
Food Restriction Counteracts Dexamethasone-Induced Downregulation of Genes Involved in Cholesterol Homeostasis in Rat Brain during Aging
 , and
, and 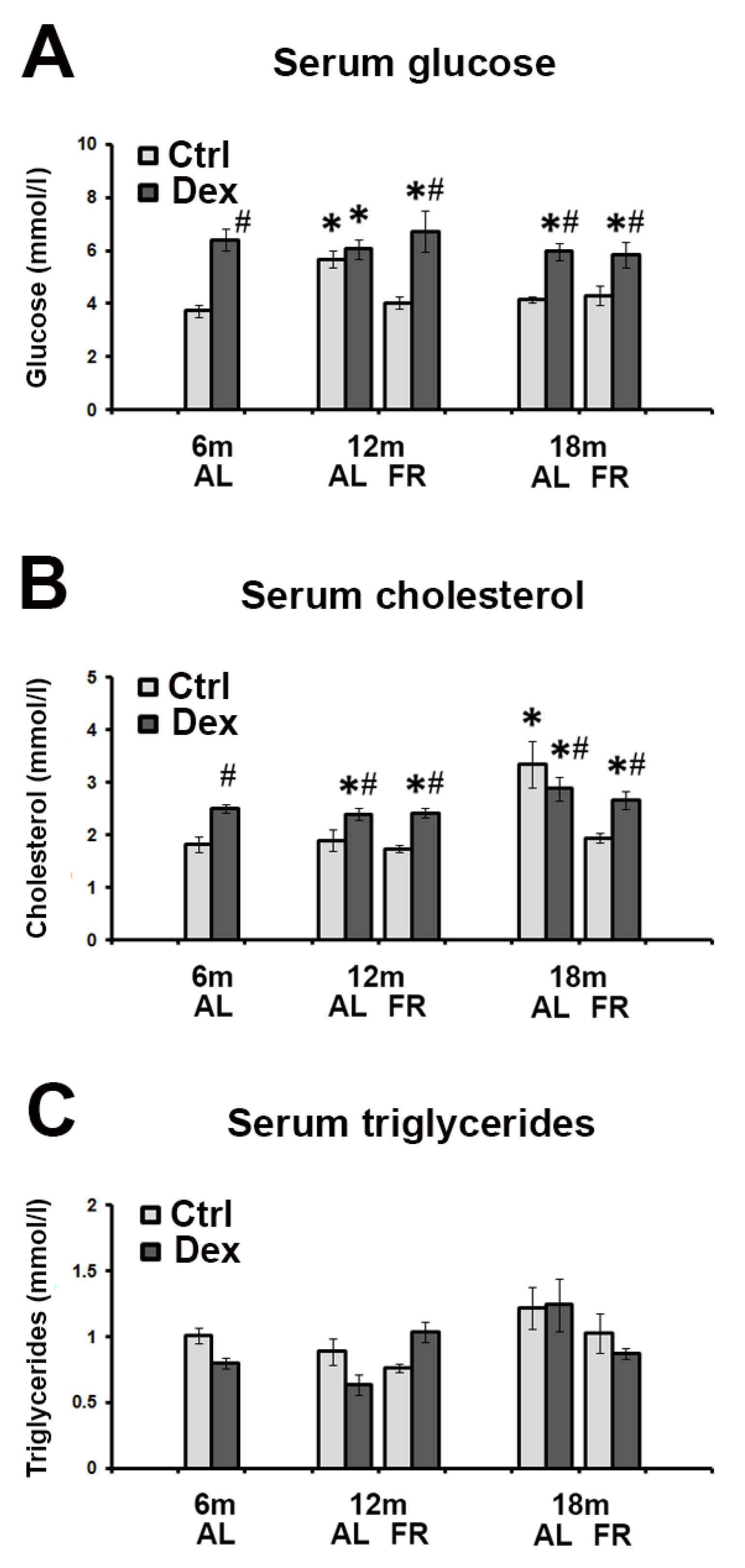{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Treatments
2.2. Analysis of Serum Biochemical Parameters (Glucose, Cholesterol, Triglycerides)
2.3. Real-Time RT-PCR
2.4. Statistical Analysis
3. Results
3.1. Biochemical Parameters in Rat Serum following Dexamethasone Treatment—The Effects of Aging and FR
3.2. FR Reversed Dexamethasone-Induced Decrease in the HMGCR mRNA Levels during Aging
3.3. FR Had No Significant Effect on Age-Related Changes in ApoE mRNA Expression following the Treatment with Dexamethasone
3.4. The Effects of Dexamethasone Treatment on CYP46A1 mRNA Levels during Aging
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- McDonough, A.K.; Curtis, J.R.; Saag, K.G. The Epidemiology of Glucocorticoid-Associated Adverse Events. Curr. Opin. Rheumatol. 2008, 20, 131–137. [Google Scholar] [CrossRef]
- Lupien, S.J.; De Leon, M.; De Santi, S.; Convit, A.; Tarshish, C.; Nair, N.P.V.; Thakur, M.; McEwen, B.S.; Hauger, R.L.; Meaney, M.J. Cortisol Levels during Human Aging Predict Hippocampal Atrophy and Memory Deficits. Nat. Neurosci. 1998, 1, 69–73. [Google Scholar] [CrossRef] [PubMed]
- Varney, N.R.; Alexander, B.; MacIndoe, J.H. Reversible Steroid Dementia in Patients without Steroid Psychosis. Am. J. Psychiatry 1984, 141, 369–372. [Google Scholar] [CrossRef]
- Lupien, S.J.; Wilkinson, C.W.; Brière, S.; Ng Ying Kin, N.M.K.; Meaney, M.J.; Nair, N.P.V. Acute Modulation of Aged Human Memory by Pharmacological Manipulation of Glucocorticoids. J. Clin. Endocrinol. Metab. 2002, 87, 3798–3807. [Google Scholar] [CrossRef]
- Schreurs, B.G. The Effects of Cholesterol on Learning and Memory. Neurosci. Biobehav. Rev. 2010, 34, 1366–1379. [Google Scholar] [CrossRef]
- Anstey, K.J.; Ashby-Mitchell, K.; Peters, R. Updating the Evidence on the Association between Serum Cholesterol and Risk of Late-Life Dementia: Review and Meta-Analysis. J. Alzheimers. Dis. 2017, 56, 215–228. [Google Scholar] [CrossRef]
- Blalock, E.M.; Chen, K.C.; Sharrow, K.; Herman, J.P.; Porter, N.M.; Foster, T.C.; Landfield, P.W. Gene Microarrays in Hippocampal Aging: Statistical Profiling Identifies Novel Processes Correlated with Cognitive Impairment. J. Neurosci. 2003, 23, 3807–3819. [Google Scholar] [CrossRef]
