
Effects of Neurofeedback on the Working Memory of Children with Learning Disorders—An EEG Power-Spectrum Analysis

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics
2.2. Participants
2.3. The z-Value of the Theta/Alpha Ratio
2.4. Neurofeedback and Sham Treatments
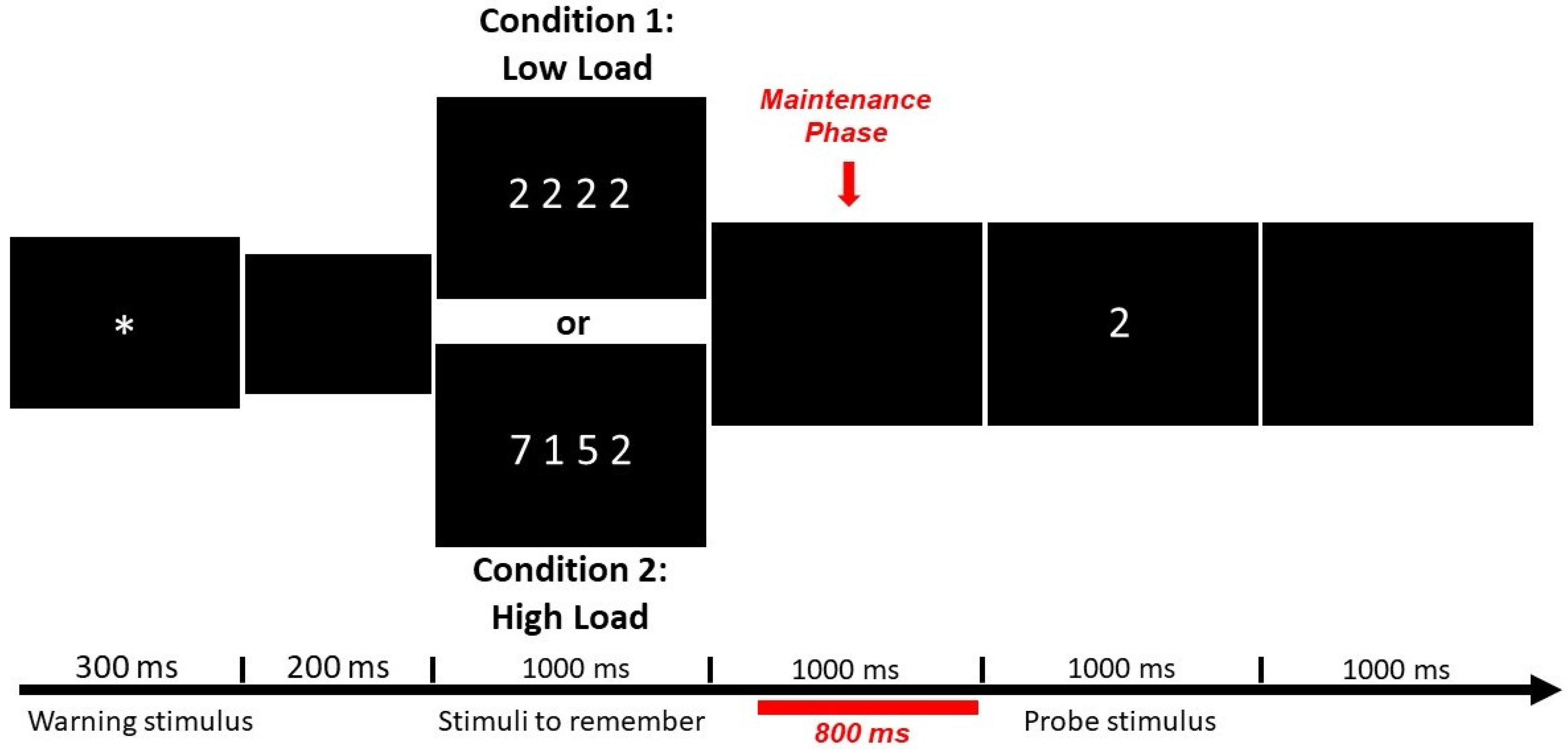
2.5. Working Memory Task
