
Association between Novel Object Recognition/Spontaneous Alternation Behavior and Emission of Ultrasonic Vocalizations in Rats: Possible Relevance to the Study of Memory



Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects
2.2. Experimental Plan
2.3. Evaluation of Basal Calling Behavior
2.4. Tickling
2.5. Novel Object Recognition Test
2.6. Single-Trial Continuous Spontaneous Alternation Behavior Test in a Y Maze
2.7. Recording of Ultrasonic Vocalizations
2.8. Data Collection and Statistical Analyses
3. Results
3.1. Basal Levels of Calling Behavior
3.2. Emission of Ultrasonic Vocalizations in Response to Tickling
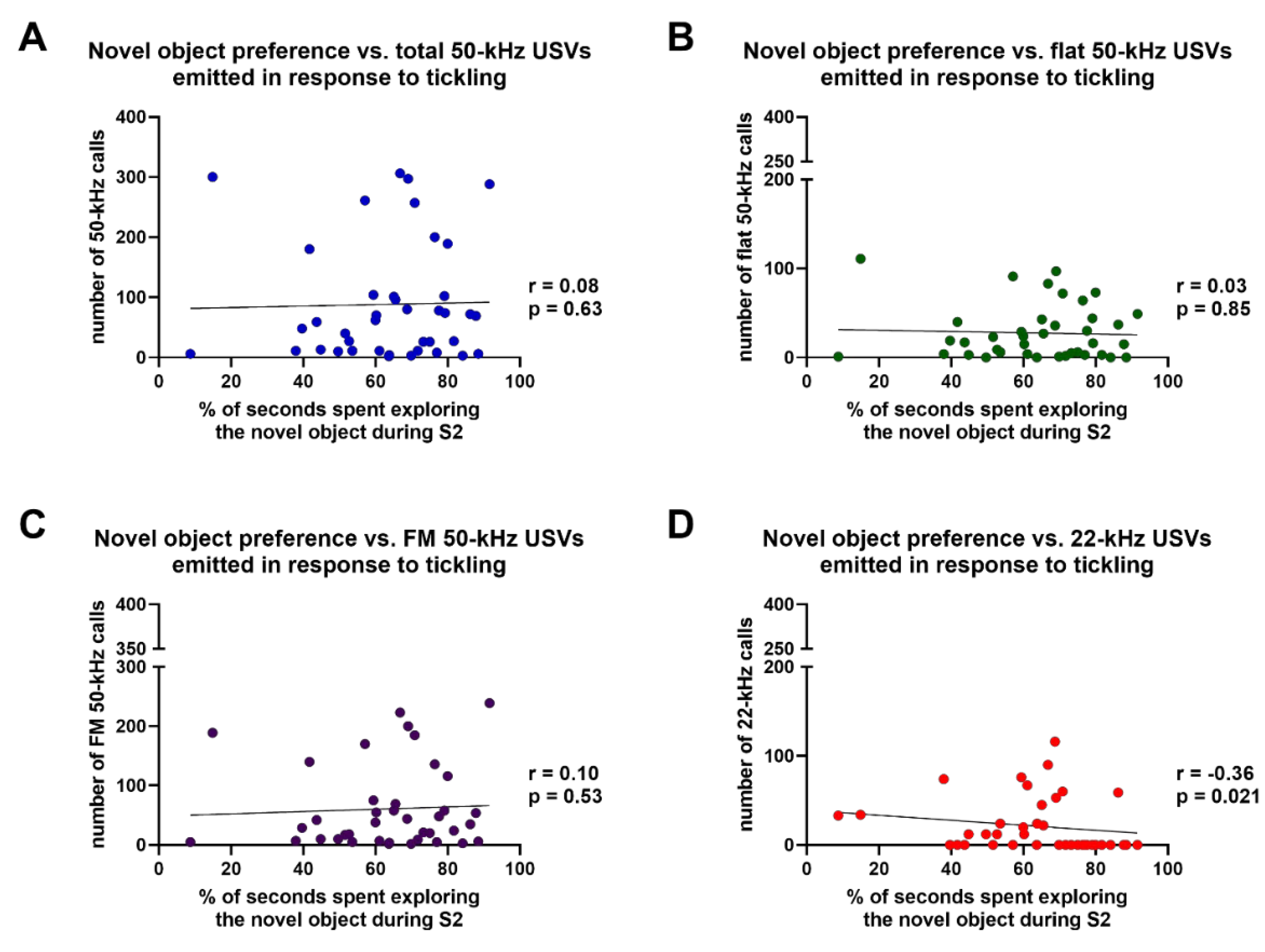
3.3. Novel Object Recognition Test and Emission of Ultrasonic Vocalizations
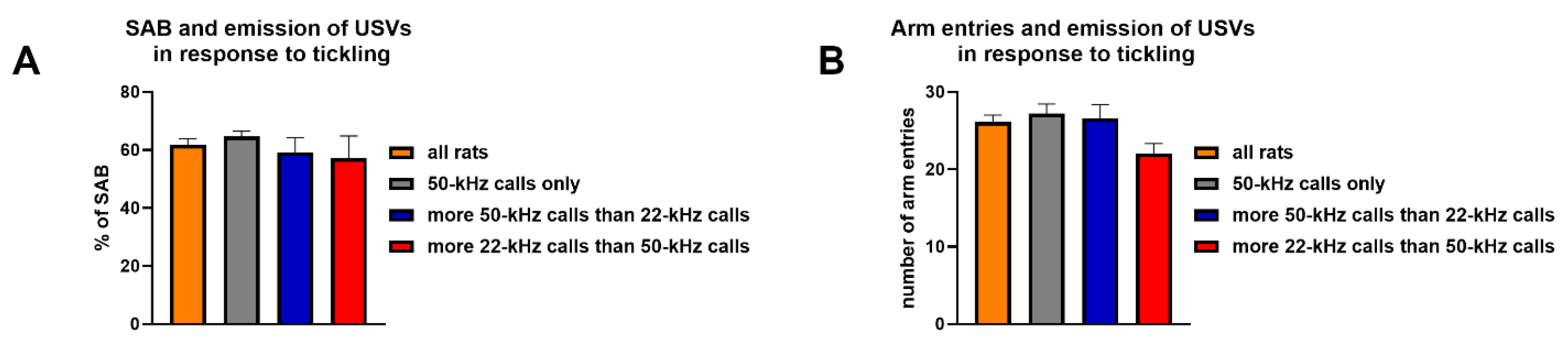
3.4. Single-Trial Continuous Spontaneous Alternation Behavior Test in a Y Maze and Emission of Ultrasonic Vocalizations
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Brudzynski, S.M. Ethotransmission: Communication of emotional states through ultrasonic vocalization in rats. Curr. Opin. Neurobiol. 2013, 23, 310–317. [Google Scholar] [CrossRef] [PubMed]
- Burgdorf, J.; Panksepp, J.; Moskal, J.R. Frequency-modulated 50kHz ultrasonic vocalizations: A tool for uncovering the molecular substrates of positive affect. Neurosci. Biobehav. Rev. 2011, 35, 1831–1836. [Google Scholar] [CrossRef]
- Wöhr, M.; Schwarting, R.K.W. Affective communication in rodents: Ultrasonic vocalizations as a tool for research on emotion and motivation. Cell Tissue Res. 2013, 354, 81–97. [Google Scholar] [CrossRef]
- Simola, N.; Brudzynski, S.M. Repertoire and Biological Function of Ultrasonic Vocalizations in Adolescent and Adult Rats. In Handbook of Ultrasonic Vocalization: A Window into the Emotional Brain; Elsevier Academic Press: Cambridge, MA, USA, 2018; Volume 25, pp. 177–186. [Google Scholar]
- Simola, N.; Brudzynski, S.M. Rat 50-kHz ultrasonic vocalizations as a tool in studying neurochemical mechanisms that regulate positive emotional states. J. Neurosci. Methods 2018, 310, 33–44. [Google Scholar] [CrossRef]
- Simola, N.; Granon, S. Ultrasonic vocalizations as a tool in studying emotional states in rodent models of social behavior and brain disease. Neuropharmacology 2019, 159, 107420. [Google Scholar] [CrossRef]
- Kassai, F.; Gyertyán, I. Effects of Selective Serotonin Reuptake Inhibitors on the Shock-Induced Ultrasonic Vocalization of Rats in Different Experimental Designs. In Handbook of Ultrasonic Vocalization: A Window into the Emotional Brain; Elsevier Academic Press: Cambridge, MA, USA, 2018; Volume 25, pp. 309–316. [Google Scholar]
- Sánchez, C. Stress-induced vocalisation in adult animals. A valid model of anxiety? Eur. J. Pharmacol. 2003, 463, 133–143. [Google Scholar] [CrossRef]
