.; Thakkar, B.; Sahin-Efe, A.; Filippaios, A.; Bowers, J.; Srnka, A.; Gavrieli, A.; et al. GLP-1 receptors exist in the parietal cortex, hypothalamus and medulla of human brains and the GLP-1 analogue liraglutide alters brain activity related to highly desirable food cues in individuals with diabetes: A crossover, randomised, placebo-controlled trial. Diabetologia 2016, 59, 954–965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larraufie, P.; Martin-Gallausiaux, C.; Lapaque, N.; Dore, J.; Gribble, F.M.; Reimann, F.; Blottiere, H.M. SCFAs strongly stimulate PYY production in human enteroendocrine cells. Sci. Rep. 2018, 8, 74. [Google Scholar] [CrossRef]
- Batterham, R.L.; Cowley, M.A.; Small, C.J.; Herzog, H.; Cohen, M.A.; Dakin, C.L.; Wren, A.M.; Brynes, A.E.; Low, M.J.; Ghatei, M.A.; et al. Gut hormone PYY3-36 physiologically inhibits food intake. Nature 2002, 418, 650–654. [Google Scholar] [CrossRef]
- Hanusch-Enserer, U.; Roden, M. News in gut-brain communication: A role of peptide YY (PYY) in human obesity and following bariatric surgery? Eur. J. Clin. Investig. 2005, 35, 425–430. [Google Scholar] [CrossRef]
- Grandt, D.; Schimiczek, M.; Beglinger, C.; Layer, P.; Goebell, H.; Eysselein, V.E.; Reeve, J.R., Jr. Two molecular forms of Peptide YY (PYY) are abundant in human blood: Characterization of a radioimmunoassay recognizing PYY 1–36 and PYY 3–36. Regul. Pept. 1994, 51, 151–159. [Google Scholar] [CrossRef]
- Baraboi, E.-D.; Michel, C.; Smith, P.; Thibaudeau, K.; Ferguson, A.V.; Richard, D. Effects of albumin-conjugated PYY on food intake: The respective roles of the circumventricular organs and vagus nerve. Eur. J. Neurosci. 2010, 32, 826–839. [Google Scholar] [CrossRef]
- Batterham, R.L.; Ffytche, D.H.; Rosenthal, J.M.; Zelaya, F.O.; Barker, G.J.; Withers, D.J.; Williams, S.C.R. PYY modulation of cortical and hypothalamic brain areas predicts feeding behaviour in humans. Nature 2007, 450, 106–109. [Google Scholar] [CrossRef]
- Perakakis, N.; Farr, O.M.; Mantzoros, C.S. Fasting oxyntomodulin, glicentin, and gastric inhibitory polypeptide levels are associated with activation of reward- and attention-related brain centres in response to visual food cues in adults with obesity: A cross-sectional functional MRI study. Diabetes Obes. Metab. 2021, 23, 1202–1207. [Google Scholar] [CrossRef] [PubMed]
- Chambers, A.P.; Sandoval, D.A.; Seeley, R.J. Integration of Satiety Signals by the Central Nervous System. Curr. Biol. 2013, 23, R379–R388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cechetto, D.F.; Saper, C.B. Evidence for a viscerotopic sensory representation in the cortex and thalamus in the rat. J. Comp. Neurol. 1987, 262, 27–45. [Google Scholar] [CrossRef]
- Ly, H.G.; Dupont, P.; Van Laere, K.; Depoortere, I.; Tack, J.; Van Oudenhove, L. Differential brain responses to gradual intragastric nutrient infusion and gastric balloon distension: A role for gut peptides? NeuroImage 2017, 144, 101–112. [Google Scholar] [CrossRef]
- Stephan, E. Functional neuroimaging of gastric distention. J. Gastrointest. Surg. 2003, 7, 740–749. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.-J.; Tomasi, D.; Backus, W.; Wang, R.; Telang, F.; Geliebter, A.; Korner, J.; Bauman, A.; Fowler, J.S.; Thanos, P.K.; et al. Gastric distention activates satiety circuitry in the human brain. NeuroImage 2008, 39, 1824–1831. [Google Scholar] [CrossRef]
- Wang, D.; He, X.; Zhao, Z.; Feng, Q.; Lin, R.; Sun, Y.; Zhan, C. Whole-brain mapping of the direct inputs and axonal projections of POMC and AgRP neurons. Front. Neuroanat. 2015, 9, 40. [Google Scholar]
- Harno, E.; Ramamoorthy, T.G.; Coll, A.P.; White, A. POMC: The Physiological Power of Hormone Processing. Physiol. Rev. 2018, 98, 2381–2430. [Google Scholar] [CrossRef]
- Chen, Y.; Lin, Y.-C.; Kuo, T.-W.; Knight, Z.A. Sensory Detection of Food Rapidly Modulates Arcuate Feeding Circuits. Cell 2015, 160, 829–841. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Essner, R.A.; Kosar, S.; Miller, O.H.; Lin, Y.-C.; Mesgarzadeh, S.; Knight, Z.A. Sustained NPY signaling enables AgRP neurons to drive feeding. eLife 2019, 8, 46348. [Google Scholar] [CrossRef]
