
Load-Dependent Prefrontal Cortex Activation Assessed by Continuous-Wave Near-Infrared Spectroscopy during Two Executive Tasks with Three Cognitive Loads in Young Adults

Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Cognitive Assessment
2.2.1. n-Back Task
2.2.2. Random Number Generation Task
2.3. Apparatus
2.4. fNIRS Data Analysis
2.5. Procedure
2.6. Statistical Analysis
3. Results
3.1. Subjective Perceived Difficulty
3.2. Behavioral Performances
3.2.1. n-Back
3.2.2. RNG
3.3. fNIRS Data
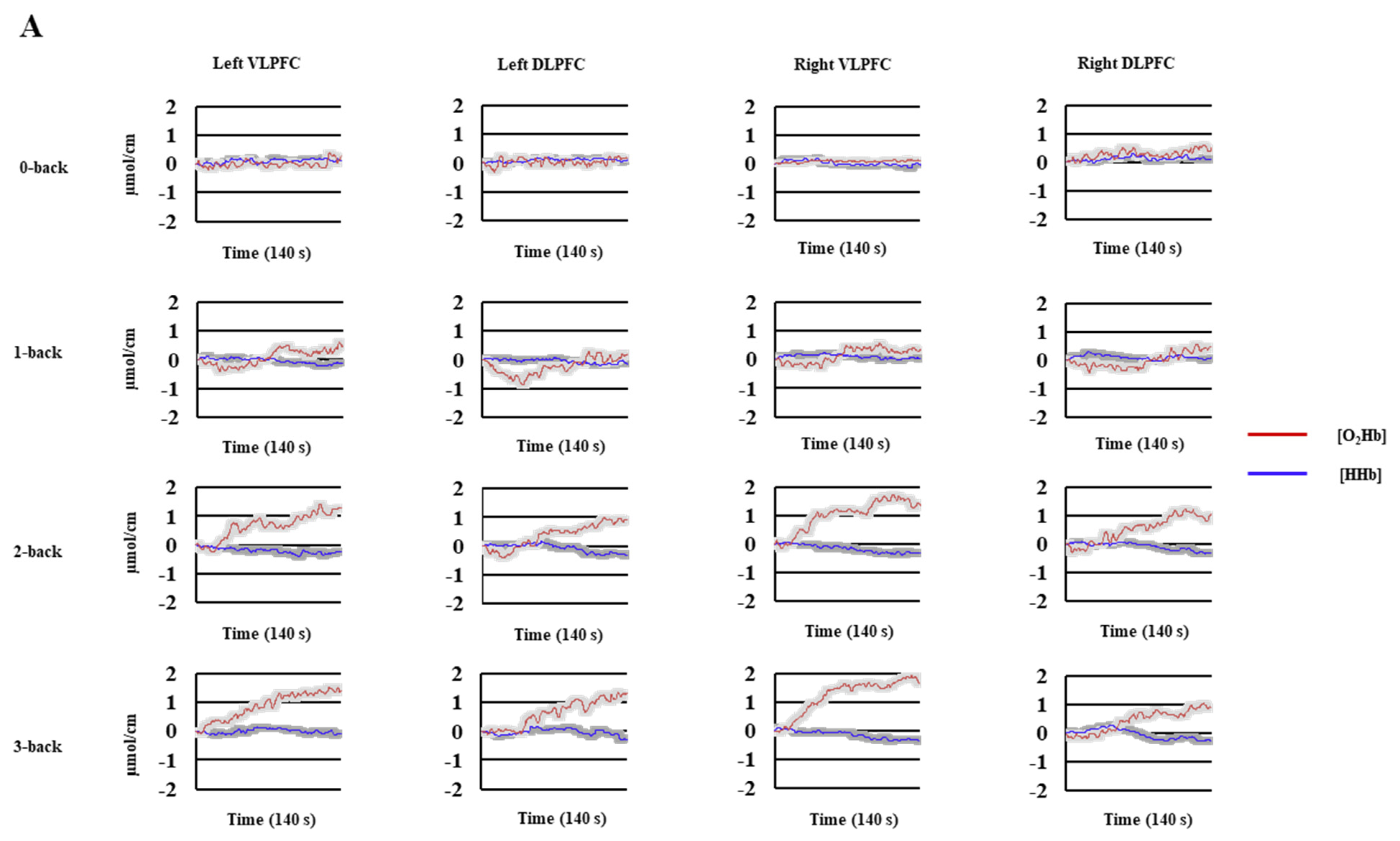
3.3.1. n-Back
3.3.2. RNG
3.4. Relationships between Behavioral Performance, Perceived Difficulty, and [O2Hb] Changes
3.4.1. n-Back
3.4.2. RNG
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Collette, F.; Hogge, M.; Salmon, E.; Van Der Linden, M. Exploration of the neural substrates of executive functioning by functional neuroimaging. Neuroscience 2006, 139, 209–221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diamond, A. Executive Functions. Ann. Rev. Psychol. 2013, 64, 135–168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friedman, N.P.; Miyake, A. The relations among inhibition and interference control functions: A latent-variable analysis. J. Exp. Psychol. Gen. 2004, 133, 101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nigg, J.T. On inhibition/disinhibition in developmental psychopathology: Views from cognitive and personality psychology and a working inhibition taxonomy. Psychol. Bull. 2000, 126, 220. [Google Scholar] [CrossRef] [PubMed]
- Braver, T.S.; Cohen, J.D.; Nystrom, L.E.; Jonides, J.; Smith, E.E.; Noll, D.C.A. Parametric study of prefrontal cortex involvement in human working memory. NeuroImage 1997, 5, 49–62. [Google Scholar] [CrossRef]
- Morris, N.; Jones, D.M. Memory updating in working memory : The role of the central executive. Br. J. Psychol. 1990, 81, 111. [Google Scholar] [CrossRef]
- Fishburn, F.A.; Norr, M.E.; Medvedev, A.V.; Vaidya, C.J. Sensitivity of fNIRS to cognitive state and load. Front. Hum. Neurosci. 2014, 8, 76. [Google Scholar] [CrossRef]
- Molteni, E.; Contini, D.; Caffini, M.; Baselli, G.; Spinelli, L.; Cubeddu, R.; Cerutti, S.; Bianchi, A.M.; Torricelli, A. Load-dependent brain activation assessed by time-domain functional near-infrared spectroscopy during a working memory task with graded levels of difficulty. J. Biomed. Opt. 2012, 17, 056005. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; He, W.; Wu, J.; Zhang, J.; Jin, Z.; Li, L. A coordinate-based meta-analysis of the n-back working memory paradigm using activation likelihood estimation. Brain Cogn. 2019, 132, 1–12. [Google Scholar] [CrossRef]
