
Influence of Motor and Cognitive Tasks on Time Estimation

 ,
, 

Abstract
:
1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Apparatus and Set-Up
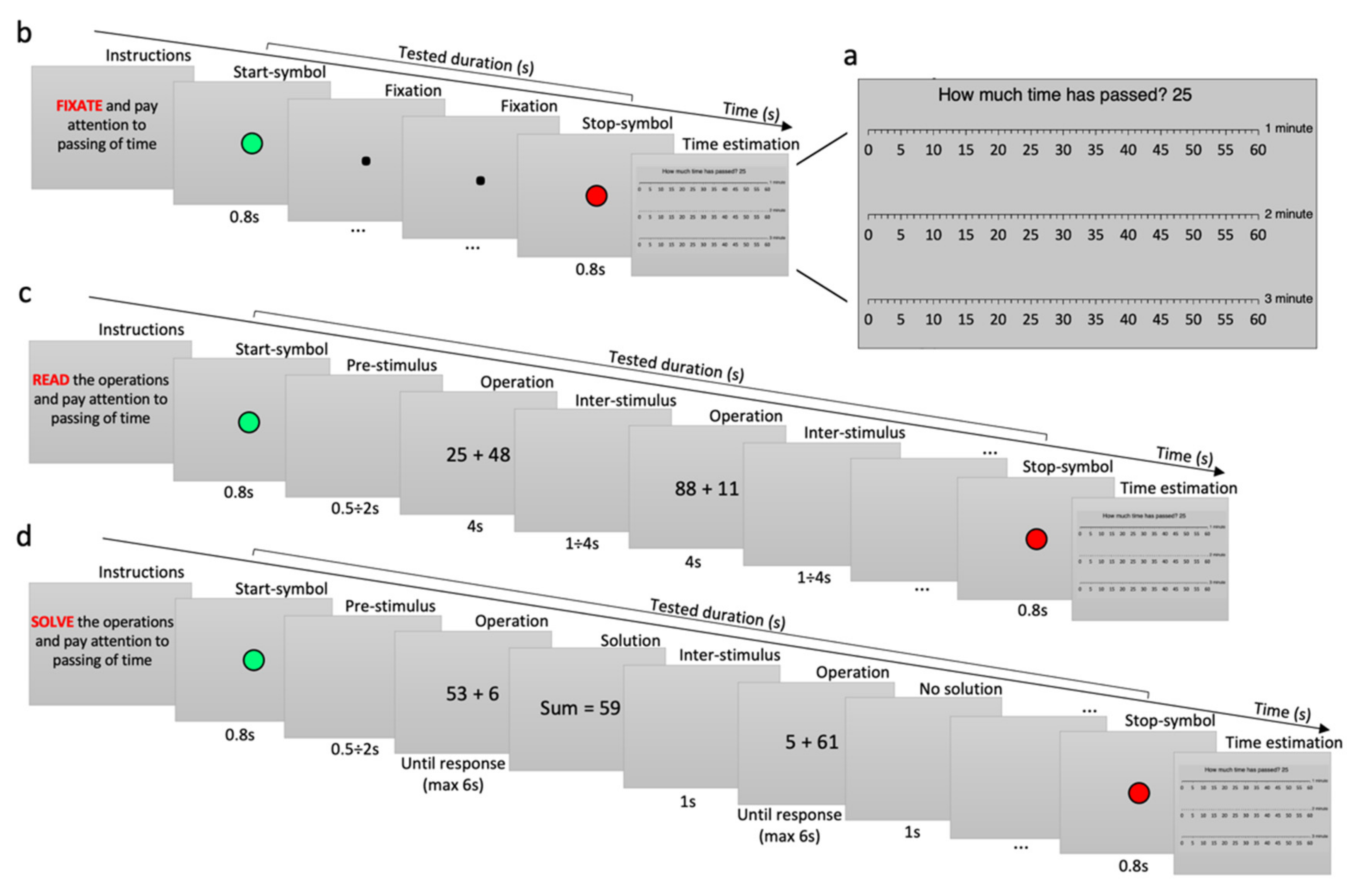
2.3. Procedure and Tasks
2.4. Data Processing and Statistical Analysis
3. Results
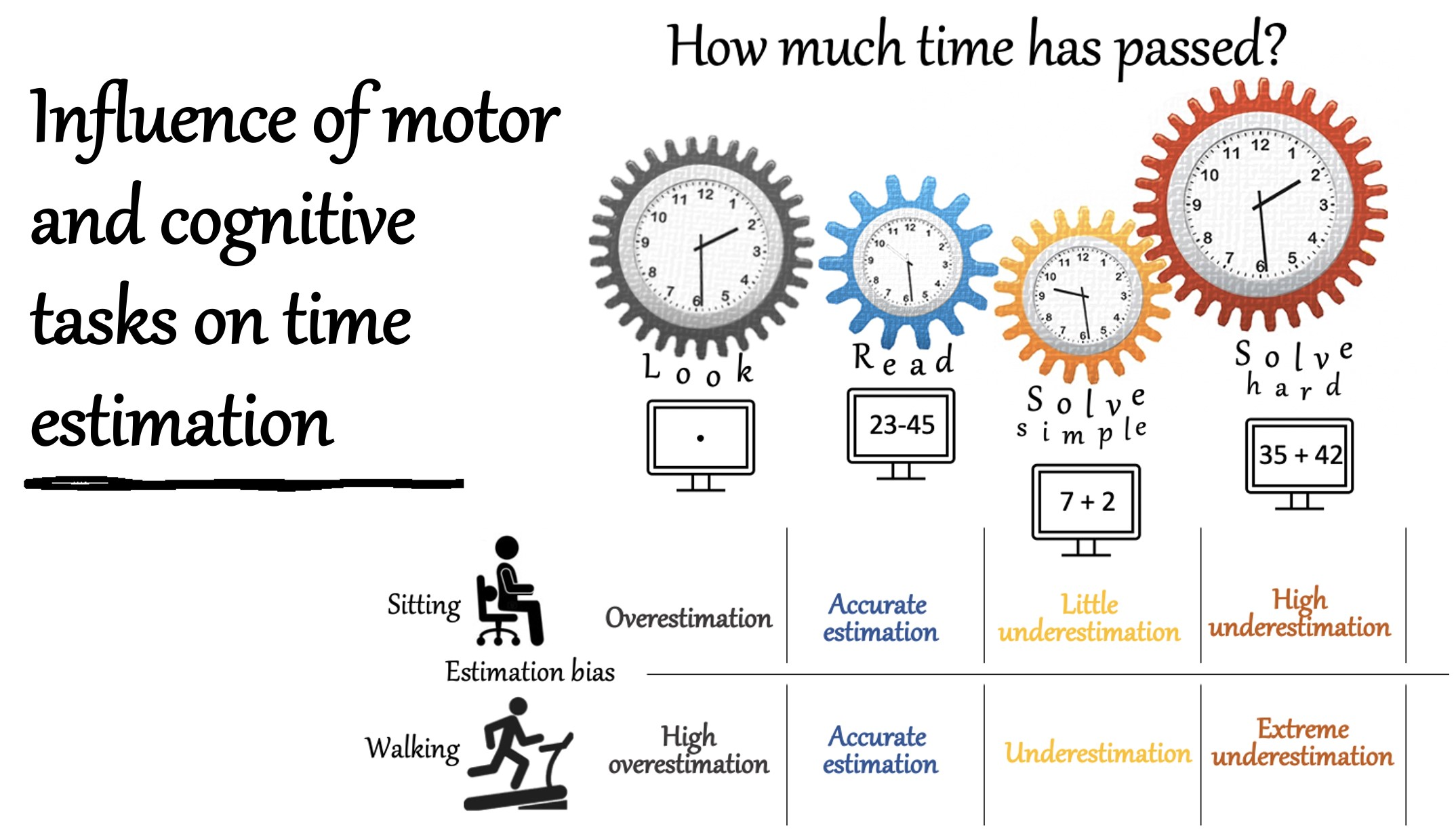
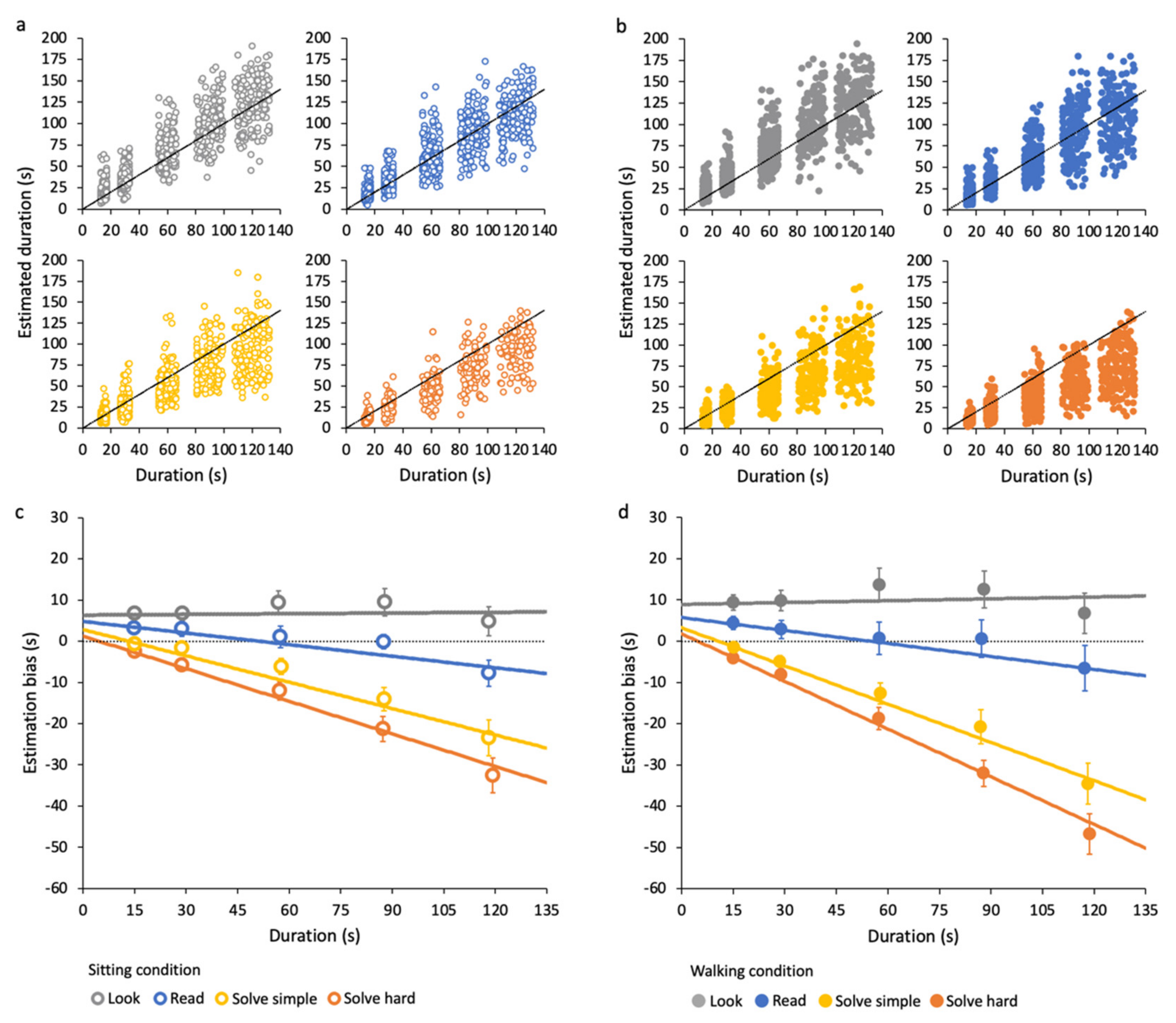
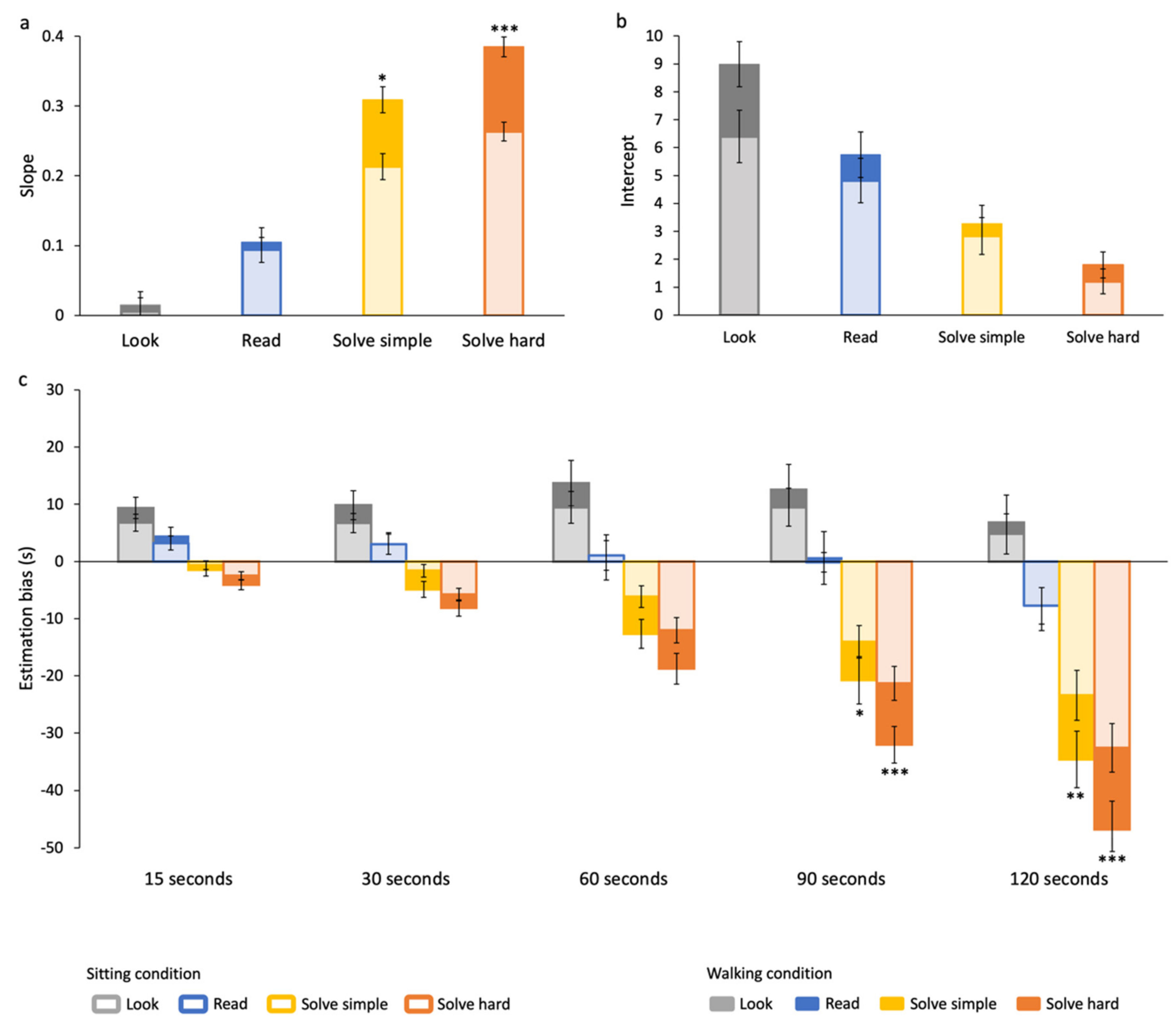
3.1. Effects of Cognitive Task on Time Estimation
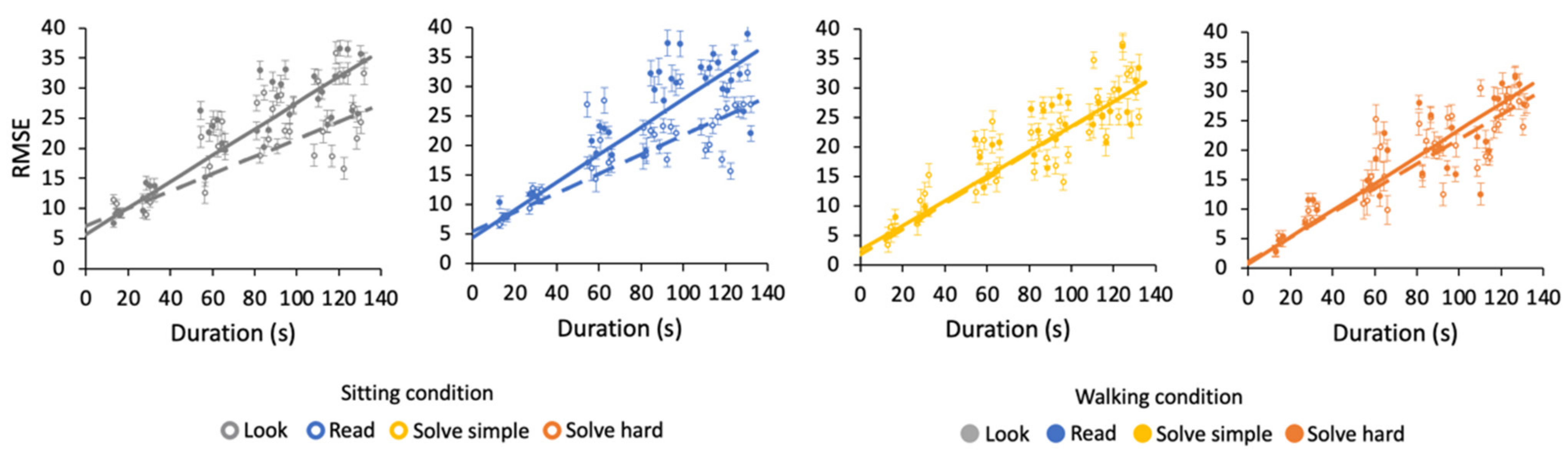
3.2. Effects of Duration on Time Estimation
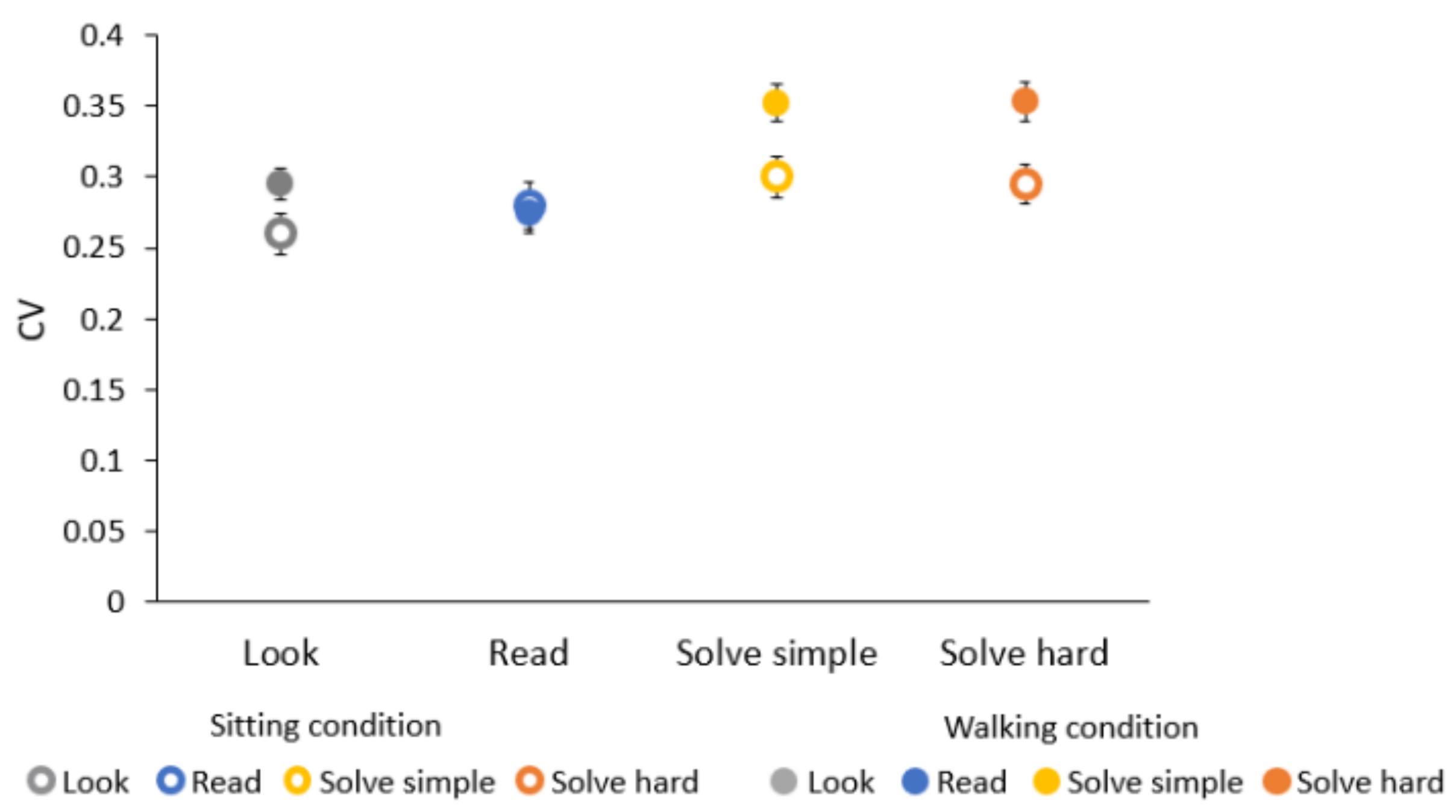
3.3. Effects of Walking on Time Estimation
3.4. Effects of Walking on Cognitive Performance
3.5. Controls of Influencing Variables: Physical Fatigue, Mental Tiredness and Body Temperature
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sucala, M.; Scheckner, B.; David, D. Psychological Time: Interval Length Judgments and Subjective Passage of Time Judgments. Curr. Psychol. Lett. 2011, 26, 2010. [Google Scholar] [CrossRef]
- Gibbon, J. Scalar expectancy theory and Weber’s law in animal timing. Psychol. Rev. 1977, 84, 279–325. [Google Scholar] [CrossRef]
- Buhusi, C.V.; Meck, W. What makes us tick? Functional and neural mechanisms of interval timing. Nat. Rev. Neurosci. 2005, 6, 755–765. [Google Scholar] [CrossRef] [PubMed]
- Wearden, J. “Beyond the fields we know …”: Exploring and developing scalar timing theory. Behav. Process. 1999, 45, 3–21. [Google Scholar] [CrossRef]
- Allman, M.J.; Teki, S.; Griffiths, T.D.; Meck, W.H. Properties of the Internal Clock: First- and Second-Order Principles of Subjective Time. Annu. Rev. Psychol. 2014, 65, 743–771. [Google Scholar] [CrossRef] [Green Version]
- Gibbon, J.; Church, R.M.; Meck, W.H. Scalar Timing in Memory. Ann. N. Y. Acad. Sci. 1984, 423, 52–77. [Google Scholar] [CrossRef]
- Rhodes, D. On the Distinction Between Perceived Duration and Event Timing: Towards a Unified Model of Time Perception. Timing Time Percept. 2018, 6, 90–123. [Google Scholar] [CrossRef] [Green Version]
