
Emergent Neuroimaging Findings for Written Expression in Children: A Scoping Review


Abstract
:1. Introduction
Objective of the Scoping Review
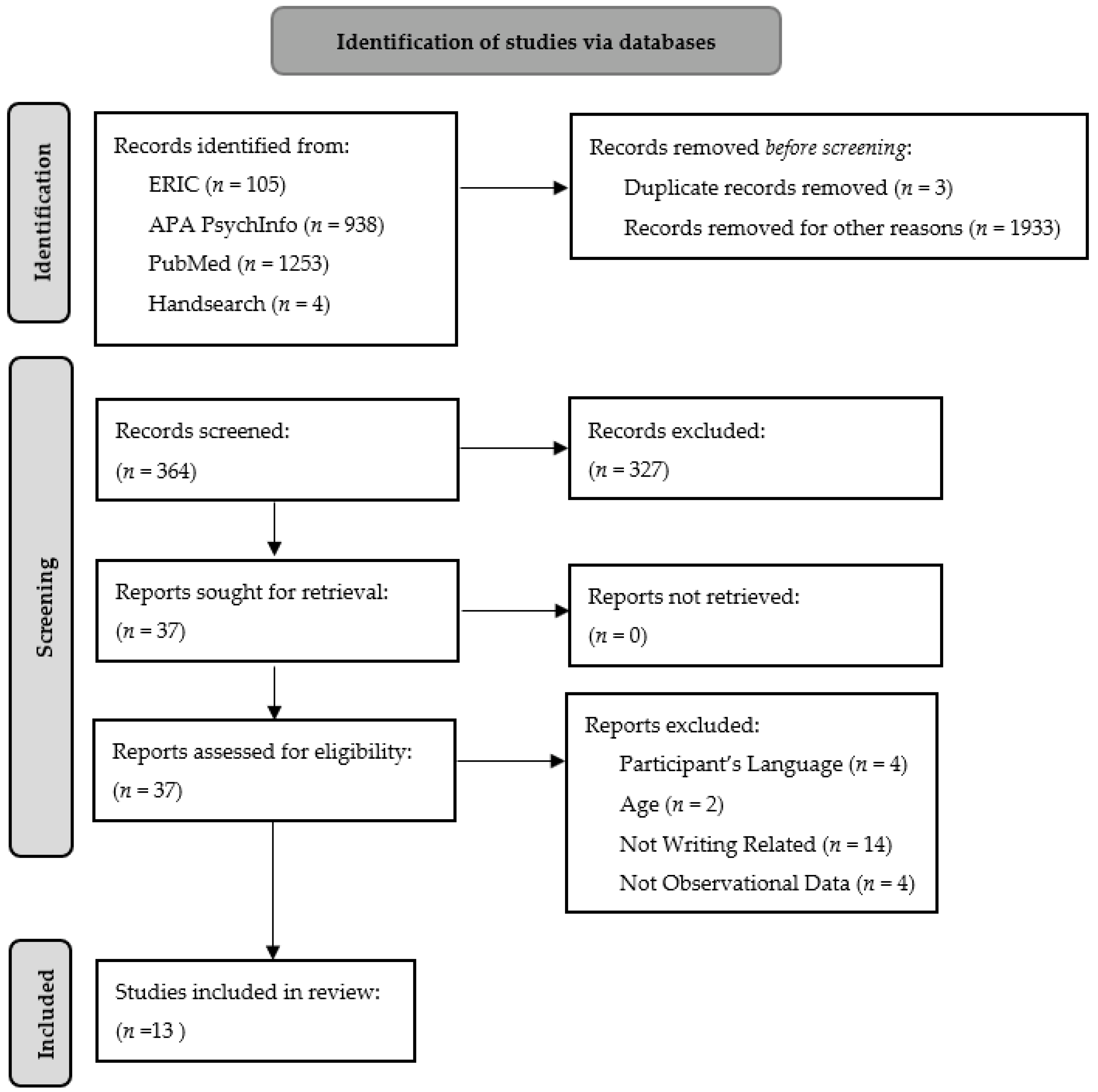
2. Methods
3. Results
3.1. Handwriting
3.2. Spelling
3.3. Idea Generation
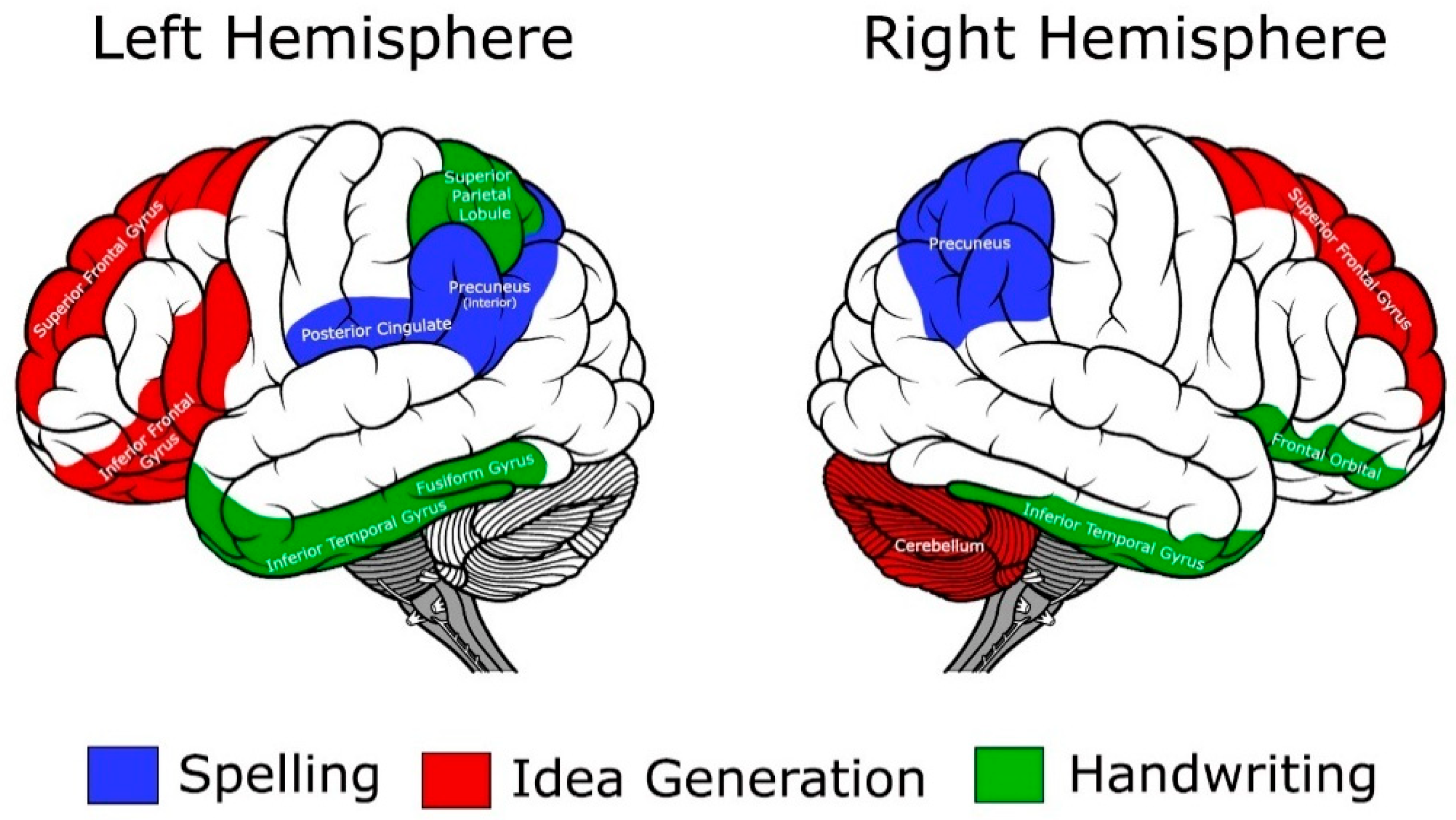
3.4. Summary
3.5. Related Research: Brain Changes following Educational Intervention for Writing Problems
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Nystrand, M. The social and historical context for writing research. In Handbook of Writing Research; MacArthur, C., Graham, S., Fitzgerald, J., Eds.; The Guilford Press: New York, NY, USA, 2006; pp. 11–27. [Google Scholar]
- MacArthur, C.A.; Graham, S. Writing research from a cognitive perspective. In Handbook of Writing Research, 2nd ed.; MacArthur, C., Graham, S., Fitzgerald, J., Eds.; The Guilford Press: New York, NY, USA, 2016; pp. 24–40. [Google Scholar]
- Munn, Z.; Peters, M.D.J.; Stern, C.; Tufanaru, C.; McArthur, A.; Aromataris, E. Systematic review or scoping review? Guidance for authors when choosing between a systematic or scoping review approach. BMC Med. Res. Method 2018, 18, 143. [Google Scholar] [CrossRef] [PubMed]
- Tricco, A.C.; Lillie, E.; Zarin, W.; O’Brien, K.K.; Colquhoun, H.; Levac, D.; Moher, D.; Peters, M.D.J.; Horsley, T.; Weeks, L.; et al. PRISMA Extension for Scoping Reviews (PRISMA-ScR): Checklist and explanation. Ann. Intern. Med. 2018, 169, 467–473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borkowska, A.R.; Francuz, P.; Soluch, P.; Wolak, T. Brain activation in teenagers with isolated spelling disorder during tasks involving spelling assessment and comparison of pseudowords. fMRI study. Brain Dev. 2014, 36, 786–793. [Google Scholar] [CrossRef] [PubMed]
- Richards, T.; Berninger, V.; Stock, P.; Altemeier, L.; Trivedi, P.; Maravilla, K. Differences between good and poor child writers on fMRI contrasts for writing newly taught and highly practiced letter forms. Read. Writ. 2011, 24, 493–516. [Google Scholar] [CrossRef] [PubMed]
- Gimenez, P.; Bugescu, N.; Black, J.; Hancock, R.; Pugh, K.; Nagamine, M.; Kutner, E.; Mazaika, P.; Hendren, R.; McCandliss, B.; et al. Neuroimaging correlates of handwriting quality as children learn to read and write. Front. Hum. Neurosci. 2014, 8, 155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richards, T.L.; Grabowski, T.J.; Boord, P.; Yagle, K.; Askren, M.; Mestre, Z.; Robinson, P.; Welker, O.; Gulliford, D.; Nagy, W.; et al. Contrasting brain patterns of writing-related DTI parameters, fMRI connectivity, and DTI-fMRI connectivity correlations in children with and without dysgraphia or dyslexia. Neuroimage Clin. 2015, 8, 408–421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richards, T.L.; Abbott, R.D.; Yagle, K.; Peterson, D.; Raskind, W.; Berninger, V.W. Self-government of complex reading and writing brains informed by cingulo-opercular network for adaptive control and working memory components for language learning. J. Syst. Integr. Neurosci. 2017, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richards, T.; Abbott, R.D.; Berninger, V.W. Relationships between Presence or Absence of ADHD and fMRI Connectivity Writing Tasks in Children with Dysgraphia. J. Nat. Sci. 2016, 2, e270. [Google Scholar]