- Albinet, C.T.; Mandrick, K.; Bernard, P.L.; Perrey, S.; Blain, H. Improved cerebral oxygenation response and executive performance as a function of cardiorespiratory fitness in older women: A fNIRS study. Front. Aging. Neurosci. 2014, 6, 272. [Google Scholar] [CrossRef]
- Daniels, C.; Witt, K.; Wolff, S.; Jansen, O.; Deuschl, G. Rate dependency of the human cortical network subserving executive functions during generation of random number series-a functional magnetic resonance imaging study. Neurosci. Lett. 2003, 345, 25–28. [Google Scholar] [CrossRef]
- Jahanshahi, M.; Dirnberger, G.; Fuller, R.; Frith, C.D. The role of the dorsolateral prefrontal cortex in random number generation: A study with positron emission tomography. Neuroimage 2000, 12, 713–725. [Google Scholar] [CrossRef]
- Owen, A.M.; McMillan, K.M.; Laird, A.R.; Bullmore, E. N-back working memory paradigm: A meta-analysis of normative functional neuroimaging studies. Hum. Brain. Mapp. 2005, 25, 46–59. [Google Scholar] [CrossRef] [Green Version]
- Li, T.; Luo, Q.; Gong, H. Gender-specific hemodynamics in prefrontal cortex during a verbal working memory task by near-infrared spectroscopy. Behav. Brain. Res. 2010, 209, 148–153. [Google Scholar] [CrossRef]
- Agbangla, N.F.; Audiffren, M.; Pylouster, J.; Albinet, C.T. Working Memory, Cognitive Load and cardiorespiratory fitness: Testing the CRUNCH model with Near-Infrared Spectroscopy. Brain Sci. 2019, 9, 38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saikia, M.J.; Besio, W.G.; Mankodiya, K. The Validation of a Portable Functional NIRS System for Assessing Mental Workload. Sensors 2021, 21, 3810. [Google Scholar] [CrossRef]
- Vermeij, A.; van Beek, A.H.; Olde Rikkert, M.G.; Claassen, J.A.; Kessels, R.P. Effects of aging on cerebral oxygenation during working-memory performance: A functional near-infrared spectroscopy study. PLoS ONE 2012, 7, e46210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoshi, Y.; Tsou, B.H.; Billock, V.A.; Tanosaki, M.; Iguchi, Y.; Shimada, M.; Shinba, T.; Yamada, Y.; Oda, I. Spatiotemporal characteristics of hemodynamic changes in the human lateral prefrontal cortex during working memory tasks. Neuroimage 2003, 20, 1493–1504. [Google Scholar] [CrossRef]
- Ayaz, H.; Shewokis, P.A.; Bunce, S.; Izzetoglu, K.; Willems, B.; Onaral, B. Optical brain monitoring for operator training and mental workload assessment. NeuroImage 2012, 59, 36–47. [Google Scholar] [CrossRef] [PubMed]
- Herff, C.; Heger, D.; Fortmann, O.; Hennrich, J.; Putze, F.; Schultz, T. Mental workload during n-back task-quantified in the prefrontal cortex using fNIRS. Front. Hum. Neurosci. 2014, 7, 935. [Google Scholar] [CrossRef] [PubMed]