- Costa, G.; Serra, M.; Marongiu, J.; Morelli, M.; Simola, N. Influence of dopamine transmission in the medial prefrontal cortex and dorsal striatum on the emission of 50-kHz ultrasonic vocalizations in rats treated with amphetamine: Effects on drug-stimulated and conditioned calls. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2020, 97, 109797. [Google Scholar] [CrossRef]
- Ma, S.T.; Maier, E.Y.; Ahrens, A.M.; Schallert, T.; Duvauchelle, C.L. Repeated intravenous cocaine experience: Development and escalation of pre-drug anticipatory 50-kHz ultrasonic vocalizations in rats. Behav. Brain Res. 2010, 212, 109–114. [Google Scholar] [CrossRef] [Green Version]
- Hamed, A.; Kursa, M.B. Inter-individual differences in serotonin and glutamate co-transmission reflect differentiation in context-induced conditioned 50-kHz USVs response after morphine withdrawal. Brain Struct. Funct. 2018, 223, 3149–3167. [Google Scholar] [CrossRef] [Green Version]
- Panksepp, J.; Burgdorf, J. 50-kHz chirping (laughter?) in response to conditioned and unconditioned tickle-induced reward in rats: Effects of social housing and genetic variables. Behav. Brain Res. 2000, 115, 25–38. [Google Scholar] [CrossRef]
- Simola, N.; Frau, L.; Plumitallo, A.; Morelli, M. Direct and long-lasting effects elicited by repeated drug administration on 50-kHz ultrasonic vocalizations are regulated differently: Implications for the study of the affective properties of drugs of abuse. Int. J. Neuropsychopharmacol. 2014, 17, 429–441. [Google Scholar] [CrossRef] [Green Version]
- Simola, N.; Costa, G.; Morelli, M. Activation of adenosine A2A receptors suppresses the emission of pro-social and drug-stimulated 50-kHz ultrasonic vocalizations in rats: Possible relevance to reward and motivation. Psychopharmacology 2016, 233, 507–519. [Google Scholar] [CrossRef]
- Simola, N.; Serra, M.; Marongiu, J.; Costa, G.; Morelli, M. Increased emissions of 50-kHz ultrasonic vocalizations in hemiparkinsonian rats repeatedly treated with dopaminomimetic drugs: A potential preclinical model for studying the affective properties of dopamine replacement therapy in Parkinson’s disease. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2021, 108, 110184. [Google Scholar] [CrossRef]
- Simola, N.; Morelli, M. Repeated amphetamine administration and long-term effects on 50-kHz ultrasonic vocalizations: Possible relevance to the motivational and dopamine-stimulating properties of the drug. Eur. Neuropsychopharmacol. 2015, 25, 343–355. [Google Scholar] [CrossRef]
- Tripi, J.A.; Dent, M.L.; Meyer, P.J. Individual differences in food cue responsivity are associated with acute and repeated cocaine-induced vocalizations, but not cue-induced vocalizations. Psychopharmacology 2017, 234, 437–446. [Google Scholar] [CrossRef] [Green Version]