- Shi, Z.; Church, R.M.; Meck, W.H. Bayesian optimization of time perception. Trends Cogn. Sci. 2013, 17, 556–564. [Google Scholar] [CrossRef]
- Macar, F.; Grondin, S.; Casini, L. Controlled attention sharing influences time estimation. Mem. Cogn. 1994, 22, 673–686. [Google Scholar] [CrossRef] [PubMed]
- Zakay, D.; Block, R.A. The role of attention in time estimation processes. Adv. Psychol. 1996, 115, 143–164. [Google Scholar] [CrossRef]
- Brown, S.W.; West, A.N. Multiple timing and the allocation of attention. Acta Psychol. 1990, 75, 103–121. [Google Scholar] [CrossRef]
- Matthews, W.J.; Meck, W.H. Temporal cognition: Connecting subjective time to perception, attention, and memory. Psychol. Bull. 2016, 142, 865–907. [Google Scholar] [CrossRef] [Green Version]
- Grondin, S. Timing and time perception: A review of recent behavioral and neuroscience findings and theoretical directions. Atten. Percept. Psychophys. 2010, 72, 561–582. [Google Scholar] [CrossRef] [PubMed]
- Matthews, W.J.; Meck, W. Time perception: The bad news and the good. Wiley Interdiscip. Rev. Cogn. Sci. 2014, 5, 429–446. [Google Scholar] [CrossRef] [Green Version]
- Droit-Volet, S.; Fayolle, S.; Lamotte, M.; Gil, S. Time, Emotion and the Embodiment of Timing. Timing Time Percept. 2013, 1, 99–126. [Google Scholar] [CrossRef]
- Lake, J.I. Recent advances in understanding emotion-driven temporal distortions. Curr. Opin. Behav. Sci. 2016, 8, 214–219. [Google Scholar] [CrossRef] [Green Version]
- Wearden, J.H.; Penton-Voak, I.S. Feeling the heat: Body temperature and the rate of subjective time, revisited. Q. J. Exp. Psychol. Sect. B 1995, 48, 129–141. [Google Scholar]
- Van Maanen, L.; Van Der Mijn, R.; Van Beurden, M.H.P.H.; Roijendijk, L.M.M.; Kingma, B.R.M.; Miletić, S.; Van Rijn, H. Core body temperature speeds up temporal processing and choice behavior under deadlines. Sci. Rep. 2019, 9, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Hoagland, H. The Physiological Control of Judgments of Duration: Evidence for a Chemical Clock. J. Gen. Psychol. 1933, 9, 267–287. [Google Scholar] [CrossRef]