- Betley, J.N.; Xu, S.; Cao, Z.F.H.; Gong, R.; Magnus, C.J.; Yu, Y.; Sternson, S.M. Neurons for hunger and thirst transmit a negative-valence teaching signal. Nature 2015, 521, 180–185. [Google Scholar] [CrossRef] [PubMed]
- Alkemade, A.; Yi, C.-X.; Pei, L.; Harakalova, M.; Swaab, D.; La Fleur, S.E.; Fliers, E.; Kalsbeek, A. AgRP and NPY Expression in the Human Hypothalamic Infundibular Nucleus Correlate with Body Mass Index, Whereas Changes in αMSH Are Related to Type 2 Diabetes. J. Clin. Endocrinol. Metab. 2012, 97, E925–E933. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holder, J.L., Jr.; Butte, N.F.; Zinn, A.R. Profound obesity associated with a balanced translocation that disrupts the SIM1 gene. Hum. Mol. Genet. 2000, 9, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Lubrano-Berthelier, C.; Durand, E.; Dubern, B.; Shapiro, A.; Dazin, P.; Weill, J.; Ferron, C.; Froguel, P.; Vaisse, C. Intracellular retention is a common characteristic of childhood obesity-associated MC4R mutations. Hum. Mol. Genet. 2003, 12, 145–153. [Google Scholar] [PubMed]
- Collet, T.H.; Dubern, B.; Mokrosinski, J.; Connors, H.; Keogh, J.M.; de Oliveira, E.M.; Henning, E.; Poitou-Bernert, C.; Michel Oppert, J.; Tounian, P.; et al. Evaluation of a melanocortin-4 receptor (MC4R) agonist (Setmelanotide) in MC4R deficiency. Mol. Metab. 2017, 6, 1321–1329. [Google Scholar]
- Haws, R.; Brady, S.; Davis, E.; Fletty, K.; Yuan, G.; Gordon, G.; Stewart, M.; Yanovski, J. Effect of setmelanotide, a melanocortin-4 receptor agonist, on obesity in Bardet-Biedl syndrome. Diabetes Obes. Metab. 2020, 22, 2133–2140. [Google Scholar] [CrossRef]
- Krashes, M.J.; Lowell, B.B.; Garfield, A.S. Melanocortin-4 receptor–regulated energy homeostasis. Nat. Neurosci. 2016, 19, 206–219. [Google Scholar] [CrossRef] [Green Version]
- Van der Klaauw, A.A.; von dem Hagen, E.A.; Keogh, J.M.; Henning, E.; O’Rahilly, S.; Lawrence, A.D.; Calder, A.J.; Farooqi, I.S. Obesity-Associated Melanocortin-4 Receptor Mutations Are Associated with Changes in the Brain Response to Food Cues. J. Clin. Endocrinol. Metab. 2014, 99, E2101–E2106. [Google Scholar]
- De Lecea, L.; Kilduff, T.S.; Peyron, C.; Gao, X.-B.; Foye, P.E.; Danielson, P.E.; Fukuhara, C.; Battenberg, E.L.F.; Gautvik, V.T.; Bartlett, F.S., III; et al. The hypocretins: Hypothalamus-specific peptides with neuroexcitatory activity. Proc. Natl. Acad. Sci. USA 1998, 95, 322–327. [Google Scholar] [CrossRef] [Green Version]
- Kilduff, T.S.; de Lecea, L. Mapping of the mRNAs for the hypocretin/orexin and melanin-concentrating hormone receptors: Networks of overlapping peptide systems. J. Comp. Neurol. 2001, 435, 1–5. [Google Scholar] [CrossRef]
- Marcus, J.N.; Aschkenasi, C.J.; Lee, C.E.; Chemelli, R.M.; Saper, C.B.; Yanagisawa, M.; Elmquist, J.K. Differential expression of orexin receptors 1 and 2 in the rat brain. J. Comp. Neurol. 2001, 435, 6–25. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, Y.; Katakura, N. Generation of masticatory rhythm in the brainstem. Neurosci. Res. 1995, 23, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Katschinski, M. Nutritional implications of cephalic phase gastrointestinal responses. Appetite 2000, 34, 189–196. [Google Scholar] [CrossRef] [PubMed]
- Thorpe, A.; Kotz, C. Orexin A in the nucleus accumbens stimulates feeding and locomotor activity. Brain Res. 2005, 1050, 156–162. [Google Scholar] [CrossRef]
- Page, K.A.; Chan, O.; Arora, J.; Belfort-DeAguiar, R.; Dzuira, J.; Roehmholdt, B.; Cline, G.W.; Naik, S.; Sinha, R.; Constable, R.; et al. Effects of Fructose vs Glucose on Regional Cerebral Blood Flow in Brain Regions Involved With Appetite and Reward Pathways. JAMA J. Am. Med Assoc. 2013, 309, 63–70. [Google Scholar] [CrossRef] [Green Version]
- Cornier, M.-A.; Von Kaenel, S.S.; Bessesen, D.H.; Tregellas, J.R. Effects of overfeeding on the neuronal response to visual food cues. Am. J. Clin. Nutr. 2007, 86, 965–971. [Google Scholar] [CrossRef] [Green Version]
- Matsuda, M.; Liu, Y.; Mahankali, S.; Pu, Y.; Wang, J.; DeFronzo, R.A.; Fox, P.T.; Gao, J.H. Altered hypothalamic function in response to glucose ingestion in obese humans. Diabetes 1999, 48, 1801–1806. [Google Scholar] [CrossRef]
