
Insights into Dyslexia Genetics Research from the Last Two Decades


Abstract
:1. Clinical Diagnosis and Educational Identification and Manifestation of Dyslexia
2. Epidemiology of Dyslexia
3. Twin Studies of Dyslexia
4. Molecular Genetics Studies of Dyslexia
4.1. GWAS: The Sample Size Problem
4.2. GWAS for Reading Skills and Dyslexia
4.3. Polygenic Risk Scores
4.4. Rare Variants
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Shaywitz, B.A.; Shaywitz, S.E. The American experience: Towards a 21st century definition of dyslexia. Oxford Rev. Educ. 2020, 46, 454–471. [Google Scholar] [CrossRef]
- Share, D.L. Common misconceptions about the phonological deficit theory of dyslexia. Brain Sci. 2021, 11, 1510. [Google Scholar] [CrossRef]
- Lyon, G.R.; Shaywitz, S.E.; Shaywitz, B.A. A definition of dyslexia. Ann. Dyslexia 2003, 53, 1–14. [Google Scholar] [CrossRef]
- American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders, 5th ed.; American Psychiatric Association: Arlington, TX, USA, 2013. [Google Scholar] [CrossRef]
- Miciak, J.; Fletcher, J.M. The critical role of instructional response for identifying dyslexia and other learning disabilities. J. Learn Disabil. 2020, 53, 343–353. [Google Scholar] [CrossRef]
- Wagner, R.K.; Zirps, F.A.; Edwards, A.A.; Wood, S.G.; Joyner, R.E.; Becker, B.J.; Liu, G.; Beal, B. The prevalence of dyslexia: A new approach to its estimation. J. Learn. Disabil. 2020, 53, 354–365. [Google Scholar] [CrossRef]
- Erbeli, F.; Hart, S.A.; Wagner, R.K.; Taylor, J. Examining the etiology of reading disability as conceptualized by the hybrid model. Sci. Stud. Read. 2018, 22, 167–180. [Google Scholar] [CrossRef]
- Wagner, R.K.; Torgesen, J.K.; Rashotte, C.A. Comprehensive Test of Phonological Processing (CTOPP); PRO-ED: Austin, TX, USA, 1999. [Google Scholar]
- Woodcock, R.W.; Johnson, M.B. Manual for the Woodcock–Johnson Tests of Achievement—Revised; RCL Enterprises: Allen, TX, USA, 1990. [Google Scholar]
- Wiederholt, J.L.; Bryant, B.R. Gray Oral Reading Test: Examiner’s Manual, 5th ed.; Pro-Ed: Austin, TX, USA, 2012. [Google Scholar]
- Goulandris, N.E. Dyslexia in Different Languages: Cross-Linguistic Comparisons; Whurr Publishers: Philadephia, PA, USA, 2003. [Google Scholar]
- Song, S.; Zhang, Y.; Shu, H.; McBride, C. Universal and specific predictors of Chinese children with dyslexia—Exploring the cognitive deficits and subtypes. Front. Psychol. 2020, 10, 2904. [Google Scholar] [CrossRef] [Green Version]
- International Dyslexia Association. Testing and Evaluation. 2021. Available online: https://dyslexiaida.org/testing-and-evaluation (accessed on 17 November 2021).
- Chung, K.K.; Ho, C.S.H. Dyslexia in Chinese language: An overview of research and practice. Aust J. Learn. Diff. 2010, 15, 213–224. [Google Scholar] [CrossRef]
- Jiménez, J.E.; de la Cadena, C.G.; Siegel, L.S.; O’Shanahan, I.; Garcia, E.; Rodriguez, C. Gender ratio and cognitive profiles in dyslexia: A cross-national study. Read. Writ. 2011, 24, 729–747. [Google Scholar] [CrossRef]
- Shaywitz, S.E.; Escobar, M.D.; Shaywitz, B.A.; Fletcher, J.M.; Makuch, R. Evidence that dyslexia may represent the lower tail of the normal distribution of reading ability. N. Engl. J. Med. 1992, 326, 145–150. [Google Scholar] [CrossRef]
- Snowling, M.J.; Melby-Lervåg, M. Oral language deficits in familial dyslexia: A meta-analysis and review. Psychol. Bull. 2016, 142, 498–545. [Google Scholar] [CrossRef]
- Hoeft, F.; McCardle, P.; Pugh, K. The myths and truths of dyslexia in different writing systems. The Examiner. 2015. Available online: https://dyslexiaida.org/the-myths-and-truths-of-dyslexia (accessed on 20 December 2021).