- Schwarz, M.; Winkler, I.; Sedlmeier, P. The heart beat does not make us tick: The impacts of heart rate and arousal on time perception. Atten. Percept. Psychophys. 2012, 75, 182–193. [Google Scholar] [CrossRef]
- Ogden, R.S.; Henderson, J.; McGlone, F.; Richter, M. Time distortion under threat: Sympathetic arousal predicts time distortion only in the context of negative, highly arousing stimuli. PLoS ONE 2019, 14, e0216704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Li, H. Interactive effects of trait and state anxieties on time perception. Acta Psychol. 2019, 195, 64–70. [Google Scholar] [CrossRef] [PubMed]
- Lueck, M.D. Anxiety levels: Do they influence the perception of time? J. Undergrad. Res. 2007, 10, 1–4. [Google Scholar]
- Van Hedger, K.; Necka, E.; Barakzai, A.K.; Norman, G.J. The influence of social stress on time perception and psychophysiological reactivity. Psychophysiology 2017, 54, 706–712. [Google Scholar] [CrossRef] [Green Version]
- Soshi, T.; Kuriyama, K.; Aritake, S.; Enomoto, M.; Hida, A.; Tamura, M.; Kim, Y.; Mishima, K. Sleep Deprivation Influences Diurnal Variation of Human Time Perception with Prefrontal Activity Change: A Functional Near-Infrared Spectroscopy Study. PLoS ONE 2010, 5, e8395. [Google Scholar] [CrossRef]
- Tang, N.; Harvey, A.G. Time Estimation Ability and Distorted Perception of Sleep in Insomnia. Behav. Sleep Med. 2005, 3, 134–150. [Google Scholar] [CrossRef]
- Shahabifar, A.; Movahedinia, A. Comparing Time Perception among Morphine-Derived Drugs Addicts and Controls. Addict. Health 2016, 8, 32–40. [Google Scholar]
- Wittmann, M.; Lehnhoff, S. Age Effects in Perception of Time. Psychol. Rep. 2005, 97, 921–935. [Google Scholar] [CrossRef]
- Hanson, N.J.; Buckworth, J. Sex Differences in Time Perception During Self-Paced Running. Med. Sci. Sports Exerc. 2014, 46, 600. [Google Scholar] [CrossRef] [Green Version]
- Behm, D.G.; Carter, T.B. Effect of Exercise-Related Factors on the Perception of Time. Front. Physiol. 2020, 11, 770. [Google Scholar] [CrossRef]