- Richards, T.; Berninger, V.; Stock, P.; Altemeier, L.; Trivedi, P.; Maravilla, K. fMRI sequential-finger movement activation differentiating good and poor writers. J. Clin. Exp. Neuropsychol. 2009, 31, 967–983. [Google Scholar] [CrossRef] [PubMed]
- Bitan, T.; Burman, D.D.; Chou, T.L.; Lu, D.; Cone, N.E.; Cao, F.; Bigio, J.D.; Booth, J.R. The interaction between orthographic and phonological information in children: An fMRI study. Hum. Brain Mapp. 2007, 28, 880–891. [Google Scholar] [CrossRef] [Green Version]
- Booth, J.R.; Mehdiratta, N.; Burman, D.D.; Bitan, T. Developmental increases in effective connectivity to brain regions involved in phonological processing during tasks with orthographic demands. Brain Res. 2008, 1189, 78–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berninger, V.W.; Richards, T.; Abbott, R.D. Differential diagnosis of dysgraphia, dyslexia, and OWL LD: Behavioral and neuroimaging evidence. Read. Writ. 2015, 28, 1119–1153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richards, T.; Berninger, V.; Fayol, M. fMRI activation differences between 11-year old good and poor spellers’ access in working memory to temporary and long-term orthographic representations. J. Neurolinguist. 2009, 22, 327–353. [Google Scholar] [CrossRef]
- Richards, T.; Berninger, V.; Winn, W.; Swanson, H.L.; Stock, P.; Liang, O.; Abbott, R. Difference in fMRI activation between children with and without spelling disability on 2-back/0-back working memory contrast. J. Writ. Res. 2009, 1, 93–123. [Google Scholar] [CrossRef]
- Berninger, V.; Berninger, V.W.; Richards, T.L.; Stock, P.; Abbott, R.; Trivedi, P.; Altemeier, L.; Hayes, J.R. fMRI activation related to nature of ideas generated and differences between good and poor writers during idea generation. Br. J. Ed. Psychol. Mono. 2009, 2, 77–93. [Google Scholar] [CrossRef]
- Wallis, P.; Richards, T.; Boord, P.; Abbott, R.; Berninger, V. Relationships between Translation and Transcription Processes during fMRI Connectivity Scanning and Coded Translation and Transcription in Writing Products after Scanning in Children with and without Transcription Disabilities. Creat. Educ. 2017, 8, 716–748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richards, T.L.; Berninger, V.W.; Yagle, K.J.; Abbott, R.D.; Peterson, D.J. Changes in DTI Diffusivity and fMRI Connectivity Cluster Coefficients for Students with and without Specific Learning Disabilities in Written Language: Brain’s Response to Writing Instruction. J. Nat. Sci. 2017, 3, e350. [Google Scholar]