- Mattay, V.S.; Fera, F.; Tessitore, A.; Hariri, A.R.; Berman, K.F.; Das, S.; Meyer-Lindenberg, A.; Goldberg, T.E.; Callicott, J.H.; Weinberger, D.R. Neurophysiological correlates of age-related changes in working memory capacity. Neurosci Lett. 2006, 392, 32–37. [Google Scholar] [CrossRef] [PubMed]
- Cappell, K.A.; Gmeindl, L.; Reuter-Lorenz, P.A. Age differences in prefontal recruitment during verbal working memory maintenance depend on memory load. Cortex 2010, 46, 462–473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schneider-Garces, N.J.; Gordon, B.A.; Brumback-Peltz, C.R.; Shin, E.; Lee, Y.; Sutton, B.P.; Maclin, E.L.; Gratton, G.; Fabiani, M. Span, CRUNCH, and beyond: Working memory capacity and the aging brain. J. Cogn. Neurosci. 2010, 22, 655–669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, D.C.; Reuter-Lorenz, P. The adaptive brain: Aging and neurocognitive scaffolding. Annu. Rev. Psychol. 2009, 60, 173–196. [Google Scholar] [CrossRef] [Green Version]
- Reuter-Lorenz, P.A.; Cappell, K.A. Neurocognitive aging and the compensation hypothesis. Curr. Dir. Psychol. Sci. 2008, 17, 177–182. [Google Scholar] [CrossRef]
- Reuter-Lorenz, P.A.; Park, D.C. How does it STAC up? Revisiting the scaffolding theory of aging and cognition. Neuropsychol. Rev. 2014, 24, 355–370. [Google Scholar] [CrossRef] [Green Version]
- Cabeza, R.; Albert, M.; Belleville, S.; Craik, F.I.M.; Duarte, A.; Grady, C.L.; Lindenberger, U.; Nyberg, L.; Park, D.C.; Reuter-Lorenz, P.A.; et al. Maintenance, reserve and compensation: The cognitive neuroscience of healthy ageing. Nat. Rev. Neurosci. 2018, 19, 701–710. [Google Scholar] [CrossRef]
- McDonough, I.M.; Madan, C.R. Structural complexity is negatively associated with brain activity: A novel multimodal test of compensation theories of aging. Neurobiol. Aging 2021, 98, 185–196. [Google Scholar] [CrossRef]
- Morcom, A.M.; Henson, R.N.A. Increased Prefrontal Activity with Aging Reflects Nonspecific Neural Responses Rather than Compensation. J. Neurosci. 2018, 38, 7303–7313. [Google Scholar] [CrossRef] [Green Version]
- Mandrick, K.; Chua, Z.; Causse, M.; Perrey, S.; Dehais, F. Why a Comprehensive Understanding of Mental Workload through the Measurement of Neurovascular Coupling Is a Key Issue for Neuroergonomics? Front. Hum. Neurosci. 2016, 10, 250. [Google Scholar] [CrossRef]
- Causse, M.; Chua, Z.; Peysakhovich, V.; Del Campo, N.; Matton, N. Mental workload and neural efficiency quantified in the prefrontal cortex using fNIRS. Sci. Rep. 2017, 7, 5222. [Google Scholar] [CrossRef] [Green Version]
- Matsuda, G.; Hiraki, K. Sustained decrease in oxygenated hemoglobin during video games in the dorsal prefrontal cortex: A NIRS study of children. Neuroimage 2006, 29, 706–711. [Google Scholar] [CrossRef] [PubMed]