- Plastira, M.N.; Avraamides, M.N. Music Tempo and Perception of Time: Musically Trained vs Nontrained Individuals. Timing Time Percept. 2021, 10, 142–157. [Google Scholar] [CrossRef]
- Cicchini, G.M.; Arrighi, R.; Cecchetti, L.; Giusti, M.; Burr, D. Optimal Encoding of Interval Timing in Expert Percussionists. J. Neurosci. 2012, 32, 1056–1060. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, B.-M.; Meck, W.H. New Perspectives on Vierordt’s Law: Memory-Mixing in Ordinal Temporal Comparison Tasks. In Lecture Notes in Computer Science (Including Subseries Lecture Notes in Artificial Intelligence and Lecture Notes in Bioinformatics); Springer: Heidelberg, Germany, 2011; pp. 67–78. [Google Scholar]
- Jazayeri, M.; Shadlen, M. Temporal context calibrates interval timing. Nat. Neurosci. 2010, 13, 1020–1026. [Google Scholar] [CrossRef] [Green Version]
- Shi, Z.; Burr, D. Predictive coding of multisensory timing. Curr. Opin. Behav. Sci. 2016, 8, 200–206. [Google Scholar] [CrossRef] [Green Version]
- Mamassian, P.; Landy, M.S.; Maloney, L.T. Bayesian Modelling of Visual Perception. In Rao, Olshausen and Lewicki: Probabilistic Models of the Brain: Perception and Neural Function; MIT Press: Cambridge, MA, USA, 2002; pp. 13–36. [Google Scholar]
- Block, R.A.; Hancock, P.A.; Zakay, D. How cognitive load affects duration judgments: A meta-analytic review. Acta Psychol. 2010, 134, 330–343. [Google Scholar] [CrossRef] [PubMed]
- Schiffman, H.R.; Bobko, D.J. The Role of Number and Familiarity of Stimuli in the Perception of Brief Temporal Intervals. Am. J. Psychol. 1977, 90, 85. [Google Scholar] [CrossRef] [PubMed]
- Thomas, E.A.C.; Weaver, W.B. Cognitive processing and time perception. Percept. Psychophys. 1975, 17, 363–367. [Google Scholar] [CrossRef]
- Zakay, D.; Nitzan, D.; Glicksohn, J. The influence of task difficulty and external tempo on subjective time estimation. Percept. Psychophys. 1983, 34, 451–456. [Google Scholar] [CrossRef] [Green Version]
- Brown, S.’I.W. Time perception and attention: The effects of prospective versus retrospective paradigms and task demands on perceived duration. Percept. Psychophys. 1985, 38, 115–124. [Google Scholar] [CrossRef]