- Reyes, K.A.E.; Kudva, P.S.; Heckler, B.; Gonzalez, A.E.; Sorg, B.A. Rat ultrasonic vocalizations as an index of memory. Neurosci. Lett. 2021, 741, 135458. [Google Scholar] [CrossRef]
- Anacker, C.; Hen, R. Adult hippocampal neurogenesis and cognitive flexibility—linking memory and mood. Nat. Rev. Neurosci. 2017, 18, 335–346. [Google Scholar] [CrossRef] [PubMed]
- Panchal, P.; Kaltenboeck, A.; Harmer, C.J. Cognitive emotional processing across mood disorders. CNS Spectr. 2019, 24, 54–63. [Google Scholar] [CrossRef] [Green Version]
- Ennaceur, A.; Delacour, J. A new one-trial test for neurobiological studies of memory in rats. 1: Behavioral data. Behav. Brain Res. 1988, 31, 47–59. [Google Scholar] [CrossRef]
- Hughes, R.N. The value of spontaneous alternation behavior (SAB) as a test of retention in pharmacological investigations of memory. Neurosci. Biobehav. Rev. 2004, 28, 497–505. [Google Scholar] [CrossRef]
- Burgdorf, J.; Panksepp, J. Tickling induces reward in adolescent rats. Physiol. Behav. 2001, 72, 167–173. [Google Scholar] [CrossRef]
- Schwarting, R.K.W. Ultrasonic vocalization in juvenile and adult male rats: A comparison among stocks. Physiol. Behav. 2018, 191, 1–11. [Google Scholar] [CrossRef]
- Cadoni, C.; Pisanu, A.; Simola, N.; Frau, L.; Porceddu, P.F.; Corongiu, S.; Dessì, C.; Sil, A.; Plumitallo, A.; Wardas, J.; et al. Widespread reduction of dopamine cell bodies and terminals in adult rats exposed to a low dose regimen of MDMA during adolescence. Neuropharmacology 2017, 123, 385–394. [Google Scholar] [CrossRef] [PubMed]
- Costa, G.; Morelli, M.; Simola, N. Involvement of glutamate NMDA receptors in the acute, long-term, and conditioned effects of amphetamine on rat 50 kHz ultrasonic vocalizations. Int. J. Neuropsychopharmacol. 2015, 18, pyv057. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simola, N.; Bustamante, D.; Pinna, A.; Pontis, S.; Morales, P.; Morelli, M.; Herrera-Marschitz, M. Acute perinatal asphyxia impairs non-spatial memory and alters motor coordination in adult male rats. Exp. Brain Res. 2008, 185, 595–601. [Google Scholar] [CrossRef] [PubMed]
- Simola, N.; Fenu, S.; Costa, G.; Pinna, A.; Plumitallo, A.; Morelli, M. Pharmacological characterization of 50-kHz ultrasonic vocalizations in rats: Comparison of the effects of different psychoactive drugs and relevance in drug-induced reward. Neuropharmacology 2012, 63, 224–234. [Google Scholar] [CrossRef]
- Wright, J.M.; Gourdon, J.C.; Clarke, P.B.S. Identification of multiple call categories within the rich repertoire of adult rat 50-kHz ultrasonic vocalizations: Effects of amphetamine and social context. Psychopharmacology 2010, 211, 1–13. [Google Scholar] [CrossRef]
