
Interoceptive Attentiveness Induces Significantly More PFC Activation during a Synchronized Linguistic Task Compared to a Motor Task as Revealed by Functional Near-Infrared Spectroscopy


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample
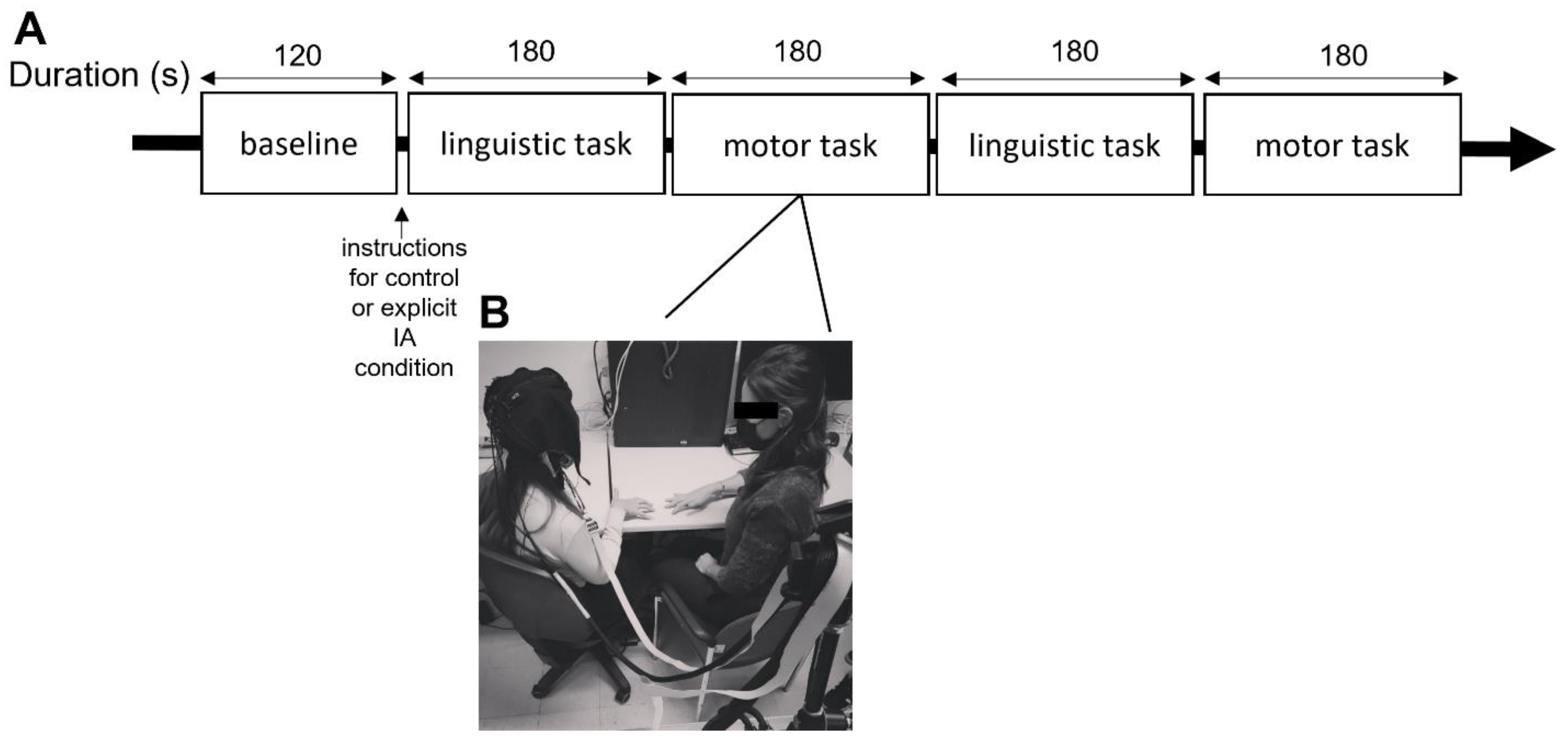
2.2. Experimental Procedure
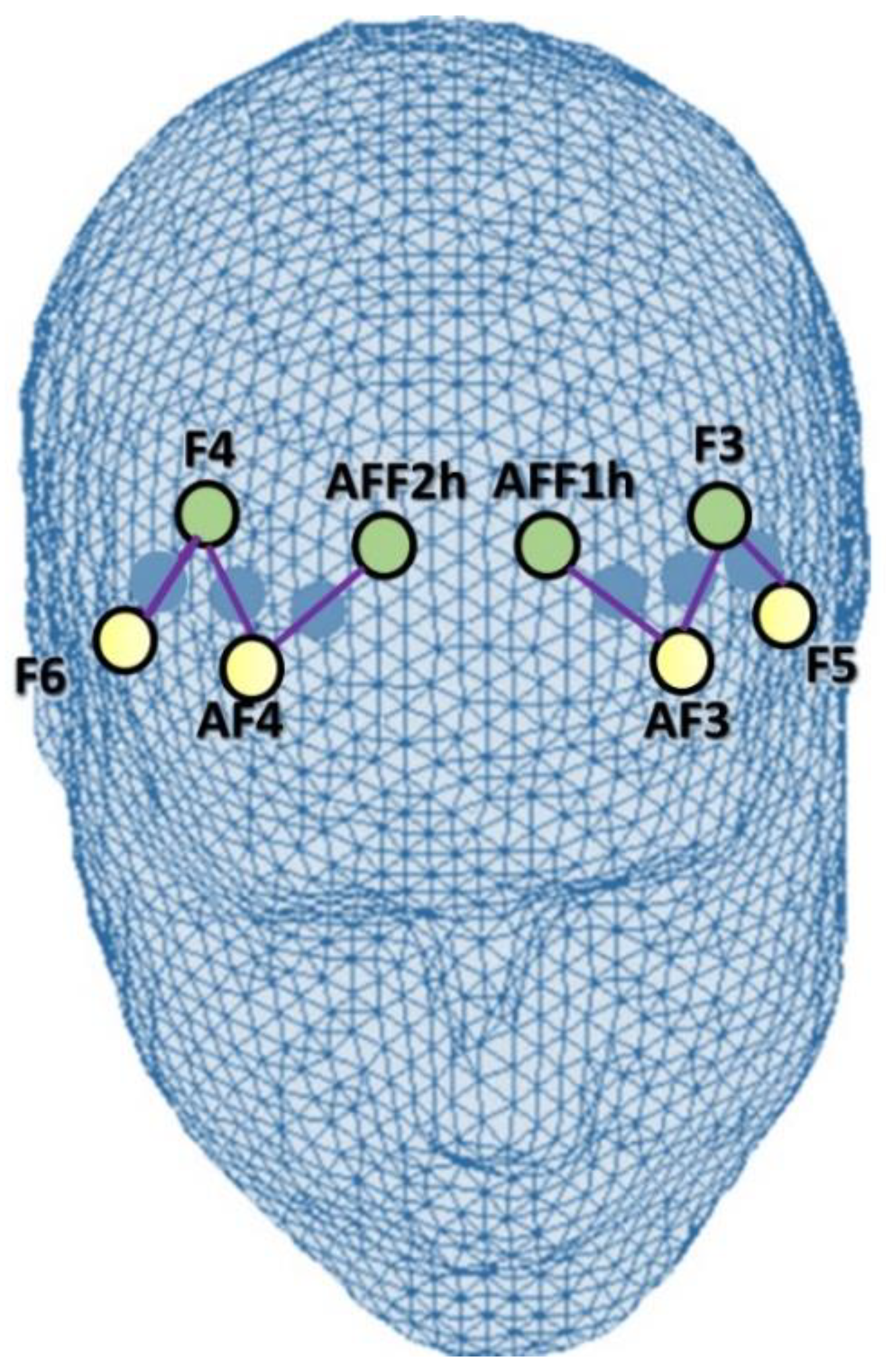
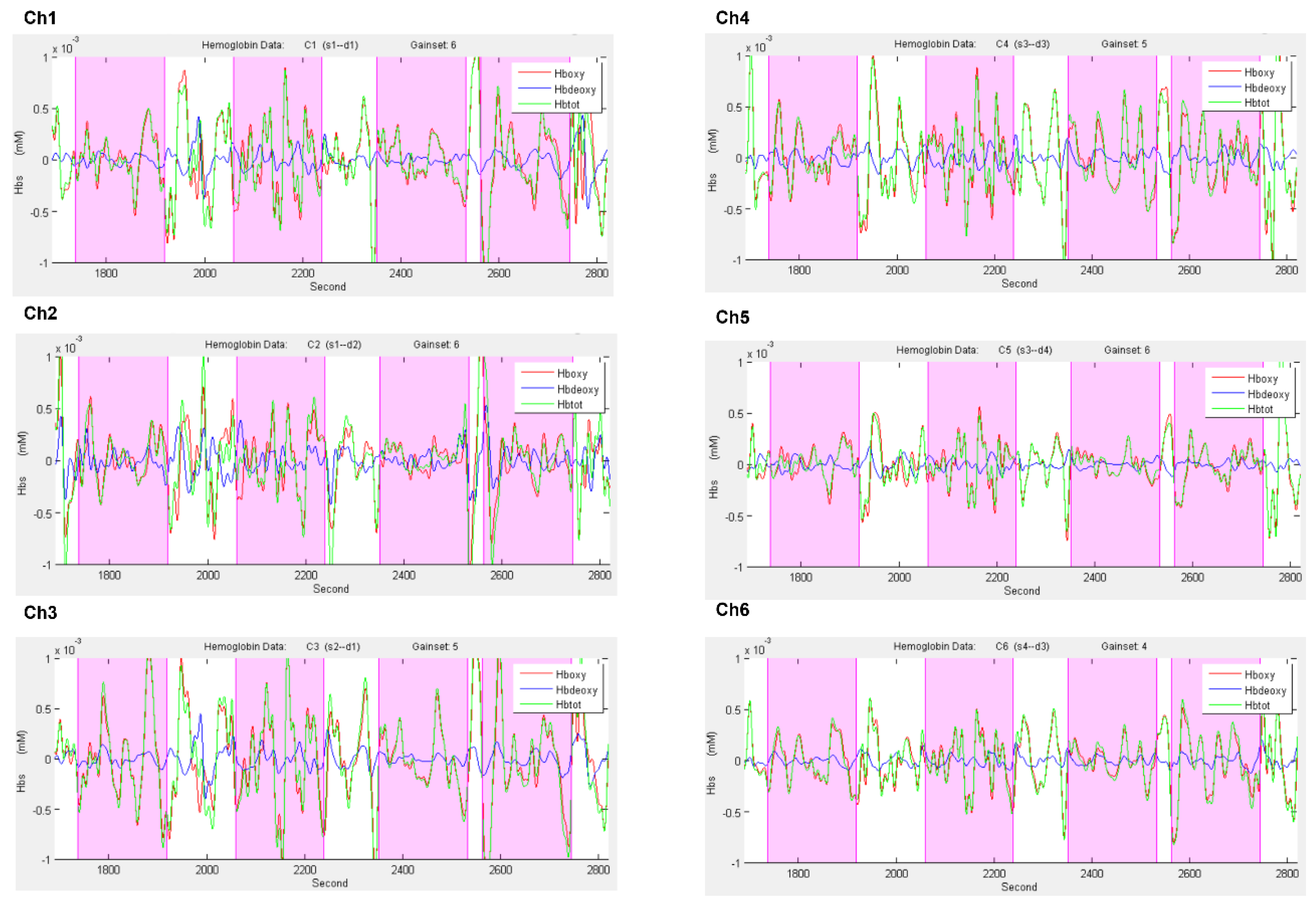
2.3. fNIRS Data Acquisition and Biosignal Analysis
2.4. Statistical Analysis
3. Results
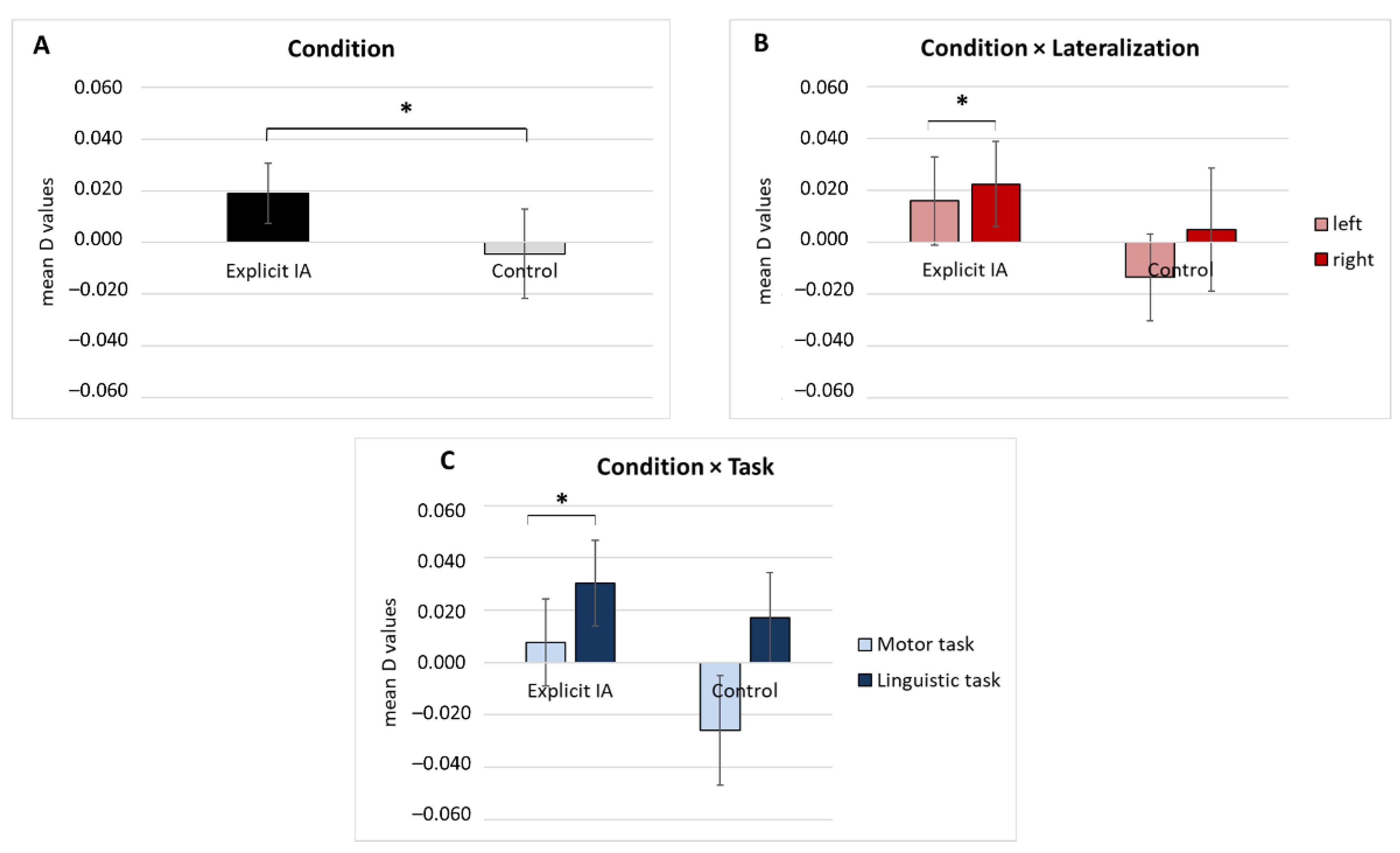
fNIRS Results Subsection
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schulz, S.M. Neural correlates of heart-focused interoception: A functional magnetic resonance imaging meta-analysis. Philos. Trans. R. Soc. B Biol. Sci. 2016, 371, 20160018. [Google Scholar] [CrossRef]
- Tsakiris, M.; De Preester, H. The Interoceptive Mind: From Homeostasis to Awareness; Oxford University Press: Oxford, UK, 2018. [Google Scholar]
- Farb, N.A.S.; Segal, Z.V.; Anderson, A.K. Mindfulness meditation training alters cortical representations of interoceptive attention. Soc. Cogn. Affect. Neurosci. 2013, 8, 15–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weng, H.Y.; Feldman, J.L.; Leggio, L.; Napadow, V.; Park, J.; Price, C.J. Interventions and Manipulations of Interoception. Trends Neurosci. 2021, 44, 52–62. [Google Scholar] [CrossRef]
- Arch, J.J.; Craske, M.G. Mechanisms of mindfulness: Emotion regulation following a focused breathing induction. Behav. Res. Ther. 2006, 44, 1849–1858. [Google Scholar] [CrossRef]
- Varga, S.; Heck, D.H. Rhythms of the body, rhythms of the brain: Respiration, neural oscillations, and embodied cognition. Conscious. Cogn. 2017, 56, 77–90. [Google Scholar] [CrossRef] [PubMed]
- Paul, G.; Elam, B.; Verhulst, S.J. A longitudinal study of students’ perceptions of using deep breathing meditation to reduce testing stresses. Teach. Learn. Med. 2007, 19, 287–292. [Google Scholar] [CrossRef] [PubMed]