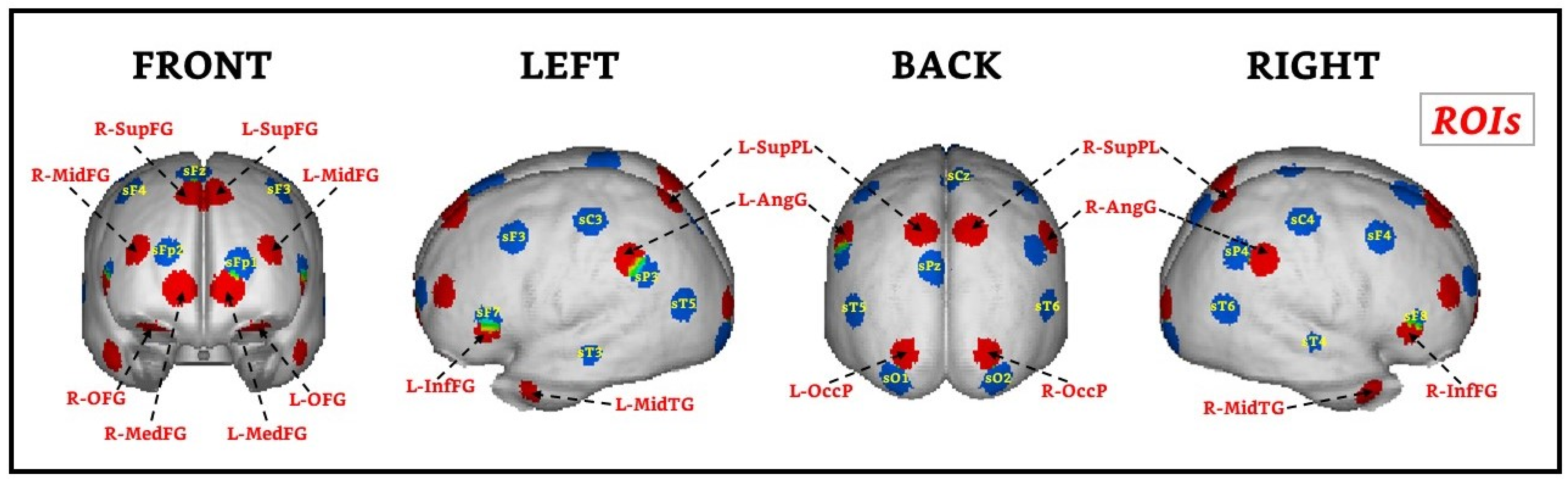
2.6. EEG Recording and Data Analysis of the WM Task
3. Results
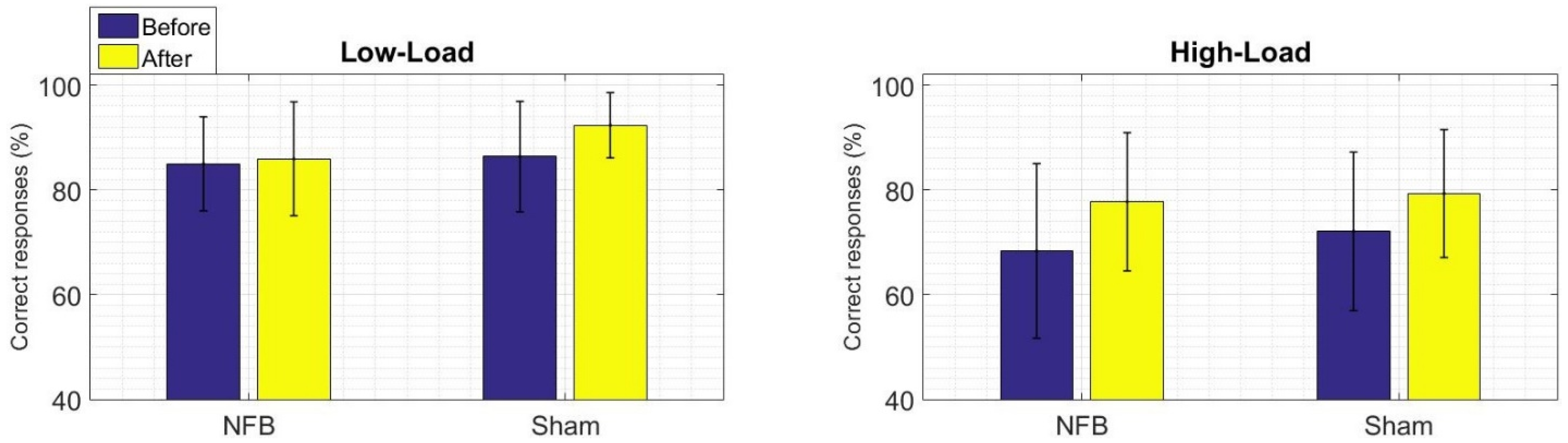
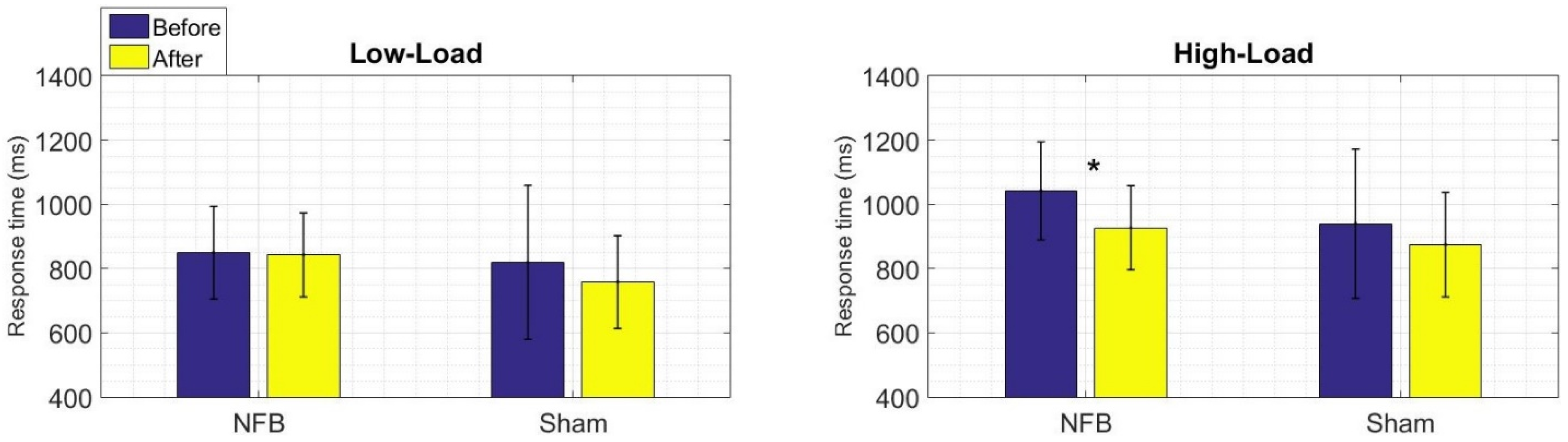
3.1. Behavioral Results of the WM Task
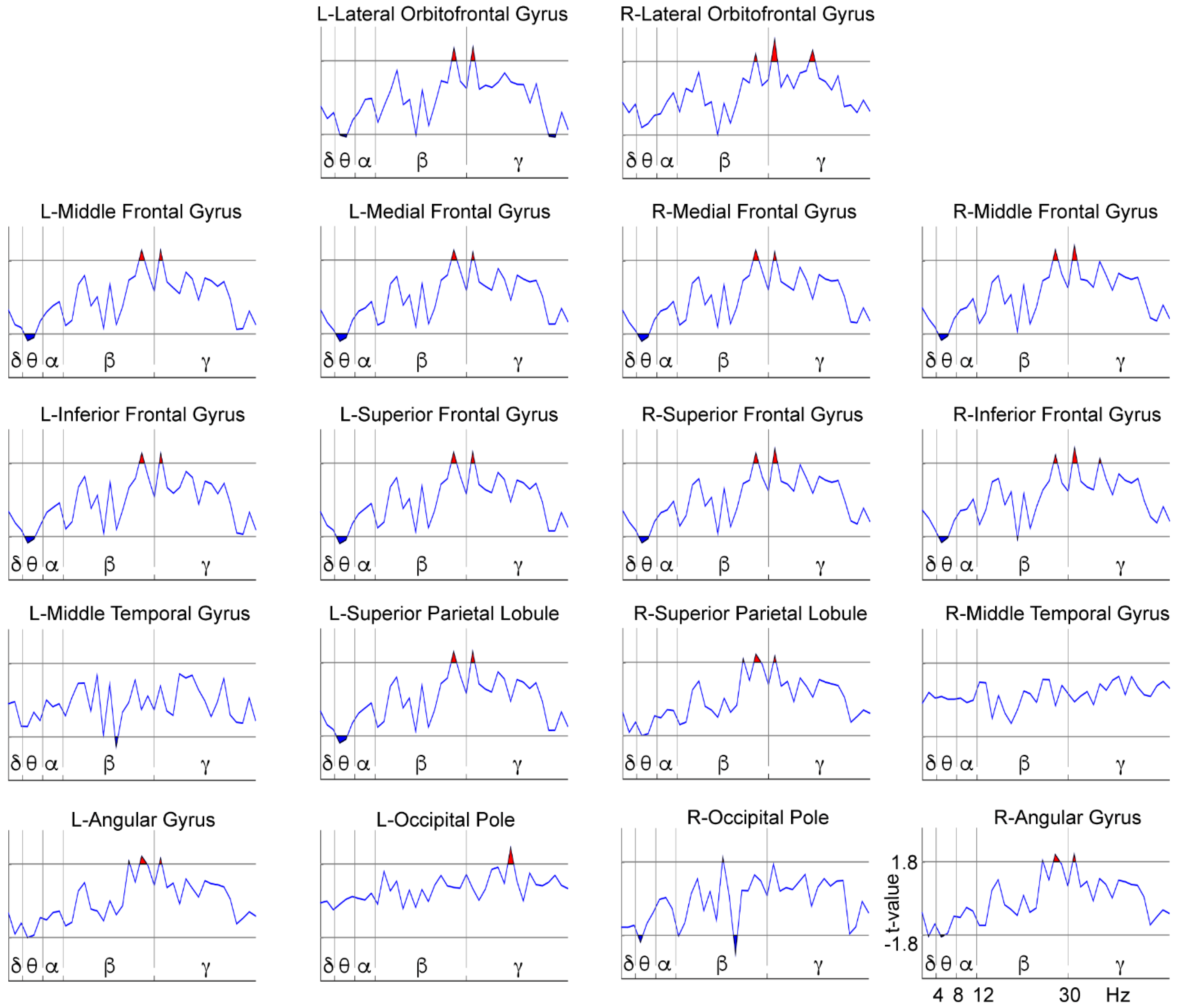
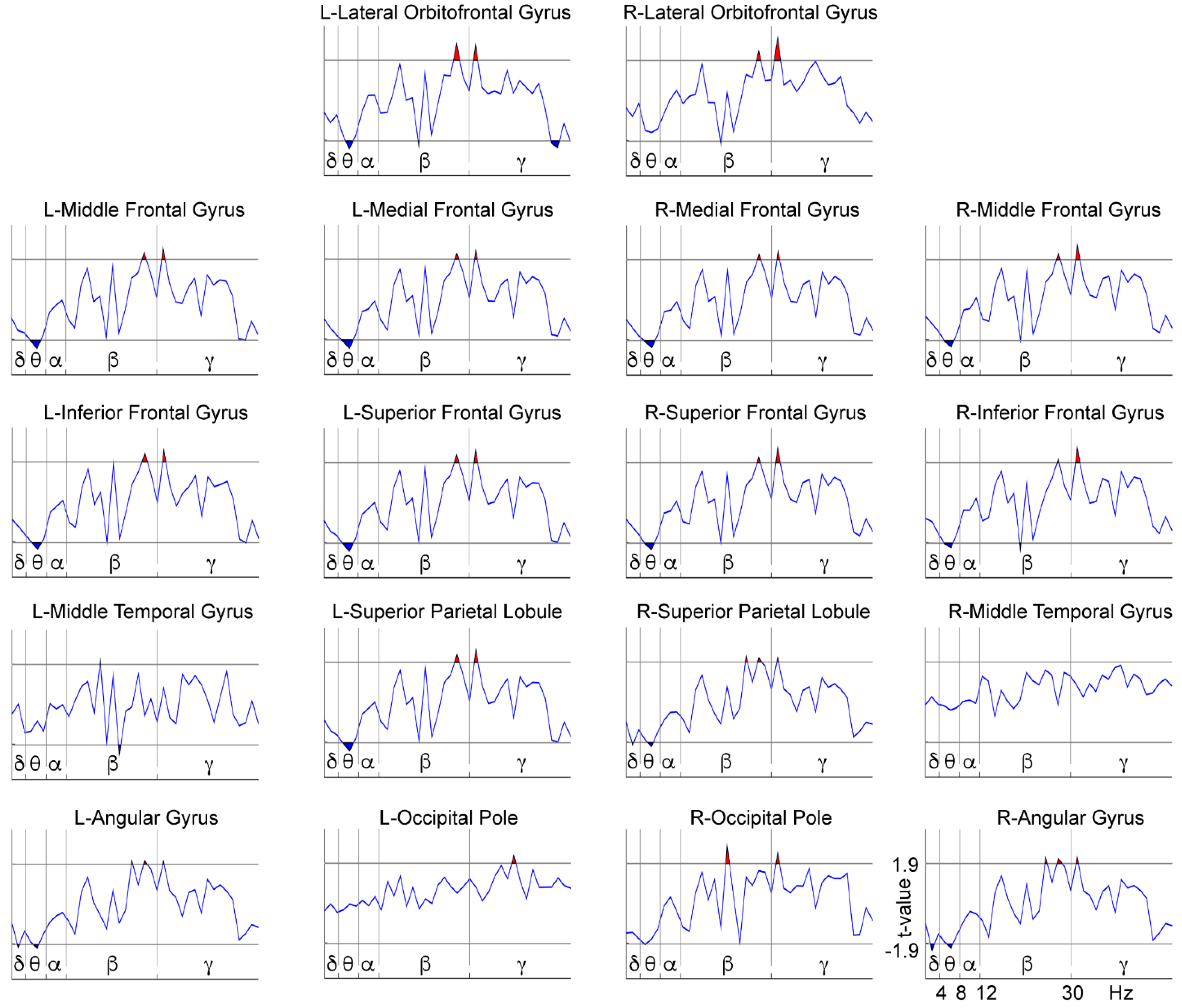
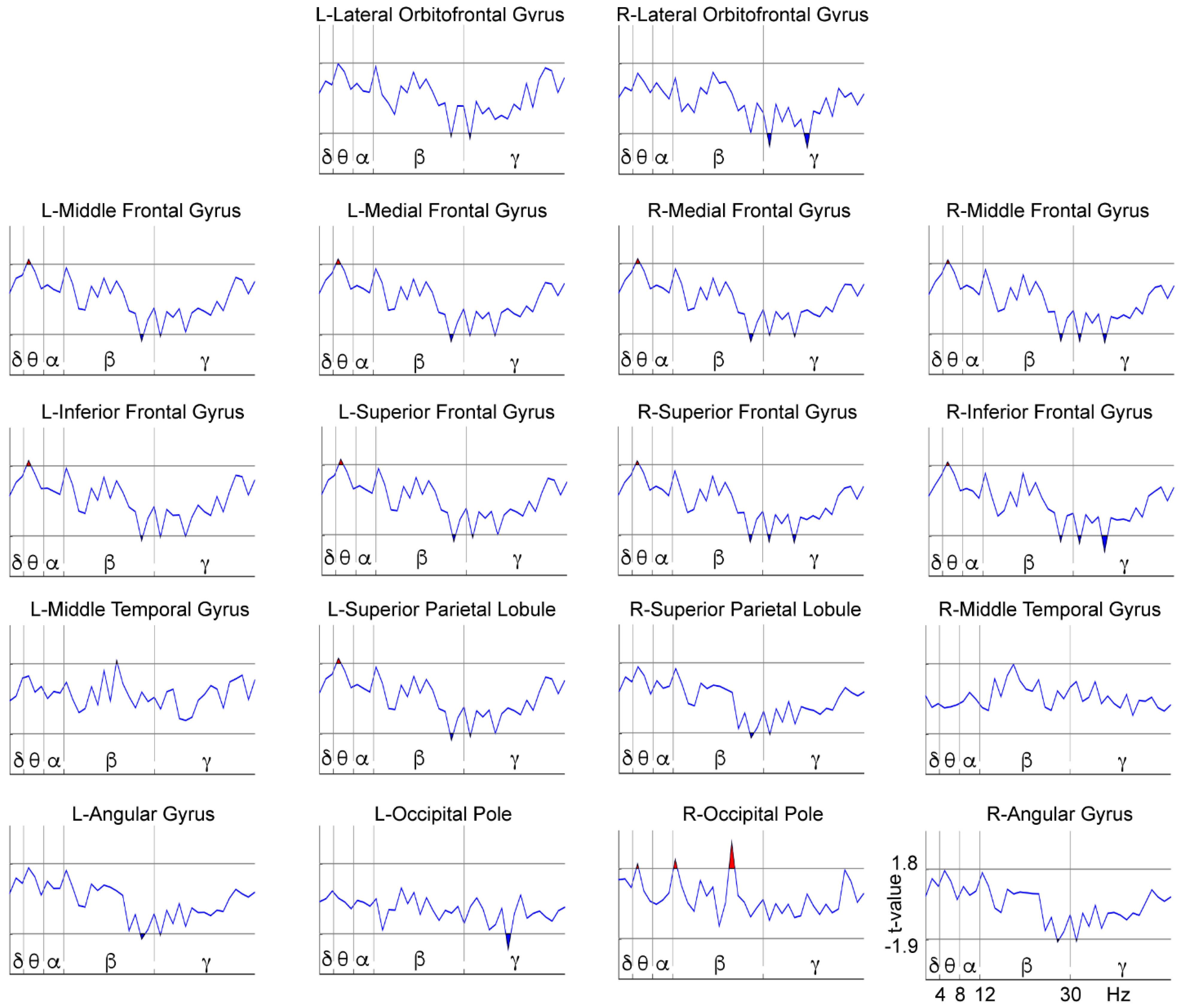