- Burgdorf, J.; Panksepp, J.; Brudzynski, S.M.; Beinfeld, M.C.; Cromwell, H.C.; Kroes, R.A.; Moskal, J.R. The effects of selective breeding for differential rates of 50-kHz ultrasonic vocalizations on emotional behavior in rats. Dev. Psychobiol. 2009, 51, 34–46. [Google Scholar] [CrossRef]
- Webber, E.S.; Harmon, K.M.; Beckwith, T.J.; Peña, S.; Burgdorf, J.; Panksepp, J.; Cromwell, H.C. Selective breeding for 50kHz ultrasonic vocalization emission produces alterations in the ontogeny and regulation of rough-and-tumble play. Behav. Brain Res. 2012, 229, 138–144. [Google Scholar] [CrossRef]
- Bihari, A.; Hrycyshyn, A.W.; Brudzynski, S.M. Role of the mesolimbic cholinergic projection to the septum in the production of 22 kHz alarm calls in rats. Brain Res. Bull. 2003, 60, 263–274. [Google Scholar] [CrossRef]
- Silkstone, M.; Brudzynski, S.M. Dissimilar interaction between dopaminergic and cholinergic systems in the initiation of emission of 50-kHz and 22-kHz vocalizations. Pharmacol. Biochem. Behav. 2020, 188, 172815. [Google Scholar] [CrossRef]
- Nanfaro, F.; Cabrera, R.; Bazzocchini, V.; Laconi, M.; Yunes, R. Pregnenolone sulfate infused in lateral septum of male rats impairs novel object recognition memory. Pharmacol. Rep. 2010, 62, 265–272. [Google Scholar] [CrossRef]
- Chao, O.Y.; Huston, J.P.; Li, J.S.; Wang, A.L.; de Souza Silva, M.A. The medial prefrontal cortex-lateral entorhinal cortex circuit is essential for episodic-like memory and associative object-recognition. Hippocampus 2016, 26, 633–645. [Google Scholar] [CrossRef]
- Nelson, A.J.D.; Thur, K.E.; Marsden, C.A.; Cassaday, H.J. Dissociable Roles of Dopamine Within the Core and Medial Shell of the Nucleus Accumbens in Memory for Objects and Place. Behav. Neurosci. 2010, 124, 789–799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maurice, T.; Su, T.P.; Privat, A. Sigma1 (σ1) receptor agonists and neurosteroids attenuate b25-35-amyloid peptide-induced amnesia in mice through a common mechanism. Neuroscience 1998, 83, 413–428. [Google Scholar] [CrossRef]
- Meunier, J.; Gué, M.; Récasens, M.; Maurice, T. Attenuation by a sigma 1 (σ 1) receptor agonist of the learning and memory deficits induced by a prenatal restraint stress in juvenile rats. Br. J. Pharmacol. 2004, 142, 689–700. [Google Scholar] [CrossRef] [Green Version]
- Knutson, B.; Burgdorf, J.; Panksepp, J. Ultrasonic vocalizations as indices of affective states in rats. Psychol. Bull. 2002, 128, 961–977. [Google Scholar] [CrossRef]
- Burgdorf, J.; Panksepp, J. The neurobiology of positive emotions. Neurosci. Biobehav. Rev. 2006, 30, 173–187. [Google Scholar] [CrossRef]
- Berlyne, D.E. The arousal and satiation of perceptual curiosity in the rat. J. Comp. Physiol. Psychol. 1955, 48, 238–246. [Google Scholar] [CrossRef] [PubMed]