- Contreras-Rodríguez, O.; Vilar-López, R.; Andrews, Z.B.; Navas, J.F.; Soriano-Mas, C.; Verdejo-Garcia, A. Altered cross-talk between the hypothalamus and non-homeostatic regions linked to obesity and difficulty to lose weight. Sci. Rep. 2017, 7, 9951. [Google Scholar] [CrossRef] [Green Version]
- Tsang, A.H.; Nuzzaci, D.; Darwish, T.; Samudrala, H.; Blouet, C. Nutrient sensing in the nucleus of the solitary tract mediates non-aversive suppression of feeding via inhibition of AgRP neurons. Mol. Metab. 2020, 42, 101070. [Google Scholar]
- Guinard, J.-X.; Mazzucchelli, R. The sensory perception of texture and mouthfeel. Trends Food Sci. Technol. 1996, 7, 213–219. [Google Scholar] [CrossRef]
- Scott, T.R.; Verhagen, J.V. Taste as a factor in the management of nutrition. Nutrition 2000, 16, 874–885. [Google Scholar] [CrossRef] [PubMed]
- Prescott, J. Comparisons of taste perceptions and preferences of Japanese and Australian consumers: Overview and implications for cross-cultural sensory research. Food Qual. Prefer. 1998, 9, 393–402. [Google Scholar] [CrossRef]
- Bertino, M.; Beauchamp, G.K.; Jen, K.-L.C. Rated taste perception in two cultural groups. Chem. Senses 1983, 8, 3–15. [Google Scholar] [CrossRef]
- Schoenfeld, M.; Neuer, G.; Tempelmann, C.; Schüßler, K.; Noesselt, T.; Hopf, J.-M.; Heinze, H.-J. Functional magnetic resonance tomography correlates of taste perception in the human primary taste cortex. Neuroscience 2004, 127, 347–353. [Google Scholar] [CrossRef]
- Lemon, C.H. It’s all a matter of taste: Gustatory processing and ingestive decisions. Mo. Med. 2010, 107, 247–251. [Google Scholar]
- Campos, C.A.; Bowen, A.J.; Schwartz, M.W.; Palmiter, R.D. Parabrachial CGRP Neurons Control Meal Termination. Cell Metab. 2016, 23, 811–820. [Google Scholar] [CrossRef] [Green Version]
- O’Doherty, J.; Rolls, E.; Francis, S.; Bowtell, R.; McGlone, F. Representation of Pleasant and Aversive Taste in the Human Brain. J. Neurophysiol. 2001, 85, 1315–1321. [Google Scholar] [CrossRef]
- Frank, G.K.; Kaye, W.H.; Carter, C.S.; Brooks, S.; May, C.; Fissell, K.; Stenger, V. The evaluation of brain activity in response to taste stimuli—a pilot study and method for central taste activation as assessed by event-related fMRI. J. Neurosci. Methods 2003, 131, 99–105. [Google Scholar] [CrossRef]
- Schmidt, L.; Skvortsova, V.; Kullen, C.; Weber, B.; Plassmann, H. How context alters value: The brain’s valuation and affective regulation system link price cues to experienced taste pleasantness. Sci. Rep. 2017, 7, 8098. [Google Scholar] [CrossRef] [Green Version]
- McClure, S.M.; Li, J.; Tomlin, D.; Cypert, K.S.; Montague, L.M.; Montague, P. Neural Correlates of Behavioral Preference for Culturally Familiar Drinks. Neuron 2004, 44, 379–387. [Google Scholar] [CrossRef] [Green Version]
- Plassmann, H.; O’Doherty, J.; Shiv, B.; Rangel, A. Marketing actions can modulate neural representations of experienced pleasantness. Proc. Natl. Acad. Sci. USA 2008, 105, 1050–1054. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masterson, T.D.; Kirwan, C.B.; Davidson, L.E.; LeCheminant, J.D. Neural reactivity to visual food stimuli is reduced in some areas of the brain during evening hours compared to morning hours: An fMRI study in women. Brain Imaging Behav. 2015, 10, 68–78. [Google Scholar] [CrossRef]
- Rothemund, Y.; Preuschhof, C.; Bohner, G.; Bauknecht, H.-C.; Klingebiel, R.; Flor, H.; Klapp, B.F. Differential activation of the dorsal striatum by high-calorie visual food stimuli in obese individuals. NeuroImage 2007, 37, 410–421. [Google Scholar] [CrossRef] [PubMed]
- Rozin, P. “Taste–smell confusions” and the duality of the olfactory sense. Percept. Psychophys. 1982, 31, 397–401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shepherd, G.M. Smell images and the flavour system in the human brain. Nature 2006, 444, 316–321. [Google Scholar] [CrossRef] [PubMed]
- Small, D.M.; Voss, J.; Mak, Y.E.; Simmons, K.B.; Parrish, T.; Gitelman, D. Experience-Dependent Neural Integration of Taste and Smell in the Human Brain. J. Neurophysiol. 2004, 92, 1892–1903. [Google Scholar] [CrossRef] [PubMed]
- Thomas, J.; Higgs, S.; Dourish, C.; Hansen, P.C.; Harmer, C.; McCabe, C. Satiation attenuates BOLD activity in brain regions involved in reward and increases activity in dorsolateral prefrontal cortex: An fMRI study in healthy volunteers. Am. J. Clin. Nutr. 2015, 101, 697–704. [Google Scholar] [CrossRef] [Green Version]