- Balconi, M.; Angioletti, L. One’s Interoception Affects the Representation of Seeing Others’ Pain: A Randomized Controlled qEEG Study. Pain Res. Manag. 2021, 2021, 5585060. [Google Scholar] [CrossRef]
- Grossman, P. Defining Mindfulness by How Poorly I Think I Pay Attention During Everyday Awareness and Other Intractable Problems for Psychology’s (Re)Invention of Mindfulness: Comment on Brown et al. Psychol. Assess. 2011, 23, 1034–1040. [Google Scholar] [CrossRef]
- Kashkouli Nejad, K.; Sugiura, M.; Nozawa, T.; Kotozaki, Y.; Furusawa, Y.; Nishino, K.; Nukiwa, T.; Kawashima, R. Supramarginal activity in interoceptive attention tasks. Neurosci. Lett. 2015, 589, 42–46. [Google Scholar] [CrossRef] [Green Version]
- Tang, Y.-Y.; Ma, Y.; Fan, Y.; Feng, H.; Wang, J.; Feng, S.; Lu, Q.; Hu, B.; Lin, Y.; Li, J.; et al. Central and autonomic nervous system interaction is altered by short-term meditation. Proc. Natl. Acad. Sci. USA 2009, 106, 8865–8870. [Google Scholar] [CrossRef] [Green Version]
- Dickenson, J.; Berkman, E.T.; Arch, J.; Lieberman, M.D. Neural correlates of focused attention during a brief mindfulness induction. Soc. Cogn. Affect. Neurosci. 2013, 8, 40–47. [Google Scholar] [CrossRef] [PubMed]
- Farb, N.A.S.; Segal, Z.V.; Mayberg, H.; Bean, J.; Mckeon, D.; Fatima, Z.; Anderson, A.K. Attending to the present: Mindfulness meditation reveals distinct neural modes of self-reference. Soc. Cogn. Affect. Neurosci. 2007, 2, 313–322. [Google Scholar] [CrossRef] [PubMed]
- Kilpatrick, L.A.; Suyenobu, B.Y.; Smith, S.R.; Bueller, J.A.; Goodman, T.; Creswell, J.D.; Tillisch, K.; Mayer, E.A.; Naliboff, B.D. Impact of mindfulness-based stress reduction training on intrinsic brain connectivity. Neuroimage 2011, 56, 290–298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, Y.L.; Wang, D.X.; Zhang, Y.R.; Tang, Y.Y. Enhancing Attention by Synchronizing Respiration and Fingertip Pressure: A Pilot Study Using Functional Near-Infrared Spectroscopy. Front. Neurosci. 2019, 13, 1209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balconi, M.; Angioletti, L. Interoception as a social alarm amplification system. What multimethod (EEG-fNIRS) integrated measures can tell us about interoception and empathy for pain? Neuropsychol. Trends 2021, 29, 39–64. [Google Scholar] [CrossRef]