- Burnside, W. Judgment of short time intervals while performing mathematical tasks. Percept. Psychophys. 1971, 9, 404–406. [Google Scholar] [CrossRef] [Green Version]
- Gu, B.-M.; van Rijn, H.; Meck, W.H. Oscillatory multiplexing of neural population codes for interval timing and working memory. Neurosci. Biobehav. Rev. 2015, 48, 160–185. [Google Scholar] [CrossRef] [PubMed]
- Brown, S.W. Attentional resources in timing: Interference effects in concurrent temporal and nontemporal working memory tasks. Percept. Psychophys. 1997, 59, 1118–1140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Block, R.A. Psychological timing without a timer: The roles of attention and memory. In Time and Mind II: Information Processing Perspectives; Helfrich, H., Ed.; Hogrefe & Huber Publishers: Cambridge, MA, USA, 2003; pp. 41–59. [Google Scholar]
- Church, R.M.; Broadbent, H.A. Alternative representations of time, number, and rate. Cognition 1990, 37, 55–81. [Google Scholar] [CrossRef]
- Rammsayer, T.; Ulrich, R. No evidence for qualitative differences in the processing of short and long temporal intervals. Acta Psychol. 2005, 120, 141–171. [Google Scholar] [CrossRef] [PubMed]
- Fortin, C.; Champagne, J.; Poirier, M. Temporal order in memory and interval timing: An interference analysis. Acta Psychol. 2007, 126, 18–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Polti, I.; Martin, B.; Van Wassenhove, V. The effect of attention and working memory on the estimation of elapsed time. Sci. Rep. 2018, 8, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Yahya, E.; Dawes, H.; Smith, L.; Dennis, A.; Howells, K.; Cockburn, J. Cognitive motor interference while walking: A systematic review and meta-analysis. Neurosci. Biobehav. Rev. 2011, 35, 715–728. [Google Scholar] [CrossRef]
- Chong, R.K.; Mills, B.; Dailey, L.; Lane, E.; Smith, S.; Lee, K.-H. Specific interference between a cognitive task and sensory organization for stance balance control in healthy young adults: Visuospatial effects. Neuropsychologia 2010, 48, 2709–2718. [Google Scholar] [CrossRef] [PubMed]