- Jiménez, J.E.; Rodríguez, C.; Ramírez, G. Spanish developmental dyslexia: Prevalence, cognitive profile and home literacy experiences. J. Exp. Child. Psychol. 2009, 103, 167–185. [Google Scholar] [CrossRef]
- Sun, Z.; Zou, L.; Zhang, J.; Mo, S.; Shao, S.; Zhong, R.; Ke, J.; Lu, X.; Miao, X.; Song, R. Prevalence and associated risk factors of dyslexic children in a middle-sized city of China: A cross-sectional study. PLoS ONE 2013, 8, e56688. [Google Scholar] [CrossRef] [Green Version]
- Quinn, J.M.; Wagner, R.K. Gender differences in reading impairment and the identification of impaired readers: Results from a large-scale study of at-risk readers. J. Learn. Disabil. 2015, 48, 433–445. [Google Scholar] [CrossRef] [Green Version]
- Erbeli, F.; Hart, S.A.; Taylor, J. Genetic and environmental influences on achievement outcomes based on family history of learning disabilities status. J. Learn. Disabil. 2019, 52, 135–145. [Google Scholar] [CrossRef]
- Daucourt, M.C.; Erbeli, F.; Little, C.W.; Haughbrook, R.; Hart, S.A. A meta-analytical review of the genetic and environmental correlations between reading and attention-deficit/hyperactivity disorder symptoms and reading and math. Sci. Stud. Read. 2020, 24, 23–56. [Google Scholar] [CrossRef]
- Snowling, M.J.; Hayiou-Thomas, M.E.; Nash, H.M.; Hulme, C. Dyslexia and Developmental Language Disorder: Comorbid disorders with distinct effects on reading comprehension. J. Child. Psychol. Psychiatry 2020, 61, 672–680. [Google Scholar] [CrossRef] [Green Version]
- Landerl, K.; Moll, K. Comorbidity of learning disorders: Prevalence and familial transmission. J. Child. Psychol. Psychiatry 2010, 51, 287–294. [Google Scholar] [CrossRef]
- Erbeli, F.; Hart, S.A.; Taylor, J. Longitudinal associations among reading-related skills and reading comprehension: A twin study. Child. Dev. 2018, 89, e480–e493. [Google Scholar] [CrossRef]
- Paracchini, S.; Scerri, T.; Monaco, A.P. The genetic lexicon of dyslexia. Annu Rev. Genom. Hum. Genet. 2007, 8, 57–79. [Google Scholar] [CrossRef]
- Lohvansuu, K.; Torppa, M.; Ahonen, T.; Eklund, K.; Hamalainen, J.A.; Leppanen, P.H.T.; Lyytinen, H. Unveiling the mysteries of dyslexia—Lessons learned from the prospective Jyväskylä longitudinal study of dyslexia. Brain Sci. 2021, 11, 427. [Google Scholar] [CrossRef]
- Fisher, S.E.; DeFries, J.C. Developmental dyslexia: Genetic dissection of a complex cognitive trait. Nat. Rev. Neurosci. 2002, 3, 767–780. [Google Scholar] [CrossRef] [Green Version]
- Raskind, W.H.; Peter, B.; Richards, T.L.; Eckert, M.A.; Berninger, V.W. The genetics of reading disabilities: From phenotypes to candidate genes. Front. Psychol. 2013, 3, 601. [Google Scholar] [CrossRef] [Green Version]