- Shah, C.; Erhard, K.; Ortheil, H.J.; Kaza, E.; Kessler, C.; Lotze, M. Neural correlates of creative writing: An fMRI study. Hum. Brain Mapp. 2013, 34, 1088–1101. [Google Scholar] [CrossRef] [PubMed]
- Segal, E.; Petrides, M. The anterior superior parietal lobule and its interactions with language and motor areas during writing. Eur. J. Neurosci. 2012, 35, 309–322. [Google Scholar] [CrossRef]
- Rapp, B.; Dufor, O. The neurotopography of written word production: An fMRI investigation of the distribution of sensitivity to length and frequency. J. Cogn. Neurosci. 2011, 23, 4067–4081. [Google Scholar] [CrossRef]
- Roux, F.-E.; Dufor, O.; Giussani, C.; Wamain, Y.; Draper, L.; Longcamp, M.; Demonet, J.-F. The graphemic/motor frontal area Exner’s area revisited. Ann. Neurol. 2009, 66, 537–545. [Google Scholar] [CrossRef] [PubMed]
- Reitz, F.; Richards, T.; Wu, K.; Boord, P.; Askren, M.; Lewis, T.; Berninger, V. A low-cost, computer-interfaced drawing pad for FMRI studies of dysgraphia and dyslexia. Sensors 2013, 13, 5099–5108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tam, F.; Churchill, N.W.; Strother, S.C.; Graham, S.J. A new tablet for writing and drawing during functional MRI. Hum. Brain Mapp. 2011, 32, 240–248. [Google Scholar] [CrossRef]
- Tomasi, D.; Caparelli, E.C.; Chang, L.; Ernst, T. fMRI-acoustic noise alters brain activation during working memory tasks. NeuroImage 2005, 27, 377–386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perrachione, T.K.; Ghosh, S.S. Optimized design and analysis of sparse-sampling fMRI experiments. Front. Neurosci. 2013, 7, 55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berninger, V.; Winn, W. Implications of advancements in brain research and technology for writing development, writing instruction, and educational evolution. In Handbook of Writing Research; MacArthur, C., Graham, S., Fitzgerald, J., Eds.; The Guilford Press: New York, NY, USA, 2006; pp. 96–114. [Google Scholar]
- Hayes, J. New directions in writing theory. In Handbook of Writing Research; MacArthur, C., Graham, S., Fitzgerald, J., Eds.; The Guilford Press: New York, NY, USA, 2006; pp. 28–40. [Google Scholar]



{kind=link}
{kind=link}
| Author(s) | Date | Population | Sample Size | Neuroimaging Methodology | Writing Task | Brain Regions Observed |
|---|---|---|---|---|---|---|
| Handwriting | ||||||
| Richards et al. [6] | 2011 | 5th graders | 20 | fMRI BOLD activation was measured during on (writing a newly taught pseudo-letter) and off (writing the real letter ‘a’) tasks | Writing pseudo-letter or real letter ‘a’ with a wooden stylus on a touch pad on chest at midline at a steady pace practiced outside the scanner | Poor writers showed significant activation in many more brain regions than good writers, as well as activation in unique regions. Good writers activated significantly more in the left fusiform gyrus than did poor writers. |