- Smeets, P.A.M.; De Graaf, C.; Stafleu, A.; Van Osch, M.J.P.; Nievelstein, R.A.J.; Van Der Grond, J. Effect of satiety on brain activation during chocolate tasting in men and women. Am. J. Clin. Nutr. 2006, 83, 1297–1305. [Google Scholar] [CrossRef]
- Del Parigi, A.; Chen, K.; Gautier, J.-F.; Salbe, A.D.; Pratley, R.E.; Ravussin, E.; Reiman, E.M.; Tataranni, P.A. Sex differences in the human brain’s response to hunger and satiation. Am. J. Clin. Nutr. 2002, 75, 1017–1022. [Google Scholar]
- Olds, J.; Milner, P. Positive reinforcement produced by electrical stimulation of septal area and other regions of rat brain. J. Comp. Physiol. Psychol. 1954, 47, 419–427. [Google Scholar] [CrossRef]
- Corbett, D.; Wise, R.A. Intracranial self-stimulation in relation to the ascending dopaminergic systems of the midbrain: A moveable electrode mapping study. Brain Res. 1980, 185, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Rossi, M.A.; Stuber, G.D. Overlapping Brain Circuits for Homeostatic and Hedonic Feeding. Cell Metab. 2018, 27, 42–56. [Google Scholar] [CrossRef] [PubMed]
- Morton, G.J.; Meek, T.H.; Schwartz, M.W. Neurobiology of food intake in health and disease. Nat. Rev. Neurosci. 2014, 15, 367–378. [Google Scholar] [CrossRef] [Green Version]
- Small, D.M.; Jones-Gotman, M.; Daghera, A. Feeding-induced dopamine release in dorsal striatum correlates with meal pleasantness ratings in healthy human volunteers. NeuroImage 2003, 19, 1709–1715. [Google Scholar] [CrossRef]
- Schultz, W. Dopamine reward prediction error coding. Dialog. Clin. Neurosci. 2016, 18, 23–32. [Google Scholar]
- Alvino, L.; Van Der Lubbe, R.; Joosten, R.A.; Constantinides, E. Which wine do you prefer? An analysis on consumer behaviour and brain activity during a wine tasting experience. Asia Pac. J. Mark. Logist. 2019, 32, 1149–1170. [Google Scholar] [CrossRef]
- Cruwys, T.; Bevelander, K.E.; Hermans, R. Social modeling of eating: A review of when and why social influence affects food intake and choice. Appetite 2015, 86, 3–18. [Google Scholar] [CrossRef]
- Berridge, K.C.; Ho, C.-Y.; Richard, J.; DiFeliceantonio, A. The tempted brain eats: Pleasure and desire circuits in obesity and eating disorders. Brain Res. 2010, 1350, 43–64. [Google Scholar] [CrossRef] [Green Version]
- Berridge, K.C.; Robinson, T.E.; Aldridge, J.W. Dissecting components of reward: ‘liking’, ‘wanting’, and learning. Curr. Opin. Pharmacol. 2009, 9, 65–73. [Google Scholar]
- Berridge, K.C.; Robinson, T.E. Liking, wanting, and the incentive-sensitization theory of addiction. Am. Psychol. 2016, 71, 670–679. [Google Scholar] [CrossRef]
- Volkow, N.D.; Wang, G.-J.; Baler, R.D. Reward, dopamine and the control of food intake: Implications for obesity. Trends Cogn. Sci. 2011, 15, 37–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, G.-J.; Volkow, N.D.; Fowler, J.S. The role of dopamine in motivation for food in humans: Implications for obesity. Expert Opin. Ther. Targets 2002, 6, 601–609. [Google Scholar] [CrossRef] [PubMed]
- Stice, E.; Yokum, S. Neural vulnerability factors that increase risk for future weight gain. Psychol. Bull. 2016, 142, 447–471. [Google Scholar] [CrossRef] [PubMed]
- Burger, K.S.; Stice, E. Variability in reward responsivity and obesity: Evidence from brain imaging studies. Curr. Drug Abus. Rev. 2011, 4, 182–189. [Google Scholar]
- Robinson, T.E.; Berridge, K.C. The neural basis of drug craving: An incentive-sensitization theory of addiction. Brain Res. Rev. 1993, 18, 247–291. [Google Scholar]
- Nederkoorn, C.; Braet, C.; Van Eijs, Y.; Tanghe, A.; Jansen, A. Why obese children cannot resist food: The role of impulsivity. Eat. Behav. 2006, 7, 315–322. [Google Scholar] [CrossRef]
- Finlayson, G.; King, N.; Blundell, J.E. Is it possible to dissociate ‘liking’and ‘wanting’for foods in humans? A novel experimental procedure. Physiol. Behav. 2007, 90, 36–42. [Google Scholar]
- Berthoud, H.-R.; Münzberg, H.; Morrison, C.D. Blaming the Brain for Obesity: Integration of Hedonic and Homeostatic Mechanisms. Gastroenterology 2017, 152, 1728–1738. [Google Scholar] [CrossRef] [Green Version]