- Zhang, Z.; Olszewska-Guizzo, A.; Husain, S.F.; Bose, J.; Choi, J.; Tan, W.; Wang, J.; Tran, B.X.; Wang, B.; Jin, Y.; et al. Brief relaxation practice induces significantly more prefrontal cortex activation during arithmetic tasks comparing to viewing greenery images as revealed by functional near-infrared spectroscopy (fNIRS). Int. J. Environ. Res. Public Health 2020, 17, 8366. [Google Scholar] [CrossRef]
- Kondo, H.; Osaka, N.; Osaka, M. Cooperation of the anterior cingulate cortex and dorsolateral prefrontal cortex for attention shifting. Neuroimage 2004, 23, 670–679. [Google Scholar] [CrossRef]
- Boorman, E.D.; Behrens, T.E.J.; Woolrich, M.W.; Rushworth, M.F.S. How Green Is the Grass on the Other Side? Frontopolar Cortex and the Evidence in Favor of Alternative Courses of Action. Neuron 2009, 62, 733–743. [Google Scholar] [CrossRef] [Green Version]
- Pollmann, S. Frontopolar Resource Allocation in Human and Nonhuman Primates. Trends Cogn. Sci. 2016, 20, 84–86. [Google Scholar] [CrossRef]
- Zajkowski, W.; Kossut, M.; Wilson, R.C. A causal role for right frontopolar cortex in directed, but not random, exploration. eLife 2017, 6, e27430. [Google Scholar] [CrossRef] [Green Version]
- Cheng, X.; Li, X.; Hu, Y. Synchronous brain activity during cooperative exchange depends on gender of partner: A fNIRS-based hyperscanning study. Hum. Brain Mapp. 2015, 36, 2039–2048. [Google Scholar] [CrossRef] [PubMed]
- Amodio, D.M.; Frith, C.D. Meeting of minds: The medial frontal cortex and social cognition. Nat. Rev. Neurosci. 2006, 7, 268–277. [Google Scholar] [CrossRef] [PubMed]