- Kadish, I.; Thibault, O.; Blalock, E.M.; Chen, K.C.; Gant, J.C.; Porter, N.M.; Landfield, P.W. Hippocampal and Cognitive Aging across the Lifespan: A Bioenergetic Shift Precedes and Increased Cholesterol Trafficking Parallels Memory Impairment. J. Neurosci. 2009, 29, 1805–1816. [Google Scholar] [CrossRef]
- Dietschy, J.M.; Turley, S.D. Cholesterol Metabolism in the Brain. Curr. Opin. Lipidol. 2001, 12, 105–112. [Google Scholar] [CrossRef]
- Mauch, D.H.; Nägier, K.; Schumacher, S.; Göritz, C.; Müller, E.C.; Otto, A.; Pfrieger, F.W. CNS Synaptogenesis Promoted by Glia-Derived Cholesterol. Science 2001, 294, 1354–1357. [Google Scholar] [CrossRef] [PubMed]
- Frank, C.; Rufini, S.; Tancredi, V.; Forcina, R.; Grossi, D.; D’Arcangelo, G. Cholesterol Depletion Inhibits Synaptic Transmission and Synaptic Plasticity in Rat Hippocampus. Exp. Neurol. 2008, 212, 407–414. [Google Scholar] [CrossRef] [PubMed]
- Martin, M.G.; Ahmed, T.; Korovaichuk, A.; Venero, C.; Menchón, S.A.; Salas, I.; Munck, S.; Herreras, O.; Balschun, D.; Dotti, C.G. Constitutive Hippocampal Cholesterol Loss Underlies Poor Cognition in Old Rodents. EMBO Mol. Med. 2014, 6, 902–917. [Google Scholar] [CrossRef]
- Korinek, M.; Gonzalez-Gonzalez, I.M.; Smejkalova, T.; Hajdukovic, D.; Skrenkova, K.; Krusek, J.; Horak, M.; Vyklicky, L. Cholesterol Modulates Presynaptic and Postsynaptic Properties of Excitatory Synaptic Transmission. Sci. Rep. 2020, 10, 12651. [Google Scholar] [CrossRef]
- Snipes, G.J.; Suter, U. Cholesterol and Myelin. Subcell. Biochem. 1997, 28, 173–204. [Google Scholar] [CrossRef] [PubMed]
- Poirier, J. Apolipoprotein E and Cholesterol Metabolism in the Pathogenesis and Treatment of Alzheimer’s Disease. Trends Mol. Med. 2003, 9, 94–101. [Google Scholar] [CrossRef]
- Wahrle, S.; Das, P.; Nyborg, A.C.; McLendon, C.; Shoji, M.; Kawarabayashi, T.; Younkin, L.H.; Younkin, S.G.; Golde, T.E. Cholesterol-Dependent Gamma-Secretase Activity in Buoyant Cholesterol-Rich Membrane Microdomains. Neurobiol. Dis. 2002, 9, 11–23. [Google Scholar] [CrossRef]
- Lund, E.G.; Xie, C.; Kotti, T.; Turley, S.D.; Dietschy, J.M.; Russell, D.W. Knockout of the Cholesterol 24-Hydroxylase Gene in Mice Reveals a Brain-Specific Mechanism of Cholesterol Turnover. J. Biol. Chem. 2003, 278, 22980–22988. [Google Scholar] [CrossRef]