- Andreola, C.; Mascheretti, S.; Belotti, R.; Ogliari, A.; Marino, C.; Battaglia, M.; Scaini, S. The heritability of reading and reading-related neurocognitive components: A multi-level meta-analysis. Neurosci. Biobehav. Rev. 2020, 121, 175–200. [Google Scholar] [CrossRef]
- Moll, K.; Loff, A.; Snowling, M.J. Cognitive endophenotypes of dyslexia. Sci. Stud. Read. 2013, 17, 1–13. [Google Scholar] [CrossRef]
- Bergen, S.E.; Gardner, C.O.; Kendler, K.S. Age-related changes in heritability of behavioral phenotypes over adolescence and young adulthood: A meta-analysis. Twin Res. Hum. Genet. 2007, 10, 423–433. [Google Scholar] [CrossRef]
- Wadsworth, S.J.; DeFries, J.C.; Olson, R.K.; Willcutt, E.G. Colorado longitudinal twin study of reading disability. Ann. Dyslexia 2007, 57, 139–160. [Google Scholar] [CrossRef]
- Hawke, J.L.; Wadsworth, S.J.; DeFries, J.C. Genetic influences on reading difficulties in boys and girls: The Colorado twin study. Dyslexia 2006, 12, 21–29. [Google Scholar] [CrossRef]
- Hawke, J.L.; Wadsworth, S.J.; Olson, R.K.; DeFries, J.C. Etiology of reading difficulties as a function of gender and severity. Read. Writ. 2007, 20, 13–25. [Google Scholar] [CrossRef]
- Wadsworth, S.J.; Knopik, V.S.; DeFries, J.C. Reading disability in boys and girls: No evidence for a differential genetic etiology. Read. Writ. 2000, 13, 133–145. [Google Scholar] [CrossRef]
- Plomin, R.; Kovas, Y. Generalist genes and learning disabilities. Psychol. Bull. 2005, 131, 592–617. [Google Scholar] [CrossRef] [Green Version]
- Whitehouse, A.J.; Spector, T.D.; Cherkas, L.F. No clear genetic influences on the association between dyslexia and anxiety in a population-based sample of female twins. Dyslexia 2009, 15, 282–290. [Google Scholar] [CrossRef]
- Willcutt, E.G.; Pennington, B.F. Psychiatric comorbidity in children and adolescents with reading disability. J. Child. Psychol Psychiatry 2000, 41, 1039–1048. [Google Scholar] [CrossRef]
- Newbury, D.F.; Monaco, A.P.; Paracchini, S. Reading and language disorders: The importance of both quantity and quality. Genes 2014, 5, 285–309. [Google Scholar] [CrossRef]
- Scerri, T.S.; Schulte-Korne, G. Genetics of developmental dyslexia. Eur. Child. Adolesc. Psychiatry 2010, 19, 179–197. [Google Scholar] [CrossRef] [Green Version]
- Galaburda, A.M.; LoTurco, J.; Ramus, F.; Fitch, R.H.; Rosen, G.D. From genes to behavior in developmental dyslexia. Nat. Neurosci. 2006, 9, 1213–1217. Available online: http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Citation&list_uids=17001339 (accessed on 20 December 2021). [CrossRef]
- Galaburda, A.M.; Kemper, T.L. Cytoarchitectonic abnormalities in developmental dyslexia: A case study. Ann. Neurol. 1979, 6, 94–100. [Google Scholar] [CrossRef]
- Galaburda, A.M.; Sherman, G.F.; Rosen, G.D.; Aboitz, F.; Geschwind, N.; Aboitiz, F.; Geschwind, N. Developmental dyslexia: Four consecutive patients with cortical anomalies. Ann. Neurol. 1985, 18, 222–233. [Google Scholar] [CrossRef]