- Balconi, M.; Pezard, L.; Nandrino, J.-L.; Vanutelli, M.E. Two is better than one: The effects of strategic cooperation on intra- and inter-brain connectivity by fNIRS. PLoS ONE 2017, 12, e0187652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, Y.; Wang, Z.; Song, B.; Pan, Y.; Cheng, X.; Zhu, Y.; Hu, Y. How to calculate and validate inter-brain synchronization in a fnirs hyperscanning study. J. Vis. Exp. 2021, 175, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Balconi, M.; Vanutelli, M.E. Cooperation and competition with hyperscanning methods: Review and future application to emotion domain. Front. Comput. Neurosci. 2017, 11, 86. [Google Scholar] [CrossRef]
- Hove, M.J.; Risen, J.L. It’s all in the timing: Interpersonal synchrony increases affiliation. Soc. Cogn. 2009, 27, 949–960. [Google Scholar] [CrossRef] [Green Version]
- Valdesolo, P.; Ouyang, J.; DeSteno, D. The rhythm of joint action: Synchrony promotes cooperative ability. J. Exp. Soc. Psychol. 2010, 46, 693–695. [Google Scholar] [CrossRef]
- Valdesolo, P.; DeSteno, D. Synchrony and the social tuning of compassion. Emotion 2011, 11, 262–266. [Google Scholar] [CrossRef]
- Reddish, P.; Tong, E.M.W.; Jong, J.; Lanman, J.A.; Whitehouse, H. Collective synchrony increases prosociality towards non-performers and outgroup members. Br. J. Soc. Psychol. 2016, 55, 722–738. [Google Scholar] [CrossRef] [Green Version]
- Montague, P.R.; Berns, G.S.; Cohen, J.D.; McClure, S.M.; Pagnoni, G.; Dhamala, M.; Wiest, M.C.; Karpov, I.; King, R.D.; Apple, N.; et al. Hyperscanning: Simultaneous fMRI during linked social interactions. Neuroimage 2002, 16, 1159–1164. [Google Scholar] [CrossRef]
- Funane, T.; Kiguchi, M.; Atsumori, H.; Sato, H.; Kubota, K.; Koizumi, H. Synchronous activity of two people’s prefrontal cortices during a cooperative task measured by simultaneous near-infrared spectroscopy. J. Biomed. Opt. 2011, 16, 077011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, X.; Bryant, D.M.; Reiss, A.L. NIRS-based hyperscanning reveals increased interpersonal coherence in superior frontal cortex during cooperation. Neuroimage 2012, 59, 2430–2437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holper, L.; Scholkmann, F.; Wolf, M. Between-brain connectivity during imitation measured by fNIRS. Neuroimage 2012, 63, 212–222. [Google Scholar] [CrossRef] [PubMed]