- Smiljanic, K.; Vanmierlo, T.; Djordjevic, A.M.; Perovic, M.; Loncarevic-Vasiljkovic, N.; Tesic, V.; Rakic, L.; Ruzdijic, S.; Lutjohann, D.; Kanazir, S. Aging Induces Tissue-Specific Changes in Cholesterol Metabolism in Rat Brain and Liver. Lipids 2013, 48, 1069–1077. [Google Scholar] [CrossRef]
- Perovic, M.; Mladenovic Djordjevic, A.; Smiljanic, K.; Tanic, N.; Rakic, L.; Ruzdijic, S.; Kanazir, S. Expression of Cholesterol Homeostasis Genes in the Brain of the Male Rat Is Affected by Age and Dietary Restriction. Biogerontology 2009, 10, 735–745. [Google Scholar] [CrossRef]
- Witte, A.V.; Fobker, M.; Gellner, R.; Knecht, S.; Flöel, A. Caloric Restriction Improves Memory in Elderly Humans. Proc. Natl. Acad. Sci. USA 2009, 106, 1255–1260. [Google Scholar] [CrossRef] [Green Version]
- Mattson, M.P.; Longo, V.D.; Harvie, M. Impact of Intermittent Fasting on Health and Disease Processes. Ageing Res. Rev. 2017, 39, 46–58. [Google Scholar] [CrossRef] [PubMed]
- Masoro, E. The Role of Hormesis in Life Extension by Dietary Restriction. Interdiscip. Top. Gerontol. 2007, 35, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Tesic, V.; Perovic, M.; Lazic, D.; Kojic, S.; Smiljanic, K.; Ruzdijic, S.; Rakic, L.; Kanazir, S. Long-Term Intermittent Feeding Restores Impaired GR Signaling in the Hippocampus of Aged Rat. J. Steroid Biochem. Mol. Biol. 2015, 149, 43–52. [Google Scholar] [CrossRef] [PubMed]
- Perović, M.; Jović, M.; Todorović, S.; Đorđević, A.M.; Milanović, D.; Kanazir, S.; Lončarević-Vasiljković, N. Neuroprotective Effects of Food Restriction in a Rat Model of Traumatic Brain Injury–the Role of Glucocorticoid Signaling. Nutr. Neurosci. 2020, 25, 537–549. [Google Scholar] [CrossRef] [PubMed]
- Tanic, N.; Perovic, M.; Mladenovic, A.; Ruzdijic, S.; Kanazir, S. Effects of Aging, Dietary Restriction and Glucocorticoid Treatment on Housekeeping Gene Expression in Rat Cortex and Hippocampus–Evaluation by Real Time RT-PCR. J. Mol. Neurosci. 2007, 32, 38–46. [Google Scholar] [CrossRef]
- Birmingham, M.K.; Sar, M.; Stumpf, W.E. Dexamethasone Target Sites in the Central Nervous System and Their Potential Relevance to Mental Illness. Cell. Mol. Neurobiol. 1993, 13, 373–386. [Google Scholar] [CrossRef]
- Cole, M.A.; Kim, P.J.; Kalman, B.A.; Spencer, R.L. Dexamethasone Suppression of Corticosteroid Secretion: Evaluation of the Site of Action by Receptor Measures and Functional Studies. Psychoneuroendocrinology 2000, 25, 151–167. [Google Scholar] [CrossRef]