- Neubauer, A.C.; Fink, A. Intelligence and neural efficiency. Neurosci. Biobehav. Rev. 2009, 33, 1004–1023. [Google Scholar] [CrossRef]
- Dunst, B.; Benedek, M.; Jauk, E.; Bergner, S.; Koschutnig, K.; Sommer, M.; Ischebeck, A.; Spinath, B.; Arendasy, M.; Bühner, M.; et al. Neural efficiency as a function of task demands. Intelligence 2014, 42, 22–30. [Google Scholar] [CrossRef] [Green Version]
- Hart, S.G.; Staveland, L.E. Development of NASA-TLX (Task Load Index): Results of empirical and theoretical research. In Human Mental Workload; Hancock, P.A., Meshkati, N., Eds.; Elsevier Science Publishers: North-Holland, The Netherlands, 1988; pp. 139–184. [Google Scholar]
- Delignières, D.; Famose, J.P.; Genty, J. Validation d’une échelle de catégories pour la perception de la difficulté. Rev. STAPS 1994, 34, 77–88. [Google Scholar]
- Jonides, J.; Schumacher, E.H.; Smith, E.E.; Lauber, E.J.; Awh, E.; Minoshima, S.; Koeppe, R.A. Verbal working memory load affects regional brain activation as measured by PET. J. Cogn. Neurosci. 1997, 9, 462–475. [Google Scholar] [CrossRef]
- Stanislaw, H.; Todorov, N. Calculation of signal detection theory measures. Behav. Res. Methods Instrum. Comput. 1999, 31, 137–149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Towse, J.N.; Neil, D. Analyzing human random generation behavior: A review of methods used and a computer program for describing performance. Behav. Res. Methods Instrum. Comput. 1998, 30, 583–591. [Google Scholar] [CrossRef] [Green Version]
- Audiffren, M.; Tomporowski, P.D.; Zagrodnik, J. Acute aerobic exercise and information processing: Modulation of executive control in a Random Number Generation task. Acta Psychol. 2009, 132, 85–95. [Google Scholar] [CrossRef]
- Towse, J.N.; Valentine, J.D. Random Generation of Numbers: A search for underlying processes. Eur. J. Cogn. Psychol. 1997, 9, 381–400. [Google Scholar] [CrossRef]
- Shinba, T.; Nagano, M.; Kariya, N.; Ogawa, K.; Shinozaki, T.; Shimosato, S.; Hoshi, Y. Near-infrared spectroscopy analysis of frontal lobe dysfunction in schizophrenia. Biol. Psychiatry 2004, 55, 154–164. [Google Scholar] [CrossRef]
- Duncan, A.; Meek, J.H.; Clemence, M.; Elwell, C.E.; Fallon, P.; Tyszczuk, L.; Cope, M.; Delpy, D.T. Measurement of cranial optical path length as a function of age using phase resolved near infrared spectroscopy. Pediatr. Res. 1996, 39, 889–894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okamoto, M.; Dan, H.; Sakamoto, K.; Takeo, K.; Shimizu, K.; Kohno, S.; Oda, I.; Isobe, S.; Suzuki, T.; Kohyama, K.; et al. Three-dimensional probabilistic anatomical cranio-cerebral correlation via the international 10–20 system oriented for transcranial functional brain mapping. Neuroimage 2004, 21, 99–111. [Google Scholar] [CrossRef] [PubMed]