- Beauchet, O.; Dubost, V.; Herrmann, F.R.; Kressig, R.W. Stride-to-stride variability while backward counting among healthy young adults. J. Neuroeng. Rehabil. 2005, 2, 26. [Google Scholar] [CrossRef] [PubMed]
- Tomassini, A.; Morrone, M.C. Perceived visual time depends on motor preparation and direction of hand movements. Sci. Rep. 2016, 6, 27947. [Google Scholar] [CrossRef] [Green Version]
- Press, C.; Berlot, E.; Bird, G.; Ivry, R.; Cook, R. Moving time: The influence of action on duration perception. J. Exp. Psychol. Gen. 2014, 143, 1787–1793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yon, D.; Edey, R.; Ivry, R.B.; Press, C. Time on your hands: Perceived duration of sensory events is biased toward concurrent actions. J. Exp. Psychol. Gen. 2017, 146, 182–193. [Google Scholar] [CrossRef] [Green Version]
- Yokosaka, T.; Kuroki, S.; Nishida, S.; Watanabe, J. Apparent Time Interval of Visual Stimuli Is Compressed during Fast Hand Movement. PLoS ONE 2015, 10, e0124901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sayalı, C.; Uslu, E.; Menceloğlu, M.; Canbeyli, R.; Balcı, F. Effect of Acute Physical Activity on Interval Timing. Timing Time Percept 2018, 6, 14–31. [Google Scholar] [CrossRef]
- Kroger-Costa, A.; Machado, A.; Santos, J. Effects of motion on time perception. Behav. Process. 2013, 95, 50–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ivry, R.B.; Schlerf, J.E. Dedicated and intrinsic models of time perception. Trends Cogn. Sci. 2008, 12, 273–280. [Google Scholar] [CrossRef] [Green Version]
- Wittmann, M. The inner experience of time. Philos. Trans. R. Soc. B Biol. Sci. 2009, 364, 1955–1967. [Google Scholar] [CrossRef]
- Grondin, S.; Plourde, M. Judging Multi-Minute Intervals Retrospectively. Q. J. Exp. Psychol. 2007, 60, 1303–1312. [Google Scholar] [CrossRef]
- Zakay, D. Time Estimation Methods—Do They Influence Prospective Duration Estimates? Perception 1993, 22, 91–101. [Google Scholar] [CrossRef]