- Humphreys, P.; Kaufmann, W.E.; Galaburda, A.M. Developmental dyslexia in women: Neuropathological findings in three patients. Ann. Neurol. 1990, 28, 727–738. [Google Scholar] [CrossRef]
- Guidi, L.G.; Velayos-Baeza, A.; Martinez-Garay, I.; Monaco, A.P.; Paracchini, S.; Bishop, D.V.M.; Molnár, Z. The neuronal migration hypothesis of dyslexia: A critical evaluation 30 years on. Eur. J. Neurosci. 2018, 48, 3212–3233. [Google Scholar] [CrossRef] [Green Version]
- Kere, J. The molecular genetics and neurobiology of developmental dyslexia as model of a complex phenotype. Biochem. Biophys. Res. Commun. 2014, 452, 236–243. [Google Scholar] [CrossRef] [Green Version]
- Paracchini, S.; Diaz, R.; Stein, J. Advances in dyslexia genetics—New insights into the role of brain asymmetries. Adv. Genet. 2016, 96, 53–97. [Google Scholar] [CrossRef]
- Ivliev, A.E.; Hoen, P.A.; van Roon-Mom, W.M.; Peters, D.J.; Sergeeva, M.G. Exploring the transcriptome of ciliated cells using in silico dissection of human tissues. PLoS ONE 2012, 7, e35618. [Google Scholar] [CrossRef]
- Diaz, R.; Kronenberg, N.M.; Martinelli, A.; Liehm, P.; Riches, A.C.; Gather, M.C.; Paracchini, S. KIAA0319 influences cilia length, cell migration and mechanical cell-substrate interaction. BioRxiv 2021, 780874. [Google Scholar] [CrossRef] [Green Version]
- Massinen, S.; Hokkanen, M.E.; Matsson, H.; Tammimies, K.; Tapia-Paez, I.; Dahlstrom-Heuser, V.; Kuja-Panula, J.; Burghoorn, J.; Jeppsson, K.E.; Swoboda, P.; et al. Increased expression of the dyslexia candidate gene DCDC2 affects length and signaling of primary cilia in neurons. PLoS ONE 2011, 6, e20580. [Google Scholar] [CrossRef]
- Tarkar, A.; Loges, N.T.; E Slagle, C.; Francis, R.; Dougherty, G.W.; Tamayo, J.V.; Shook, B.; Cantino, M.; Schwartz, D.; Jahnke, C.; et al. DYX1C1 is required for axonemal dynein assembly and ciliary motility. Nat. Genet. 2013, 45, 995–1003. [Google Scholar] [CrossRef]
- Schueler, M.; Braun, D.A.; Chandrasekar, G.; Gee, H.Y.; Klasson, T.D.; Halbritter, J.; Bieder, A.; Porath, J.D.; Airik, R.; Zhou, W.; et al. DCDC2 mutations cause a renal-hepatic ciliopathy by disrupting Wnt signaling. Am. J. Hum. Genet. 2015, 96, 81–92. [Google Scholar] [CrossRef] [Green Version]
- Becker, J.; Czamara, D.; Scerri, T.S.; Ramus, F.; Csépe, V.; Talcott, J.B.; Stein, J.; Morris, A.; Ludwig, K.U.; Hoffmann, P.; et al. Genetic analysis of dyslexia candidate genes in the European cross-linguistic NeuroDys cohort. Eur. J. Hum. Genet. 2014, 22, 675–680. [Google Scholar] [CrossRef] [Green Version]
- Doust, C.; Fontanillas, P.; Eising, E.; Gordon, S.D.; Wang, Z.; Alagöz, G.; Molz, B.; Me Research Team; Quantitative Trait Working Group of the GenLang Consortium; Pourcain, B.S.; et al. Discovery of 42 genome-wide significant loci associated with dyslexia. MedRxiv 2021. [Google Scholar] [CrossRef]