| Giminez et al. [7] | 2014 | 5- and 6-year-olds | 51 (5 left-handed students were later excluded) | fMRI activation was measured during 3 matching tasks; in one condition, stimuli were presented simultaneously, and after a small delay in the other condition | Phonological processing task; non-verbalize visual-symbol matching task; color-matching task | Poorer handwriting was associated with stronger activation of the right pars triangularis of the inferior frontal gyrus. |
| Richards et al. [8] | 2015 | Grades 4–9 (M = 12 yrs, 3 mos) | 40 (15 F, 25 M); Typical = 9; Dysgraphia = 14; Dyslexia = 17) | Diffusion Tensor Imaging (DTI) scans and fMRI connectivity scales | Resting state, alphabet writing task, spelling writing task, planning task (planned in task, then completed composition outside of scanner) | On structural connectivity, Dysgraphia Group showed less white matter integrity in the bilateral anterior thalamic radiation, left cingulate, and forceps minor than either of the other 2 groups; and less integrity in the left cortical spinal tract when compared to Typicals. The Dysgraphia Group showed higher radial diffusivity in 7 white mater tracts in the left hemisphere than Dyslexia Group. For functional connectivity, the Dysgraphia Group showed greater functional connectivity during planning tasks in left-occipital temporal, left inferior frontal gyrus, and left precuneus, but not during the resting task. For Dysgraphia Group, DTI-fMRI functional connectivity correlations between the left supramarginal gyrus and the left cingulum the anterior cingulate gyrus, and between the left inferior frontal regions and the superior parietal lobule, superior frontal gyrus, and the precuneus cortex. No differences between Typical versus Dysgraphia Group on the resting condition or alphabet writing task, but did show greater functional connectivity than Typicals on planning for composing. |
| Richards, Abbott, Yagle, Peterson et al. [9] | 2017 | Grades 4-9 | 42 | MRI, fMRI | Alphabet writing, spelling fill in letter task, planning task | Found significant fMRI connectivity with this network was uncovered in left cingulate gyrus during the fMRI alphabet writing task. |
| Richards, Abbott, and Berninger [10] | 2016 | Grades 4–9; dysgraphia and ADHD | 13 | fMRI | Resting state (mind wandering), alphabet writing, spelling fill in letter task, planning task | Presence of ADHD was significantly related to the degree of brain connectivity across 3 of 4 writing tasks. For mind wandering, significant correlations for the left occipital temporal with fusiform 2; left supramarginal with fusiform 2. For the transcription writing tasks, significant correlations between the left supramarginal region and Broca’s area for both alphabet writing and spelling tasks. For the planning task, no significant correlations were observed. |