- Baker, J.M.; Liu, N.; Cui, X.; Vrticka, P.; Saggar, M.; Hosseini, S.M.H.; Reiss, A.L. Sex differences in neural and behavioral signatures of cooperation revealed by fNIRS hyperscanning. Sci. Rep. 2016, 6, 26492. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.; Cheng, X.; Zhang, Z.; Li, X.; Hu, Y. Cooperation in lovers: An fNIRS-based hyperscanning study. Hum. Brain Mapp. 2017, 38, 831–841. [Google Scholar] [CrossRef]
- Jiang, J.; Dai, B.; Peng, D.; Zhu, C.; Liu, L.; Lu, C. Neural synchronization during face-to-face communication. J. Neurosci. 2012, 32, 16064–16069. [Google Scholar] [CrossRef]
- Scholkmann, F.; Holper, L.; Wolf, U.; Wolf, M. A new methodical approach in neuroscience: Assessing inter-personal brain coupling using functional near-infrared imaging (fNIRI) hyperscanning. Front. Hum. Neurosci. 2013, 7, 813. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.; Saito, G.; Lin, C.; Saito, H. Inter-brain network underlying turn-based cooperation and competition: A hyperscanning study using near-infrared spectroscopy. Sci. Rep. 2017, 7, 8684. [Google Scholar] [CrossRef]
- Zhang, Y.; Meng, T.; Hou, Y.; Pan, Y.; Hu, Y. Interpersonal brain synchronization associated with working alliance during psychological counseling. Psychiatry Res. Neuroimaging 2018, 282, 103–109. [Google Scholar] [CrossRef]
- Descorbeth, O.; Zhang, X.; Adam Noah, J.; Hirsch, J. Neural processes for live pro-social dialogue between dyads with socioeconomic disparity. Soc. Cogn. Affect. Neurosci. 2020, 15, 875–887. [Google Scholar] [CrossRef]
- Hirsch, J.; Noah, J.A.; Zhang, X.; Dravida, S.; Ono, Y. A cross-brain neural mechanism for human-to-human verbal communication. Soc. Cogn. Affect. Neurosci. 2018, 13, 907–920. [Google Scholar] [CrossRef] [Green Version]