- Strangman, G.; Franceschini, M.A.; Boas, D.A. Factors affecting the accuracy of near-infrared spectroscopy concentration calculations for focal changes in oxygenation parameters. NeuroImage 2003, 18, 865–879. [Google Scholar] [CrossRef]
- Scholkmann, F.; Spichtig, S.; Muehlemann, T.; Wolf, M. How to detect and reduce movement artifacts in near-infrared imaging using moving standard deviation and spline interpolation. Physiol. Meas. 2010, 31, 649–662. [Google Scholar] [CrossRef] [Green Version]
- Limongi, T.; Di Sante, G.; Ferrari, M.; Quaresima, V. Detecting mental calculation related frontal cortex oxygenation changes for brain computer interface using multi-channel functional near infrared topography. Int. J. Bioelectromagn. 2009, 11, 86–90. [Google Scholar]
- Gagnon, C.; Desjardins-Crépeau, L.; Tournier, I.; Desjardins, M.; Lesage, F.; Greenwood, C.E.; Bherer, L. Near-infrared imaging of the effects of glucose ingestion and regulation on prefrontal activation during dual-task execution in healthy fasting older adults. Behav. Brain. Res. 2012, 232, 137–147. [Google Scholar] [CrossRef]
- Oldfield, R.C. The assessment and analysis of handedness: The Edinburgh inventory. Neuropsychologia 1971, 9, 97–113. [Google Scholar] [CrossRef]
- Reese, H.W. Counterbalancing and other uses of repeated-measures Latin-square designs: Analyses and interpretations. J. Exp. Child Psychol. 1997, 64, 137–158. [Google Scholar] [CrossRef]
- Borg, G. Perceived exertion as an indicator of somatic stress. Scand. J. Rehabil. Med. 1970, 2, 92–98. [Google Scholar] [PubMed]
- Howell, D.C. Statistical Methods for Psychology; PWS-Kent Publishing Co.: Boston, MA, USA, 1992. [Google Scholar]
- Albinet, C.T.; Tomporowski, P.D.; Beasman, K. Aging and concurrent task performance: Cognitive demand and motor control. Educ. Gerontol. 2006, 32, 689–706. [Google Scholar] [CrossRef]
- Jahanshahi, M.; Saleem, T.; Ho, A.K.; Dirnberger, G.; Fuller, R. Random number generation as an index of controlled processing. Neuropsychol 2006, 20, 391–399. [Google Scholar] [CrossRef]
- Cain, B. A Review of the Mental Workload Literature; NATO RTO Report, RTO-TR-HFM-121-Part-II; NATO: Brussels, Belgium, 2007. [Google Scholar]
- Longo, L.; Wickens, C.D.; Hancock, P.A.; Hancock, G.M. Human Mental Workload: A Survey and a Novel Inclusive Definition. Front. Psychol. 2022, 13, 883321. [Google Scholar] [CrossRef]