- Coura, R.S.; Cressant, A.; Xia, J.; De Chaumont, F.; Olivo-Marin, J.C.; Pelloux, Y.; Dalley, J.W.; Granon, S. Nonaggressive and adapted social cognition is controlled by the interplay between noradrenergic and nicotinic receptor mechanisms in the prefrontal cortex. FASEB J. 2013, 27, 4343–4354. [Google Scholar] [CrossRef] [Green Version]
- Wöhr, M.; Houx, B.; Schwarting, R.K.W.; Spruijt, B. Effects of experience and context on 50-kHz vocalizations in rats. Physiol. Behav. 2008, 93, 766–776. [Google Scholar] [CrossRef] [PubMed]
- Simola, N.; Costa, G. Emission of categorized 50-kHz ultrasonic vocalizations in rats repeatedly treated with amphetamine or apomorphine: Possible relevance to drug-induced modifications in the emotional state. Behav. Brain Res. 2018, 347, 88–98. [Google Scholar] [CrossRef]
- Garcia, E.J.; McCowan, T.J.; Cain, M.E. Harmonic and frequency modulated ultrasonic vocalizations reveal differences in conditioned and unconditioned reward processing. Behav. Brain Res. 2015, 287, 207–214. [Google Scholar] [CrossRef]
- Schwarting, R.K.W.; Jegan, N.; Wöhr, M. Situational factors, conditions and individual variables which can determine ultrasonic vocalizations in male adult Wistar rats. Behav. Brain Res. 2007, 182, 208–222. [Google Scholar] [CrossRef] [PubMed]
- Natusch, C.; Schwarting, R.K.W. Using bedding in a test environment critically affects 50-kHz ultrasonic vocalizations in laboratory rats. Pharmacol. Biochem. Behav. 2010, 96, 251–259. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cumulative Time of Object Exploration during S1 | Cumulative Time of Object Exploration during S2 | |
|---|---|---|
| all rats | 13.59 ± 1.29 | 15.33 ± 1.21 |
| 50-kHz calls only | 13.21 ± 1.43 | 14.99 ± 1.38 |
| more 50-kHz calls than 22-kHz calls | 12.27 ± 1.96 | 14.75 ± 2.26 |
| more 22-kHz calls than 50-kHz calls | 16.99 ± 5.15 | 17.34 ± 4.19 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Costa, G.; Serra, M.; Simola, N. Association between Novel Object Recognition/Spontaneous Alternation Behavior and Emission of Ultrasonic Vocalizations in Rats: Possible Relevance to the Study of Memory. Brain Sci. 2021, 11, 1053. https://doi.org/10.3390/brainsci11081053
Costa G, Serra M, Simola N. Association between Novel Object Recognition/Spontaneous Alternation Behavior and Emission of Ultrasonic Vocalizations in Rats: Possible Relevance to the Study of Memory. Brain Sciences. 2021; 11(8):1053. https://doi.org/10.3390/brainsci11081053
Chicago/Turabian StyleCosta, Giulia, Marcello Serra, and Nicola Simola. 2021. "Association between Novel Object Recognition/Spontaneous Alternation Behavior and Emission of Ultrasonic Vocalizations in Rats: Possible Relevance to the Study of Memory" Brain Sciences 11, no. 8: 1053. https://doi.org/10.3390/brainsci11081053