- Oberfeld, D.; Thãnes, S.; Palayoor, B.J.; Hecht, H.; Thönes, S. Depression does not affect time perception and time-to-contact estimation. Front. Psychol. 2014, 5, 810. [Google Scholar] [CrossRef] [Green Version]
- Kleiner, M.; Brainard, D.H.; Pelli, D.G. What’s new in Psychtoolbox-3? Perception 2007, 36, 1–16. [Google Scholar]
- Pelli, D.G. The VideoToolbox software for visual psychophysics: Transforming numbers into movies. Spat. Vis. 1997, 10, 437–442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brainard, D.H. The Psychophysics Toolbox. Spat. Vis. 1997, 10, 433–436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hodges, J.L.; Lehmann, E.L. Estimates of Location Based on Rank Tests. Ann. Math. Stat. 1963, 34, 598–611. [Google Scholar] [CrossRef]
- Molet, M.; Alessandri, J.; Zentall, T.R. Subjective Time: Cognitive and Physical Secondary Tasks Affect Timing Differently. Q. J. Exp. Psychol. 2011, 64, 1344–1353. [Google Scholar] [CrossRef]
- Merchant, H.; Harrington, D.L.; Meck, W.H. Neural Basis of the Perception and Estimation of Time. Annu. Rev. Neurosci. 2013, 36, 313–336. [Google Scholar] [CrossRef]
- Merchant, H.; Yarrow, K. How the motor system both encodes and influences our sense of time. Curr. Opin. Behav. Sci. 2016, 8, 22–27. [Google Scholar] [CrossRef] [Green Version]
- Coull, J.T.; Cheng, R.-K.; Meck, W. Neuroanatomical and Neurochemical Substrates of Timing. Neuropsychopharmacology 2010, 36, 3–25. [Google Scholar] [CrossRef] [Green Version]
- Leisman, G.; Moustafa, A.; Shafir, T. Thinking, Walking, Talking: Integratory Motor and Cognitive Brain Function. Front. Public Heal. 2016, 4, 94. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.S.; Chang, T.-T.; Majid, A.; Caspers, S.; Eickhoff, S.B.; Menon, V. Functional Heterogeneity of Inferior Parietal Cortex during Mathematical Cognition Assessed with Cytoarchitectonic Probability Maps. Cereb. Cortex 2009, 19, 2930–2945. [Google Scholar] [CrossRef] [Green Version]