- Kelsen, B.A.; Sumich, A.; Kasabov, N.; Liang, S.H.Y.; Wang, G.Y. What has social neuroscience learned from hyperscanning studies of spoken communication? A systematic review. Neurosci. Biobehav. Rev. 2020, 132, 1249–1262. [Google Scholar] [CrossRef] [PubMed]
- Xue, H.; Lu, K.; Hao, N. Cooperation makes two less-creative individuals turn into a highly-creative pair. Neuroimage 2018, 172, 527–537. [Google Scholar] [CrossRef]
- Uddin, L.Q.; Supekar, K.; Menon, V. Typical and atypical development of functional human brain networks: Insights from resting-state fMRI. Front. Syst. Neurosci. 2010, 4, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhat, A.N.; Hoffman, M.D.; Trost, S.L.; Culotta, M.L.; Eilbott, J.; Tsuzuki, D.; Pelphrey, K.A. Cortical activation during action observation, action execution, and interpersonal synchrony in adults: A functional near-infrared spectroscopy (fNIRS) study. Front. Hum. Neurosci. 2017, 11, 431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balconi, M.; Molteni, E. Past and future of near-infrared spectroscopy in studies of emotion and social neuroscience. J. Cogn. Psychol. 2016, 28, 129–146. [Google Scholar] [CrossRef]
- Quaresima, V.; Ferrari, M. Functional Near-Infrared Spectroscopy (fNIRS) for Assessing Cerebral Cortex Function During Human Behavior in Natural/Social Situations: A Concise Review. Organ. Res. Methods 2019, 22, 46–68. [Google Scholar] [CrossRef]
- Angioletti, L.; Vanutelli, M.E.; Fronda, G.; Balconi, M. Exploring the Connected Brain by fNIRS: Human-to-Human Interactions Engineering. Appl. Mech. Mater. 2019, 893, 13–19. [Google Scholar] [CrossRef]
- Arnold, A.J.; Winkielman, P.; Dobkins, K. Interoception and Social Connection. Front. Psychol. 2019, 10, 2589. [Google Scholar] [CrossRef] [Green Version]
- Khoe, H.C.H.; Low, J.W.; Wijerathne, S.; Ann, L.S.; Salgaonkar, H.; Lomanto, D.; Choi, J.K.; Baek, J.Y.; Tam, W.W.; Pei, H.; et al. Use of prefrontal cortex activity as a measure of learning curve in surgical novices: Results of a single blind randomised controlled trial. Surg. Endosc. 2020, 34, 5604–5615. [Google Scholar] [CrossRef]