- Volkow, N.D.; Wang, G.-J.; Fowler, J.S.; Tomasi, D. Addiction Circuitry in the Human Brain. Annu. Rev. Pharmacol. Toxicol. 2012, 52, 321–336. [Google Scholar] [CrossRef]
- Volkow, N.D.; Wang, G.-J.; Tomasi, D.; Baler, R.D. Obesity and addiction: Neurobiological overlaps. Obes. Rev. 2013, 14, 2–18. [Google Scholar] [CrossRef] [Green Version]
- Garcia, I.G.; Horstmann, A.; Jurado, M.A.; Garolera, M.; Chaudhry, S.; Margulies, D.; Villringer, A.; Neumann, J. Reward processing in obesity, substance addiction and non-substance addiction. Obes. Rev. 2014, 15, 853–869. [Google Scholar] [CrossRef]
- Lindgren, E.; Gray, K.; Miller, G.; Tyler, R.; Wiers, C.E.; Volkow, N.D.; Wang, G.J. Food addiction: A common neurobiological mechanism with drug abuse. Front. Biosci. Landmark Ed. 2018, 23, 811–836. [Google Scholar] [PubMed] [Green Version]
- Finlayson, G. Food addiction and obesity: Unnecessary medicalization of hedonic overeating. Nat. Rev. Endocrinol. 2017, 13, 493–498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Volkow, N.D.; O’Brien, C.P. Issues for DSM-V: Should Obesity Be Included as a Brain Disorder? Am. J. Psychiatry 2007, 164, 708–710. [Google Scholar] [CrossRef]
- Ziauddeen, H.; Farooqi, I.S.; Fletcher, P.C. Obesity and the brain: How convincing is the addiction model? Nat. Rev. Neurosci. 2012, 13, 279–286. [Google Scholar] [CrossRef]
- Avena, N.M.; Gearhardt, A.; Gold, M.S.; Wang, G.-J.; Potenza, M.N. Tossing the baby out with the bathwater after a brief rinse? The potential downside of dismissing food addiction based on limited data. Nat. Rev. Neurosci. 2012, 13, 514. [Google Scholar] [CrossRef] [Green Version]
- Coccurello, R.; Maccarrone, M. Hedonic Eating and the “Delicious Circle”: From Lipid-Derived Mediators to Brain Dopamine and Back. Front. Neurosci. 2018, 12, 271. [Google Scholar] [CrossRef] [Green Version]
- Berridge, K.C. The debate over dopamine’s role in reward: The case for incentive salience. Psychopharmacology 2007, 191, 391–431. [Google Scholar] [CrossRef]
- Epstein, L.H.; Temple, J.L.; Roemmich, J.N.; Bouton, M. Habituation as a determinant of human food intake. Psychol. Rev. 2009, 116, 384–407. [Google Scholar] [CrossRef]
- Carr, K.A.; Epstein, L.H. Relationship between food habituation and reinforcing efficacy of food. Learn. Motiv. 2011, 42, 165–172. [Google Scholar] [CrossRef] [Green Version]
- Epstein, L.H.; Carr, K.A. Food reinforcement and habituation to food are processes related to initiation and cessation of eating. Physiol. Behav. 2021, 239, 113512. [Google Scholar] [CrossRef] [PubMed]
- Lenoir, M.; Serre, F.; Cantin, L.; Ahmed, S.H. Intense Sweetness Surpasses Cocaine Reward. PLoS ONE 2007, 2, e698. [Google Scholar] [CrossRef] [Green Version]
- Avena, N.M.; Rada, P.; Hoebel, B.G. Evidence for sugar addiction: Behavioral and neurochemical effects of intermittent, excessive sugar intake. Neurosci. Biobehav. Rev. 2008, 32, 20–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Avena, N.M.; Rada, P.; Hoebel, B.G. Sugar and Fat Bingeing Have Notable Differences in Addictive-like Behavior. J. Nutr. 2009, 139, 623–628. [Google Scholar] [CrossRef] [PubMed]
- Figlewicz, D.; Evans, S.; Murphy, J.; Hoen, M.; Baskin, D. Expression of receptors for insulin and leptin in the ventral tegmental area/substantia nigra (VTA/SN) of the rat. Brain Res. 2003, 964, 107–115. [Google Scholar] [CrossRef] [PubMed]
- Leshan, R.L.; Opland, D.M.; Louis, G.W.; Leinninger, G.M.; Patterson, C.M.; Rhodes, C.J.; Münzberg, H.; Myers, M.G. Ventral Tegmental Area Leptin Receptor Neurons Specifically Project to and Regulate Cocaine- and Amphetamine-Regulated Transcript Neurons of the Extended Central Amygdala. J. Neurosci. 2010, 30, 5713–5723. [Google Scholar] [CrossRef]
- Fadel, J.; Deutch, A. Anatomical substrates of orexin–dopamine interactions: Lateral hypothalamic projections to the ventral tegmental area. Neuroscience 2002, 111, 379–387. [Google Scholar] [CrossRef]
- Davis, J.F.; Choi, D.L.; Shurdak, J.D.; Krause, E.G.; Fitzgerald, M.F.; Lipton, J.W.; Sakai, R.R.; Benoit, S.C. Central melanocortins modulate mesocorticolimbic activity and food seeking behavior in the rat. Physiol. Behav. 2011, 102, 491–495. [Google Scholar] [CrossRef] [Green Version]