- Gialluisi, A.; Andlauer, T.F.M.; Mirza-Schreiber, N.; Moll, K.; Becker, J.; Hoffmann, P.; Ludwig, K.U.; Czamara, D.; Pourcain, B.S.; Honbolygó, F.; et al. Genome-wide association study reveals new insights into the heritability and genetic correlates of developmental dyslexia. Mol. Psychiatry 2021, 26, 3004–3017. [Google Scholar] [CrossRef]
- Uffelmann, E.; Huang, Q.Q.; Munung, N.S.; de Vries, J.; Okada, Y.; Martin, A.R.; Martin, H.C.; Lappalainen, T.; Posthuma, D. Genome-wide association studies. Nat. Rev. Methods Primers 2021, 1, 59. Available online: https://www.nature.com/articles/s43586-021-00056-9 (accessed on 20 December 2021). [CrossRef]
- Bycroft, C.; Freeman, C.; Petkova, D.; Band, G.; Elliott, L.T.; Sharp, K.; Motyer, A.; Vukcevic, D.; Delaneau, O.; O’Connell, J.; et al. The UK Biobank resource with deep phenotyping and genomic data. Nature 2018, 562, 203–209. [Google Scholar] [CrossRef] [Green Version]
- Meaburn, E.L.; Harlaar, N.; Craig, I.W.; Schalkwyk, L.C.; Plomin, R. Quantitative trait locus association scan of early reading disability and ability using pooled DNA and 100K SNP microarrays in a sample of 5760 children. Mol. Psychiatry 2008, 13, 729–740. [Google Scholar] [CrossRef] [Green Version]
- Davis, O.S.P.; The Wellcome Trust Case Control Consortium; Band, G.; Pirinen, M.; Haworth, C.; Meaburn, E.; Kovas, Y.; Harlaar, N.; Docherty, S.J.; Hanscombe, K.B.; et al. The correlation between reading and mathematics ability at age twelve has a substantial genetic component. Nat. Commun. 2014, 5, 4204. [Google Scholar] [CrossRef]
- Luciano, M.; Evans, D.; Hansell, N.; Medland, S.; Montgomery, G.; Martin, N.; Wright, M.; Bates, T.C. A genome-wide association study for reading and language abilities in two population cohorts. Genes Brain Behav. 2013, 12, 645–652. [Google Scholar] [CrossRef] [Green Version]
- Gialluisi, A.; Newbury, D.F.; Wilcutt, E.G.; Olson, R.K.; DeFries, J.C.; Brandler, W.M.; Pennington, B.F.; Smith, S.D.; Scerri, T.S.; Simpson, N.; et al. Genome-wide screening for DNA variants associated with reading and language traits. Genes Brain Behav. 2014, 13, 686–701. [Google Scholar] [CrossRef] [Green Version]
- Gialluisi, A.; Andlauer, T.F.M.; Mirza-Schreiber, N.; Moll, K.; Becker, J.; Hoffmann, P.; Ludwig, K.U.; Czamara, D.; Pourcain, B.S.; Brandler, W.; et al. Genome-wide association scan identifies new variants associated with a cognitive predictor of dyslexia. Transl. Psychiatry 2019, 9, 77. [Google Scholar] [CrossRef] [Green Version]
- Truong, D.T.; Adams, A.K.; Paniagua, S.; Frijters, J.C.; Boada, R.; Hill, D.E.; Lovett, M.W.; Mahone, E.M.; Willcutt, E.G.; Wolf, M.; et al. Multivariate genome-wide association study of rapid automatised naming and rapid alternating stimulus in Hispanic American and African-American youth. J. Med. Genet. 2019, 56, 557–566. [Google Scholar] [CrossRef]