| Richards, Berninger, Stock et al. [11] | 2009 | Fifth graders | 20 | fMRI BOLD activation was measured during on and off tasks | Finger tapping with and without sequencing | Left superior parietal, right inferior frontal orbital, right and left inferior temporal areas were found to be associated in good but not poor writers. BOLD activation in these areas was significantly correlated with handwriting, spelling, and composing. |
| Spelling | ||||||
| Bitan et al. [12] | 2007 | 9–15-year-olds | 38 | fMRI BOLD activation was measured during the task | Words spelled the same or not the same | During the spelling task, the left inferior/middle frontal gyri, superior/medial frontal gyri, superior/medial temporal gyri, thalamus, cuneus/calcarine sulcus, middle temporal gyrus, and the left inferior parietal lobule were active. |
| Booth et al. [13] | 2007 | 9–15-year-olds | 48 children completed the visual spelling task | fMRI BOLD activation was measured during the visual spelling task | Visual spelling | On the visual spelling task, developmental increases were found in the effective connectivity from calcarine to the superior temporal gyrus. |
| Berninger et al. [14] | 2015 | 9–15-year-olds, grades 4–9 | 45 children (controls n = 9, dysgraphia n = 14, dyslexia n = 17, OWL LD n = 5) | T2-weighted fMRI BOLD activation was used during a word-specific spelling task, deciding whether a letter string pronounced as a real word with meaning is a correctly spelled real word. fMRI looked at 4 regions: left occipital temporal, left supramarginal gyrus, left precuneus, and left inferior frontal gyrus | Participants were asked to complete the word judgement task on which they had been trained prior to scanning | OWL LD had the least connectivity in left occipital temporal seed point and left supramarginal gyrus, OWL LD had the least connectivity. OWL LD were similar to controls in the left precuneus seed point, OWL LD were similar to controls. OWL LD had slightly more connectivity than controls in the left inferior frontal seed point, OWL LD had slightly more connectivity than controls. In all seed points, dysgraphia had more connectivity than controls, and dyslexia had the most connectivity. |
| Richards, Berninger, and Fayol [15] | 2009 | 10–12-year-olds, mean age of 11 years, 4 months | 19 (12 good writers, 7 poor writers) | Used structural MRI scans and fMRI scans during on and off tasks | Real-word spelling judgment and pseudoword spelling judgment | Significant correlations between WIAT II composition and brain activation in 3 areas: left posterior cingulated, left precuneus, and right precuneus. Positive correlations for good spellers’ brain activation and negative correlations for poor spellers’ activation |
| Richards, Berninger, Winn et al. [16] | 2009 | 10- and 11-year-olds | 30 (10 good spellers, 20 with spelling disability) | Structural MRI and fMRI scans measured during presentation of stimuli | 0-back control and 2-back condition using colored pictures of sea creatures | Poor spellers activated more than the good spellers in the bilateral superior frontal, middle frontal, inferior frontal, anterior cingulum, left postcentral, and right superior frontal. Suggested differential activation of regions associated with working memory. |