- Alhadeff, A.L.; Rupprecht, L.E.; Hayes, M.R. GLP-1 Neurons in the Nucleus of the Solitary Tract Project Directly to the Ventral Tegmental Area and Nucleus Accumbens to Control for Food Intake. Endocrinology 2012, 153, 647–658. [Google Scholar] [CrossRef] [Green Version]
- Jerlhag, E.; Egecioglu, E.; Dickson, S.L.; Douhan, A.; Svensson, L.; Engel, J.A. Ghrelin administration into tegmental areas stimulates locomotor activity and increases extracellular concentration of dopamine in the nucleus accumbens. Addict. Biol. 2007, 12, 6–16. [Google Scholar] [CrossRef]
- Cason, A.M.; Smith, R.J.; Tahsili-Fahadan, P.; Moorman, D.; Sartor, G.C.; Aston-Jones, G. Role of orexin/hypocretin in reward-seeking and addiction: Implications for obesity. Physiol. Behav. 2010, 100, 419–428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dickson, S.; Shirazi, R.H.; Hansson, C.; Bergquist, F.; Nissbrandt, H.; Skibicka, K.P. The Glucagon-Like Peptide 1 (GLP-1) Analogue, Exendin-4, Decreases the Rewarding Value of Food: A New Role for Mesolimbic GLP-1 Receptors. J. Neurosci. 2012, 32, 4812–4820. [Google Scholar] [CrossRef] [PubMed]
- Stice, E.; Spoor, S.; Bohon, C.; Small, D.M. Relation between obesity and blunted striatal response to food is moderated by TaqIA A1 allele. Science 2008, 322, 449–452. [Google Scholar] [PubMed] [Green Version]
- Thanarajah, S.E.; Backes, H.; Di Feliceantonio, A.G.; Albus, K.; Cremer, A.L.; Hanssen, R.; Lippert, R.; Cornely, O.A.; Small, D.M.; Brüning, J.C.; et al. Food Intake Recruits Orosensory and Post-ingestive Dopaminergic Circuits to Affect Eating Desire in Humans. Cell Metab. 2019, 29, 695–706.e4. [Google Scholar] [CrossRef] [Green Version]
- Pignatelli, M.; Bonci, A. Role of Dopamine Neurons in Reward and Aversion: A Synaptic Plasticity Perspective. Neuron 2015, 86, 1145–1157. [Google Scholar] [CrossRef] [Green Version]
- Heisler, L.K.; Jobst, E.E.; Sutton, G.M.; Zhou, L.; Borok, E.; Thornton-Jones, Z.; Liu, H.Y.; Zigman, J.M.; Balthasar, N.; Kishi, T.; et al. Serotonin Reciprocally Regulates Melanocortin Neurons to Modulate Food Intake. Neuron 2006, 51, 239–249. [Google Scholar] [CrossRef] [Green Version]
- Kranz, G.; Kasper, S.; Lanzenberger, R. Reward and the serotonergic system. Neuroscience 2010, 166, 1023–1035. [Google Scholar] [CrossRef]
- Kaye, W.H.; Frank, G.K.; Meltzer, C.C.; Price, J.C.; McConaha, C.W.; Crossan, P.J.; Klump, K.L.; Rhodes, L. Altered Serotonin 2A Receptor Activity in Women Who Have Recovered From Bulimia Nervosa. Am. J. Psychiatry 2001, 158, 1152–1155. [Google Scholar] [CrossRef]
- Frank, G.; Kaye, W.H.; Meltzer, C.C.; Price, J.C.; Greer, P.; McConaha, C.; Skovira, K. Reduced 5-HT2A receptor binding after recovery from anorexia nervosa. Biol. Psychiatry 2002, 52, 896–906. [Google Scholar] [CrossRef]
- Audenaert, K.; Van Laere, K.; Dumont, F.; Vervaet, M.; Goethals, I.; Slegers, G.; Mertens, J.; Van Heeringen, C.; Dierckx, R.A. Decreased 5-HT2a receptor binding in patients with anorexia nervosa. J. Nucl. Med. 2003, 44, 163–169. [Google Scholar]
- Haahr, M.; Rasmussen, P.; Madsen, K.; Marner, L.; Ratner, C.; Gillings, N.; Baaré, W.; Knudsen, G.M. Obesity is associated with high serotonin 4 receptor availability in the brain reward circuitry. NeuroImage 2012, 61, 884–888. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, S.; Doya, K.; Okada, G.; Ueda, K.; Okamoto, Y.; Yamawaki, S. Prediction of immediate and future rewards differentially recruits cortico-basal ganglia loops. Nat. Neurosci. 2004, 7, 887–893. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, S.C.; Schweighofer, N.; Asahi, S.; Shishida, K.; Okamoto, Y.; Yamawaki, S.; Doya, K. Serotonin Differentially Regulates Short- and Long-Term Prediction of Rewards in the Ventral and Dorsal Striatum. PLoS ONE 2007, 2, e1333. [Google Scholar] [CrossRef]
- Thomas, J.; Dourish, C.; Tomlinson, J.; Hassan-Smith, Z.; Hansen, P.C.; Higgs, S. The 5-HT2C receptor agonist meta-chlorophenylpiperazine (mCPP) reduces palatable food consumption and BOLD fMRI responses to food images in healthy female volunteers. Psychopharmacology 2017, 235, 257–267. [Google Scholar] [CrossRef] [Green Version]