- Eising, E.; Mirza-Schreiber, N.; de Zeeuw, E.L.; Wang, C.A.; Truong, D.T.; Allegrini, A.G.; Shapland, C.Y.; Zhu, G.; Wigg, K.G.; Gerritse, M.; et al. Genome-wide association analyses of individual differences in quantitatively assessed reading- and language-related skills in up to 34,000 people. BioRxiv 2021. [Google Scholar] [CrossRef]
- Little, C.W.; Hart, S.A. Examining the genetic and environmental associations among spelling, reading fluency, reading comprehension and a high stakes reading test in a combined sample of third and fourth grade students. Learn. Individ. Differ. 2016, 45, 25–32. [Google Scholar] [CrossRef] [Green Version]
- Wray, N.R.; Wijmenga, C.; Sullivan, P.F.; Yang, J.; Visscher, P.M. Common disease is more complex than implied by the core gene omnigenic model. Cell 2018, 173, 1573–1580. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.J.; Wedow, R.; Okbay, A.; Kong, E.; Maghzian, O.; Zacher, M.; Nguyen-Viet, T.A.; Bowers, P.; Sidorenko, J.; Linnér, R.K.; et al. Gene discovery and polygenic prediction from a genome-wide association study of educational attainment in 1.1 million individuals. Nat. Genet. 2018, 50, 1112–1121. [Google Scholar] [CrossRef] [Green Version]
- Selzam, S.; Dale, P.S.; Wagner, R.K.; DeFries, J.C.; Cederlöf, M.; O’Reilly, P.F.; Krapohl, E.; Plomin, R. Genome-wide polygenic scores predict reading performance throughout the school years. Sci. Stud. Read. 2017, 21, 334–349. [Google Scholar] [CrossRef] [Green Version]
- Price, K.M.; Wigg, K.G.; Feng, Y.; Blokland, K.; Wilkinson, M.; He, G.; Kerr, E.N.; Carter, T.-C.; Guger, S.L.; Lovett, M.W.; et al. Genome-wide association study of word reading: Overlap with risk genes for neurodevelopmental disorders. Genes Brain Behav. 2020, 19, 6. [Google Scholar] [CrossRef]
- Taipale, M.; Kaminen, N.; Nopola-Hemmi, J.; Haltia, T.; Myllyluoma, B.; Lyytinen, H.; Muller, K.; Kaaranen, M.; Lindsberg, P.J.; Hannula-Jouppi, K.; et al. A candidate gene for developmental dyslexia encodes a nuclear tetratricopeptid.de repeat domain protein dynamically regulated in brain. Proc. Natl. Acad. Sci. USA 2003, 100, 11553–11558. Available online: http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Citation&list_uids=12954984 (accessed on 20 December 2021). [CrossRef] [Green Version]
- Stefansson, H.; Meyer-Lindenberg, A.; Steinberg, S.; Magnusdottir, B.B.; Morgen, K.; Arnarsdottir, S.; Bjornsdottir, G.; Walters, G.B.; Jonsdottir, G.A.; Doyle, O.; et al. CNVs conferring risk of autism or schizophrenia affect cognition in controls. Nature 2014, 505, 361–366. [Google Scholar] [CrossRef] [Green Version]
- Ulfarsson, M.O.; Walters, G.B.; Gustafsson, O.; Steinberg, S.; Silva, A.; Doyle, O.M.; Brammer, M.; Gudbjartsson, D.F.; Arnarsdottir, S.; A Jonsdottir, G.; et al. 15q11.2 CNV affects cognitive, structural and functional correlates of dyslexia and dyscalculia. Transl. Psychiatry 2017, 7, e1109. [Google Scholar] [CrossRef] [Green Version]