- Rupprecht, R.; Reul, J.M.H.M.; van Steensel, B.; Spengler, D.; Söder, M.; Berning, B.; Holsboer, F.; Damm, K. Pharmacological and Functional Characterization of Human Mineralocorticoid and Glucocorticoid Receptor Ligands. Eur. J. Pharmacol. 1993, 247, 145–154. [Google Scholar] [CrossRef]
- Sapolsky, R.M.; Krey, L.C.; McEwen, B.S. The Neuroendocrinology of Stress and Aging: The Glucocorticoid Cascade Hypothesis. Endocr. Rev. 1986, 7, 284–301. [Google Scholar] [CrossRef]
- Arumugam, T.V.; Phillips, T.M.; Cheng, A.; Morrell, C.H.; Mattson, M.P.; Wan, R. Age and Energy Intake Interact to Modify Cell Stress Pathways and Stroke Outcome. Ann. Neurol. 2010, 67, 41–52. [Google Scholar] [CrossRef] [Green Version]
- Bruce-Keller, A.J.; Umberger, G.; McFall, R.; Mattson, M.P. Food Restriction Reduces Brain Damage and Improves Behavioral Outcome Following Excitotoxic and Metabolic Insults. Ann. Neurol. 1999, 45, 8–15. [Google Scholar] [CrossRef]
- Halagappa, V.K.M.; Guo, Z.; Pearson, M.; Matsuoka, Y.; Cutler, R.G.; LaFerla, F.M.; Mattson, M.P. Intermittent Fasting and Caloric Restriction Ameliorate Age-Related Behavioral Deficits in the Triple-Transgenic Mouse Model of Alzheimer’s Disease. Neurobiol. Dis. 2007, 26, 212–220. [Google Scholar] [CrossRef] [PubMed]
- Lončarević-Vasiljković, N.; Pešić, V.; Tanić, N.; Milanović, D.; Popić, J.; Kanazir, S.; Ruždijić, S. Changes in Markers of Neuronal and Glial Plasticity after Cortical Injury Induced by Food Restriction. Exp. Neurol. 2009, 220, 198–206. [Google Scholar] [CrossRef] [PubMed]
- Guarnieri, D.J.; Brayton, C.E.; Richards, S.M.; Maldonado-Aviles, J.; Trinko, J.R.; Nelson, J.; Taylor, J.R.; Gourley, S.L.; Dileone, R.J. Gene Profiling Reveals a Role for Stress Hormones in the Molecular and Behavioral Response to Food Restriction. Biol. Psychiatry 2012, 71, 358–365. [Google Scholar] [CrossRef] [PubMed]
- Brown, M.S.; Goldstein, J.L. Cholesterol Feedback: From Schoenheimer’s Bottle to Scap’s MELADL. J. Lipid Res. 2009, 50, S15–S27. [Google Scholar] [CrossRef] [PubMed]
- Rodwell, V.W.; Nordstrom, J.L.; Mitschelen, J.J. Regulation of HMG-CoA Reductase. Adv. Lipid Res. 1976, 14, 1–74. [Google Scholar] [CrossRef]
- Cavenee, W.K.; Melnykovych, G. Induction of 3-Hydroxy-3-Methylglutaryl Coenzyme A Reductase in HeLa Cells by Glucocorticoids. J. Biol. Chem. 1977, 252, 3272–3276. [Google Scholar] [CrossRef]