- Hayashi, M.J.; van der Zwaag, W.; Bueti, D.; Kanai, R. Representations of time in human frontoparietal cortex. Commun. Biol. 2018, 1, 233. [Google Scholar] [CrossRef] [PubMed]
- Paul, S.S.; Ada, L.; Canning, C.G. Automaticity of walking—Implications for physiotherapy practice. Phys. Ther. Rev. 2005, 10, 15–23. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sitting | 15 s | 30 s | 60 s | 90 s | 120 s |
|---|---|---|---|---|---|
| Look | M = 21.8, SE = 1.5 | M = 35.6, SE = 1.7 | M = 66.3, SE = 2.8 | M = 97.3, SE = 3.2 | M = 122.9, SE = 3.4 |
| Read | M = 18.1, SE = 1.2 | M = 31.8, SE = 1.7 | M = 58.5, SE = 2.4 | M = 87.3, SE = 1.7 | M = 110.3, SE = 3.4 |
| Solve simple | M = 14.4, SE = 0.7 | M = 27.1, SE = 1.1 | M = 51.5, SE = 1.7 | M = 73.6, SE = 2.8 | M = 94.7, SE = 4.3 |
| Solve hard | M = 12.7, SE = 0.7 | M = 23.1, SE = 1.1 | M = 45.2, SE = 2.2 | M = 65.9, SE = 2.9 | M = 86.7, SE = 3.9 |
| Walking | 15 s | 30 s | 60 s | 90 s | 120 s |
| Look | M = 24.4, SE = 1.8 | M = 38.7, SE = 2.4 | M = 71.2, SE = 3.8 | M = 100.6, SE = 4.5 | M = 124, SE = 4.7 |
| Read | M = 19.4, SE = 1.5 | M = 31.8, SE = 1.9 | M = 58.2, SE = 3.9 | M = 87.8, SE = 4.5 | M = 110.8, SE = 5.4 |
| Solve simple | M = 13.6, SE = 1.1 | M = 23.7, SE = 1.3 | M = 45.1, SE = 2.5 | M = 66.2, SE = 4.1 | M = 83.7, SE = 4.7 |
| Solve hard | M = 10.9, SE = 0.8 | M = 20.8, SE = 1.4 | M = 38.6, SE = 2.6 | M = 55.9, SE = 3.1 | M = 71.9, SE = 4.6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Castellotti, S.; D’Agostino, O.; Biondi, A.; Pignatiello, L.; Del Viva, M.M. Influence of Motor and Cognitive Tasks on Time Estimation. Brain Sci. 2022, 12, 404. https://doi.org/10.3390/brainsci12030404
Castellotti S, D’Agostino O, Biondi A, Pignatiello L, Del Viva MM. Influence of Motor and Cognitive Tasks on Time Estimation. Brain Sciences. 2022; 12(3):404. https://doi.org/10.3390/brainsci12030404
Chicago/Turabian StyleCastellotti, Serena, Ottavia D’Agostino, Alessandra Biondi, Luigi Pignatiello, and Maria Michela Del Viva. 2022. "Influence of Motor and Cognitive Tasks on Time Estimation" Brain Sciences 12, no. 3: 404. https://doi.org/10.3390/brainsci12030404