| Idea Generation/Planning | ||||||
| Berninger et al. [17] | 2009 | 7 and 9 years old during intervention, 10 years old during the fMRI scanning | 20 (12 good writers, 8 poor writers) | Structural MRI and fMRI scans measured during presentation of verbal task | On/off tasks. 1) Off task: rest. 2) On-task: prompt was “Think about what you learned this summer that you have never learned in school. In a little while, you will leave the scanner and write about this topic.” | Good writers had more activation in brain regions associated with access to concepts and higher order cognition (left and right superior frontal gyrus), language, and executive functions related to language (left inferior frontal), working memory (right middle frontal orbital gyrus), and coordination (right cerebellum); poor writers had more activation in the left hemisphere, associated with working memory (left middle frontal gyrus); suggests that poor writers are inefficient in engaging working memory during idea generation. |
| Wallis et al. [18] | 2017 | Students with transcription disabilities (dysgraphia and dyslexia); controls; grades 4–9 | 39 | fMRI | Resting state (mind wandering), alphabet writing, spelling fill in letter task, planning task | For the dysgraphia group, fMRI connectivity for 3 of 4 tasks was significantly related to one or more of the composition outcomes created outside the scanner. Handwriting was the only exception. The significant correlations for spelling were observed from left precuneus with the fusiform gyrus and amygdala, and from left inferior frontal gyrus with Broca’s area and fusiform gyrus. For planning, the connectivity was observed with left precuneus with cingulate, from left inferior frontal to hippocampus, and from left precuneus to fusiform gyrus. For resting state, significant correlations were noted from left occipital temporal regions with hippocampus, from left supramarginal area with hippocampus, from left precuneus with hippocampus, from left inferior frontal with hippocampus, and from left precuneus with fusiform gyrus. The patterns of correlation also differed across groups. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Costa, L.-J.C.; Spencer, S.V.; Hooper, S.R. Emergent Neuroimaging Findings for Written Expression in Children: A Scoping Review. Brain Sci. 2022, 12, 406. https://doi.org/10.3390/brainsci12030406
Costa L-JC, Spencer SV, Hooper SR. Emergent Neuroimaging Findings for Written Expression in Children: A Scoping Review. Brain Sciences. 2022; 12(3):406. https://doi.org/10.3390/brainsci12030406
Chicago/Turabian StyleCosta, Lara-Jeane C., Sarah V. Spencer, and Stephen R. Hooper. 2022. "Emergent Neuroimaging Findings for Written Expression in Children: A Scoping Review" Brain Sciences 12, no. 3: 406. https://doi.org/10.3390/brainsci12030406