- Sato, T.; Yamamoto, H.; Sawada, N.; Nashiki, K.; Tsuji, M.; Muto, K.; Kume, H.; Sasaki, H.; Arai, H.; Nikawa, T.; et al. Restraint Stress Alters the Duodenal Expression of Genes Important for Lipid Metabolism in Rat. Toxicology 2006, 227, 248–261. [Google Scholar] [CrossRef]
- Pullinger, C.R.; Gibbons, G.F. The Role of Substrate Supply in the Regulation of Cholesterol Biosynthesis in Rat Hepatocytes. Biochem. J. 1983, 210, 625–632. [Google Scholar] [CrossRef]
- Thelen, K.M.; Falkai, P.; Bayer, T.A.; Lütjohann, D. Cholesterol Synthesis Rate in Human Hippocampus Declines with Aging. Neurosci. Lett. 2006, 403, 15–19. [Google Scholar] [CrossRef] [PubMed]
- Marino, M.; Pallottini, V.; D’eramo, C.; Cavallini, G.; Bergamini, E.; Trentalance, A. Age-Related Changes of Cholesterol and Dolichol Biosynthesis in Rat Liver. Mech. Ageing Dev. 2002, 123, 1183–1189. [Google Scholar] [CrossRef]
- Pallottini, V.; Martini, C.; Cavallini, G.; Bergamini, E.; Mustard, K.J.; Hardie, D.G.; Trentalance, A. Age-Related HMG-CoA Reductase Deregulation Depends on ROS-Induced P38 Activation. Mech. Ageing Dev. 2007, 128, 688–695. [Google Scholar] [CrossRef] [PubMed]
- Shinitzky, M. Patterns of Lipid Changes in Membranes of the Aged Brain. Gerontology 1987, 33, 149–154. [Google Scholar] [CrossRef] [PubMed]
- Rosenzweig, E.S.; Barnes, C.A. Impact of Aging on Hippocampal Function: Plasticity, Network Dynamics, and Cognition. Prog. Neurobiol. 2003, 69, 143–179. [Google Scholar] [CrossRef]
- Lane-Donovan, C.; Wong, W.M.; Durakoglugil, M.S.; Wasser, C.R.; Jiang, S.; Xian, X.; Herz, J. Genetic Restoration of Plasma ApoE Improves Cognition and Partially Restores Synaptic Defects in ApoE-Deficient Mice. J. Neurosci. 2016, 36, 10141–10150. [Google Scholar] [CrossRef]
- Gee, J.R.; Ding, Q.; Keller, J.N. Age-Related Alterations of Apolipoprotein E and Interleukin-1beta in the Aging Brain. Biogerontology 2006, 7, 69–79. [Google Scholar] [CrossRef]
- Maloney, B.; Ge, Y.W.; Alley, G.M.; Lahiri, D.K. Important Differences between Human and Mouse APOE Gene Promoters: Limitation of Mouse APOE Model in Studying Alzheimer’s Disease. J. Neurochem. 2007, 103, 1237–1257. [Google Scholar] [CrossRef]
- Ohyama, Y.; Meaney, S.; Heverin, M.; Ekström, L.; Brafman, A.; Shafir, M.; Andersson, U.; Olin, M.; Eggertsen, G.; Diczfalusy, U.; et al. Studies on the Transcriptional Regulation of Cholesterol 24-Hydroxylase (CYP46A1): Marked Insensitivity toward Different Regulatory Axes. J. Biol. Chem. 2006, 281, 3810–3820. [Google Scholar] [CrossRef]