- Sandrini, M.; Fertonani, A.; Cohen, L.G.; Miniussi, C. Double dissociation of working memory load effects induced by bilateral parietal modulation. Neuropsychologia 2012, 50, 396–402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smirni, D.; Turriziani, P.; Mangano, G.R.; Cipolotti, L.; Oliveri, M. Modulating Memory Performance in Healthy Subjects with Transcranial Direct Current Stimulation Over the Right Dorsolateral Prefrontal Cortex. PLoS ONE 2015, 10, e0144838. [Google Scholar] [CrossRef] [PubMed]
- Pinti, P.; Scholkmann, F.; Hamilton, A.; Burgess, P.; Tachtsidis, I. Current Status and Issues RegardingPre-processing of fNIRS Neuroimaging Data: An Investigation of Diverse Signal Filtering Methods Within a General Linear Model Framework. Front. Hum. Neurosci. 2019, 12, 505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tachtsidis, I.; Scholkmann, F. False positives and false negatives in functional near-infrared spectroscopy: Issues, challenges, and the way forward. Neurophotonics 2016, 3, 031405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yücel, M.A.; Lühmann, A.V.; Scholkmann, F.; Gervain, J.; Dan, I.; Ayaz, H.; Boas, D.; Cooper, R.J.; Culver, J.; Elwell, C.E.; et al. Best practices for fNIRS publications. Neurophotonics 2021, 8, 012101. [Google Scholar] [CrossRef]
- Levy, B.J.; Wagner, A.D. Cognitive control and right ventrolateral prefrontal cortex: Reflexive reorienting, motor inhibition, and action updating. Ann. N. Y. Acad. Sci. 2011, 1224, 40–62. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control | Cognitive Load 1 | Cognitive Load 2 | Cognitive Load 3 | ||
|---|---|---|---|---|---|
| n-back | A’ (accuracy) | 0.99 ± 0.01 | 0.98 ± 0.02 NS | 0.93 ± 0.06 *† | 0.85 ± 0.06 *†‡ |
| RT (ms) | 367 ± 41 | 404 ± 74 NS | 517 ± 153 *† | 593 ± 186 *†‡ | |
| RNG | Success rate | - | 1±0 | 0.99 ± 0.01 NS | 0.98 ± 0.02 † |
| Adjacency (%) | - | 25.93 ± 8.08 | 26.88 ± 8.57 NS | 29.91 ± 9.15 †‡ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Agbangla, N.F.; Audiffren, M.; Pylouster, J.; Albinet, C.T. Load-Dependent Prefrontal Cortex Activation Assessed by Continuous-Wave Near-Infrared Spectroscopy during Two Executive Tasks with Three Cognitive Loads in Young Adults. Brain Sci. 2022, 12, 1462. https://doi.org/10.3390/brainsci12111462
Agbangla NF, Audiffren M, Pylouster J, Albinet CT. Load-Dependent Prefrontal Cortex Activation Assessed by Continuous-Wave Near-Infrared Spectroscopy during Two Executive Tasks with Three Cognitive Loads in Young Adults. Brain Sciences. 2022; 12(11):1462. https://doi.org/10.3390/brainsci12111462
Chicago/Turabian StyleAgbangla, Nounagnon Frutueux, Michel Audiffren, Jean Pylouster, and Cédric T. Albinet. 2022. "Load-Dependent Prefrontal Cortex Activation Assessed by Continuous-Wave Near-Infrared Spectroscopy during Two Executive Tasks with Three Cognitive Loads in Young Adults" Brain Sciences 12, no. 11: 1462. https://doi.org/10.3390/brainsci12111462