- Pettigrew, K.; Reeves, E.; Leavett, R.; Hayiou-Thomas, E.; Sharma, A.; Simpson, N.H.; Martinelli, A.; Thompson, P.; Hulme, C.; Snowling, M.J.; et al. Copy number variation screen identifies a rare de novo deletion at chromosome 15q13.1-13.3 in a child with language impairment. PLoS ONE 2015, 10, e0134997. [Google Scholar] [CrossRef]
- Martinelli, A.; Rice, M.L.; Talcott, J.B.; Diaz, R.; Smith, S.; Raza, M.H.; Snowling, M.J.; Hulme, C.; Stein, J.; E Hayiou-Thomas, M.; et al. A rare missense variant in the ATP2C2 gene is associated with language impairment and related measures. Hum. Mol. Genet. 2021, 30, 1160–1171. [Google Scholar] [CrossRef]
- Grimm, T.; Garshasbi, M.; Puettmann, L.; Chen, W.; Ullmann, R.; Müller-Myhsok, B.; Klopocki, E.; Herbst, L.; Haug, J.; Jensen, L.R.; et al. A novel locus and candidate gene for familial developmental dyslexia on chromosome 4q. Z. Kinder Jug. Psych. 2020, 48, 478–489. [Google Scholar] [CrossRef]


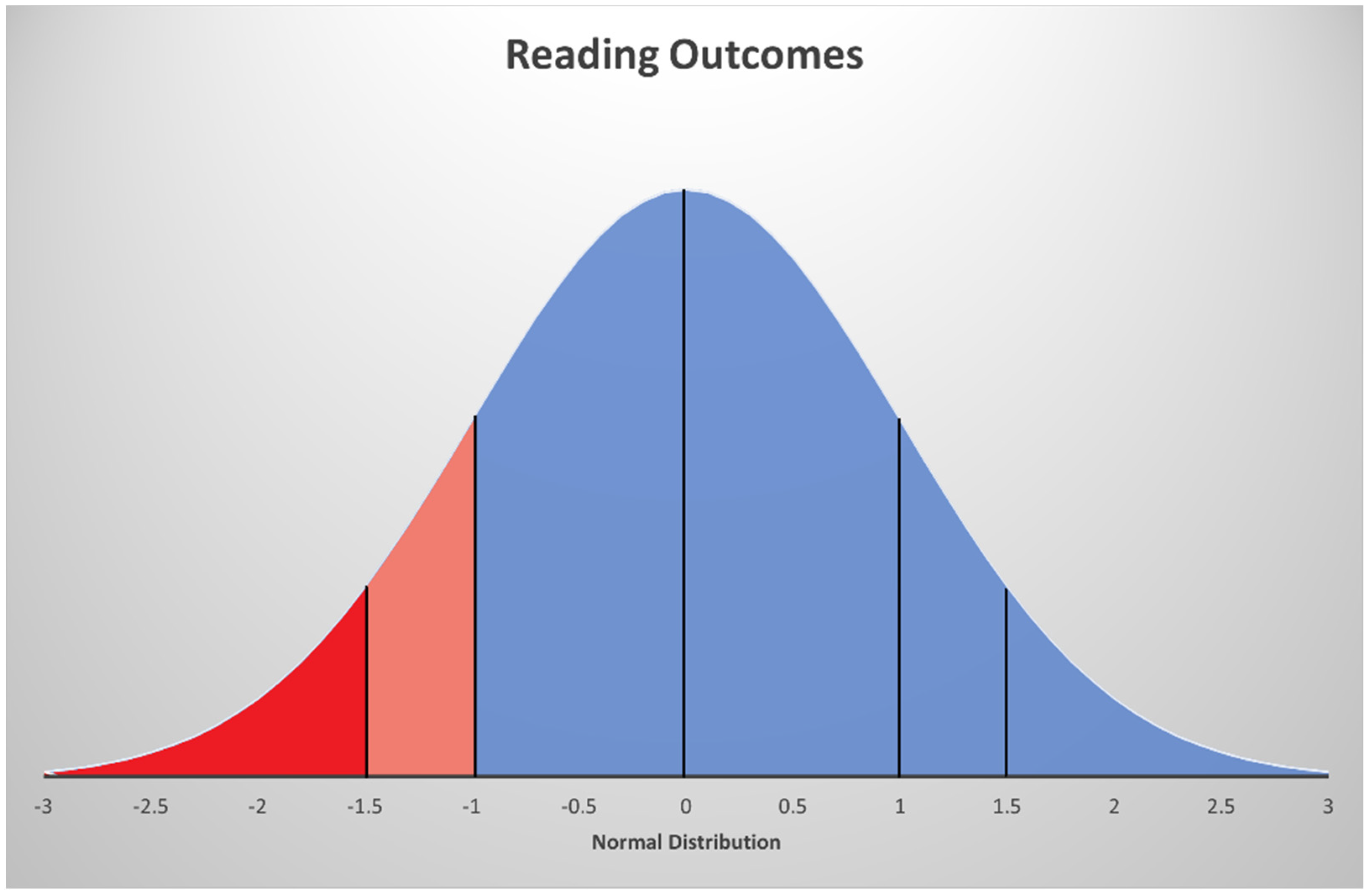{kind=link}
| Construct Measured | Description | Examples |
|---|---|---|
| Word Recognition | Ability to read single words or sight words | Wechsler Individual Achievement Test—Word Reading |
| Decoding | Ability to read unfamiliar words using letter–sound knowledge (often nonsense words) | Wechsler Individual Achievement Test—Decoding |
| Spelling | Ability to spell words from memory | Woodcock–Johnson Spelling |
| Phonological Processing | Ability to identify, pronounce, or recall individual sounds (phonemes) | Comprehensive Test of Phonological Processing |
| Fluency | Ability to read word lists or passages at an appropriate pace | Gray Oral Reading Test |
| Rapid Automatized Naming | Ability to quickly name a series of objects, colors, numbers, or letters | Comprehensive Test of Phonological Processing |
| Reading Comprehension | Ability to understand and answer questions about an independently read text | Woodcock–Johnson Reading Comprehension |
| Listening Comprehension or Oral Language | Ability to understand a story read-aloud or spoken directions | Wechsler Individual Achievement Test—Oral Language |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Erbeli, F.; Rice, M.; Paracchini, S. Insights into Dyslexia Genetics Research from the Last Two Decades. Brain Sci. 2022, 12, 27. https://doi.org/10.3390/brainsci12010027
Erbeli F, Rice M, Paracchini S. Insights into Dyslexia Genetics Research from the Last Two Decades. Brain Sciences. 2022; 12(1):27. https://doi.org/10.3390/brainsci12010027
Chicago/Turabian StyleErbeli, Florina, Marianne Rice, and Silvia Paracchini. 2022. "Insights into Dyslexia Genetics Research from the Last Two Decades" Brain Sciences 12, no. 1: 27. https://doi.org/10.3390/brainsci12010027