- Farr, O.M.; Upadhyay, J.; Gavrieli, A.; Camp, M.; Spyrou, N.; Kaye, H.; Mathew, H.; Vamvini, M.; Koniaris, A.; Kilim, H.; et al. Lorcaserin Administration Decreases Activation of Brain Centers in Response to Food Cues and These Emotion- and Salience-Related Changes Correlate With Weight Loss Effects: A 4-Week-Long Randomized, Placebo-Controlled, Double-Blind Clinical Trial. Diabetes 2016, 65, 2943–2953. [Google Scholar] [CrossRef] [Green Version]
- Bello, N.T.; Yeomans, B.L. Safety of pharmacotherapy options for bulimia nervosa and binge eating disorder. Expert Opin. Drug Saf. 2018, 17, 17–23. [Google Scholar] [CrossRef]
- Rahman, S.K.; Uyama, T.; Hussain, Z.; Ueda, N. Roles of Endocannabinoids and Endocannabinoid-Like Molecules in Energy Homeostasis and Metabolic Regulation: A Nutritional Perspective. Annu. Rev. Nutr. 2021, 41, 177–202. [Google Scholar] [CrossRef]
- Piomelli, D.; Tagne, A.M. Endocannabinoid-Based Therapies. Annu. Rev. Pharmacol. Toxicol. 2022, 62, 483–507. [Google Scholar] [CrossRef]
- Verme, J.L.; Gaetani, S.; Fu, J.; Oveisi, F.; Burton, K.; Piomelli, D. Regulation of food intake by oleoylethanolamide. Cell. Mol. Life Sci. 2005, 62, 708–716. [Google Scholar] [CrossRef] [Green Version]
- Piomelli, D. A fatty gut feeling. Trends Endocrinol. Metab. 2013, 24, 332–341. [Google Scholar] [CrossRef] [Green Version]
- Sihag, J.; Jones, P.J.H. Oleoylethanolamide: The role of a bioactive lipid amide in modulating eating behaviour. Obes. Rev. 2017, 19, 178–197. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.D.; Azari, E.K.; Ayala, J.E. Oleoylethanolamide: A fat ally in the fight against obesity. Physiol. Behav. 2017, 176, 50–58. [Google Scholar] [CrossRef] [PubMed]
- TuTunchi, H.; Ostadrahimi, A.; Saghafi-Asl, M.; Maleki, V. The effects of oleoylethanolamide, an endogenous PPAR-α agonist, on risk factors for NAFLD: A systematic review. Obes. Rev. 2019, 20, 1057–1069. [Google Scholar] [CrossRef] [PubMed]
- Veilleux, A.; Di Marzo, V.; Silvestri, C. The Expanded Endocannabinoid System/Endocannabinoidome as a Potential Target for Treating Diabetes Mellitus. Curr. Diabetes Rep. 2019, 19, 117. [Google Scholar] [CrossRef]
- Sihag, J.; Di Marzo, V. (Wh)olistic (E)ndocannabinoidome-Microbiome-Axis Modulation through (N)utrition (WHEN) to Curb Obesity and Related Disorders. Lipids Health Dis. 2022, 21, 1–21. [Google Scholar] [CrossRef]
- Iannotti, F.A.; Di Marzo, V. The gut microbiome, endocannabinoids and metabolic disorders. J. Endocrinol. 2021, 248, R83–R97. [Google Scholar] [CrossRef] [PubMed]
- Di Marzo, V. Endocannabinoids: Synthesis and degradation. In Reviews of Physiology Biochemistry and Pharmacology; Springer: Berlin/Heidelberg, Germany, 2006; pp. 1–24. [Google Scholar]
- Mackie, K. Distribution of cannabinoid receptors in the central and peripheral nervous system. In Cannabinoids; Springer: Berlin/Heidelberg, Germany, 2005; pp. 299–325. [Google Scholar]
- Ceccarini, J.; Weltens, N.; Ly, H.G.; Tack, J.; Van Oudenhove, L.; Van Laere, K. Association between cerebral cannabinoid 1 receptor availability and body mass index in patients with food intake disorders and healthy subjects: A [(18)F]MK-9470 PET study. Transl. Psychiatry 2016, 6, e853. [Google Scholar] [CrossRef] [Green Version]
- Di Marzo, V.; Goparaju, S.K.; Wang, L.; Liu, J.; Bátkai, S.; Járai, Z.; Fezza, F.; Miura, G.I.; Palmiter, R.D.; Sugiura, T.; et al. Leptin-regulated endocannabinoids are involved in maintaining food intake. Nature 2001, 410, 822–825. [Google Scholar] [CrossRef] [PubMed]
- Kirkham, T.C.; Williams, C.; Fezza, F.; Di Marzo, V. Endocannabinoid levels in rat limbic forebrain and hypothalamus in relation to fasting, feeding and satiation: Stimulation of eating by 2-arachidonoyl glycerol. J. Cereb. Blood Flow Metab. 2002, 136, 550–557. [Google Scholar] [CrossRef]
- Sam, A.H.; Salem, V.; Ghatei, M.A. Rimonabant: From RIO to Ban. J. Obes. 2011, 2011, 432607. [Google Scholar] [CrossRef]
- Horder, J.; Harmer, C.J.; Cowen, P.J.; McCabe, C. Reduced neural response to reward following 7 days treatment with the cannabinoid CB1 antagonist rimonabant in healthy volunteers. Int. J. Neuropsychopharmacol. 2010, 13, 1103–1113. [Google Scholar] [CrossRef] [PubMed]
- Tudge, L.; Williams, C.; Cowen, P.J.; McCabe, C. Neural Effects of Cannabinoid CB1 Neutral Antagonist Tetrahydrocannabivarin on Food Reward and Aversion in Healthy Volunteers. Int. J. Neuropsychopharmacol. 2015, 18, pyu094. [Google Scholar] [CrossRef] [Green Version]