- Sänger, J.; Lindenberger, U.; Müller, V. Interactive brains, social minds. Commun. Integr. Biol. 2011, 4, 655–663. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Pelowski, M. Clarifying the interaction types in two-person neuroscience research. Front. Hum. Neurosci. 2014, 8, 276. [Google Scholar] [CrossRef] [PubMed]
- Faul, F.; Erdfelder, E.; Lang, A.G.; Buchner, A. G*Power 3: A flexible statistical power analysis program for the social, behavioral, and biomedical sciences. J. Mater. Environ. Sci. 2007, 39, 175–191. [Google Scholar] [CrossRef] [PubMed]
- Kawasaki, M.; Yamada, Y.; Ushiku, Y.; Miyauchi, E.; Yamaguchi, Y. Inter-brain synchronization during coordination of speech rhythm in human-to-human social interaction. Sci. Rep. 2013, 3, 1692. [Google Scholar] [CrossRef] [Green Version]
- Jobbágy, Á.; Harcos, P.; Karoly, R.; Fazekas, G. Analysis of finger-tapping movement. J. Neurosci. Methods 2005, 141, 29–39. [Google Scholar] [CrossRef]
- Oostenveld, R.; Praamstra, P. The five percent electrode system for high-resolution EEG and ERP measurements. Clin. Neurophysiol. 2001, 112, 713–719. [Google Scholar] [CrossRef]
- Balconi, M.; Vanutelli, M.E. Empathy in negative and positive interpersonal interactions. What is the relationship between central (EEG, fNIRS) and peripheral (autonomic) neurophysiological responses? Adv. Cogn. Psychol. 2017, 13, 105–120. [Google Scholar] [CrossRef]
- Giacometti, P.; Perdue, K.L.; Diamond, S.G. Algorithm to find high density EEG scalp coordinates and analysis of their correspondence to structural and functional regions of the brain. J. Neurosci. Methods 2014, 229, 84–96. [Google Scholar] [CrossRef] [Green Version]
- Koessler, L.; Maillard, L.; Benhadid, A.; Vignal, J.P.; Felblinger, J.; Vespignani, H.; Braun, M. Automated cortical projection of EEG sensors: Anatomical correlation via the international 10-10 system. Neuroimage 2009, 46, 64–72. [Google Scholar] [CrossRef]