- He, X.; Jenner, A.M.; Ong, W.Y.; Farooqui, A.A.; Patel, S.C. Lovastatin Modulates Increased Cholesterol and Oxysterol Levels and Has a Neuroprotective Effect on Rat Hippocampal Neurons after Kainate Injury. J. Neuropathol. Exp. Neurol. 2006, 65, 652–663. [Google Scholar] [CrossRef]
- Cartagena, C.M.; Burns, M.P.; Rebeck, G.W. 24S-Hydroxycholesterol Effects on Lipid Metabolism Genes Are Modeled in Traumatic Brain Injury. Brain Res. 2010, 1319, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Fon Tacer, K.; Pompon, D.; Rozman, D. Adaptation of Cholesterol Synthesis to Fasting and TNF-Alpha: Profiling Cholesterol Intermediates in the Liver, Brain, and Testis. J. Steroid Biochem. Mol. Biol. 2010, 121, 619–625. [Google Scholar] [CrossRef] [PubMed]
- Lütjohann, D.; Papassotiropoulos, A.; Björkhem, I.; Locatelli, S.; Bagli, M.; Oehring, R.D.; Schlegel, U.; Jessen, F.; Rao, M.L.; Von Bergmann, K.; et al. Plasma 24S-Hydroxycholesterol (Cerebrosterol) Is Increased in Alzheimer and Vascular Demented Patients. J. Lipid Res. 2000, 41, 195–198. [Google Scholar] [CrossRef]
- Vanmierlo, T.; Rutten, K.; Dederen, J.; Bloks, V.W.; van Vark-van der Zee, L.C.; Kuipers, F.; Kiliaan, A.; Blokland, A.; Sijbrands, E.J.G.; Steinbusch, H.; et al. Liver X Receptor Activation Restores Memory in Aged AD Mice without Reducing Amyloid. Neurobiol. Aging 2011, 32, 1262–1272. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ciric, J.; Tesic, V.; Milovanovic, N.; Jovanovic Macura, I.; Ivkovic, S.; Kanazir, S.; Perovic, M. Food Restriction Counteracts Dexamethasone-Induced Downregulation of Genes Involved in Cholesterol Homeostasis in Rat Brain during Aging. Brain Sci. 2022, 12, 1297. https://doi.org/10.3390/brainsci12101297
Ciric J, Tesic V, Milovanovic N, Jovanovic Macura I, Ivkovic S, Kanazir S, Perovic M. Food Restriction Counteracts Dexamethasone-Induced Downregulation of Genes Involved in Cholesterol Homeostasis in Rat Brain during Aging. Brain Sciences. 2022; 12(10):1297. https://doi.org/10.3390/brainsci12101297
Chicago/Turabian StyleCiric, Jelena, Vesna Tesic, Nikola Milovanovic, Irena Jovanovic Macura, Sanja Ivkovic, Selma Kanazir, and Milka Perovic. 2022. "Food Restriction Counteracts Dexamethasone-Induced Downregulation of Genes Involved in Cholesterol Homeostasis in Rat Brain during Aging" Brain Sciences 12, no. 10: 1297. https://doi.org/10.3390/brainsci12101297