- Lord, J.A.H.; Waterfield, A.A.; Hughes, J.; Kosterlitz, H.W. Endogenous opioid peptides: Multiple agonists and receptors. Nature 1977, 267, 495–499. [Google Scholar] [CrossRef] [PubMed]
- Peciña, S.; Smith, K.S. Hedonic and motivational roles of opioids in food reward: Implications for overeating disorders. Pharmacol. Biochem. Behav. 2010, 97, 34–46. [Google Scholar] [CrossRef] [PubMed]
- Nummenmaa, L.; Saanijoki, T.; Tuominen, L.; Hirvonen, J.; Tuulari, J.J.; Nuutila, P.; Kalliokoski, K. μ-opioid receptor system mediates reward processing in humans. Nat. Commun. 2018, 9, 1500. [Google Scholar] [CrossRef]
- Yanovski, S.Z.; Yanovski, J. Naltrexone Extended-Release Plus Bupropion Extended-Release for Treatment of Obesity. JAMA: J. Am. Med. Assoc. 2015, 313, 1213–1214. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.-J.; Tomasi, D.; Volkow, N.D.; Wang, R.; Telang, F.; Caparelli, E.C.; Dunayevich, E. Effect of combined naltrexone and bupropion therapy on the brain’s reactivity to food cues. Int. J. Obes. 2013, 38, 682–688. [Google Scholar] [CrossRef] [Green Version]
- Murray, E.; Brouwer, S.; McCutcheon, R.; Harmer, C.; Cowen, P.; McCabe, C. Opposing neural effects of naltrexone on food reward and aversion: Implications for the treatment of obesity. Psychopharmacology 2014, 231, 4323–4335. [Google Scholar] [CrossRef]
- Berthoud, H.-R. The neurobiology of food intake in an obesogenic environment. Proc. Nutr. Soc. 2012, 71, 478–487. [Google Scholar] [CrossRef] [Green Version]
- Logothetis, N.K. What we can do and what we cannot do with fMRI. Nature 2008, 453, 869–878. [Google Scholar]
- Kropotov, J.D. Functional Neuromarkers for Psychiatry; Elsevier BV: Amsterdam, The Netherlands, 2016; pp. 27–30. [Google Scholar]
- Newberg, A.B.; Alavi, A. Single Photon Emission Computed Tomography☆, in Reference Module in Neuroscience and Biobehavioral Psychology; Elsevier: Amsterdam, The Netherlands, 2017. [Google Scholar]
- Smeets, P.A.M.; Dagher, A.; Hare, T.; Kullmann, S.; van der Laan, L.N.; Poldrack, R.; Preissl, H.; Small, D.; Stice, E.; Veldhuizen, M.G. Good practice in food-related neuroimaging. Am. J. Clin. Nutr. 2019, 109, 491–503. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Definitions | |
|---|---|
| Appetite | Desire to fulfill energetic needs, divided into hunger, satiety, and satiation |
| Hunger | Drive to consume |
| Satiation | The process that brings an eating episode to an end (intra-meal inhibition) |
| Satiety | The process that inhibits eating or hunger in the postprandial period (inter-meal inhibition) |
| Fullness | The visceral sensation of gastric distention |
| Reward | The stimulus for which animals are willing to work for |
| Hedonism | Derived from the Greek hēdonikós, “pleasurable”; a sensation of positive feelings |
| Hedonic Eating | Eating solely to elicit pleasurable feelings and/or to escape from anhedonic states, disregarding the metabolic status or the nutritional value of the food consumed. |
| Appetition | The post-oral process where nutrient properties of food promote intake beyond inhibition processes (e.g., satiety and satiation). |
| Preference | Selection among different options based on subjective liking |
| Incentive Salience | The cognitive process of “wanting” a rewarding stimulus based on its attributed salient characteristics |
| Reward prediction error | Difference between the received and predicted reward from a stimulus |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Campos, A.; Port, J.D.; Acosta, A. Integrative Hedonic and Homeostatic Food Intake Regulation by the Central Nervous System: Insights from Neuroimaging. Brain Sci. 2022, 12, 431. https://doi.org/10.3390/brainsci12040431
Campos A, Port JD, Acosta A. Integrative Hedonic and Homeostatic Food Intake Regulation by the Central Nervous System: Insights from Neuroimaging. Brain Sciences. 2022; 12(4):431. https://doi.org/10.3390/brainsci12040431
Chicago/Turabian StyleCampos, Alejandro, John D. Port, and Andres Acosta. 2022. "Integrative Hedonic and Homeostatic Food Intake Regulation by the Central Nervous System: Insights from Neuroimaging" Brain Sciences 12, no. 4: 431. https://doi.org/10.3390/brainsci12040431