- Pinti, P.; Aichelburg, C.; Lind, F.; Power, S.; Swingler, E.; Merla, A.; Hamilton, A.; Gilber, S.; Burgess, P.; Tachtsidis, I. Using fiberless, wearable fnirs to monitor brain activity in real-world cognitive tasks. J. Vis. Exp. 2015, e53336. [Google Scholar] [CrossRef] [Green Version]
- Naseer, N.; Hong, K.S. Classification of functional near-infrared spectroscopy signals corresponding to the right- and left-wrist motor imagery for development of a brain-computer interface. Neurosci. Lett. 2013, 553, 84–89. [Google Scholar] [CrossRef] [PubMed]
- Naseer, N.; Hong, M.J.; Hong, K.S. Online binary decision decoding using functional near-infrared spectroscopy for the development of brain-computer interface. Exp. Brain Res. 2014, 232, 555–564. [Google Scholar] [CrossRef] [PubMed]
- Balconi, M.; Vanutelli, M.E. Interbrains cooperation: Hyperscanning and self-perception in joint actions. J. Clin. Exp. Neuropsychol. 2017, 39, 607–620. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Balconi, M.; Angioletti, L. Interoceptive Attentiveness Induces Significantly More PFC Activation during a Synchronized Linguistic Task Compared to a Motor Task as Revealed by Functional Near-Infrared Spectroscopy. Brain Sci. 2022, 12, 301. https://doi.org/10.3390/brainsci12030301
Balconi M, Angioletti L. Interoceptive Attentiveness Induces Significantly More PFC Activation during a Synchronized Linguistic Task Compared to a Motor Task as Revealed by Functional Near-Infrared Spectroscopy. Brain Sciences. 2022; 12(3):301. https://doi.org/10.3390/brainsci12030301
Chicago/Turabian StyleBalconi, Michela, and Laura Angioletti. 2022. "Interoceptive Attentiveness Induces Significantly More PFC Activation during a Synchronized Linguistic Task Compared to a Motor Task as Revealed by Functional Near-Infrared Spectroscopy" Brain Sciences 12, no. 3: 301. https://doi.org/10.3390/brainsci12030301