3.2. WM-Related Power Spectrum Results
4. Discussion
5. Limitations
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Altarac, M.; Saroha, E. Lifetime Prevalence of Learning Disability Among US Children. Pediatrics 2007, 119, S77. [Google Scholar] [CrossRef] [Green Version]
- American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders, 5th ed.; American Psychiatric Association: Arlington, VA, USA, 2013; ISBN 089042554X. [Google Scholar]
- Lagae, L. Learning Disabilities: Definitions, Epidemiology, Diagnosis, and Intervention Strategies. Pediatr. Clin. N. Am. 2008. [Google Scholar] [CrossRef]
- Shaywitz, S.E.; Fletcher, J.M.; Holahan, J.M.; Shneider, A.E.; Marchione, K.E.; Stuebing, K.K.; Francis, D.J.; Pugh, K.R.; Shaywitz, B.A. Persistence of dyslexia: The Connecticut Longitudinal Study at adolescence. Pediatrics 1999, 104. [Google Scholar] [CrossRef] [Green Version]
- American Psychiatric Association [APA]. Diagnostic and Statistical Manual of Mental Disorders, 4th ed.; Text Revision (DSM-IV-TR); American Psychiatric Association: Washington, DC, USA, 2000; Volume 1, p. 943. [Google Scholar] [CrossRef]
- Willcutt, E.G.; Petrill, S.A.; Wu, S.; Boada, R.; DeFries, J.C.; Olson, R.K.; Pennington, B.F. Comorbidity Between Reading Disability and Math Disability: Concurrent Psychopathology, Functional Impairment, and Neuropsychological Functioning. J. Learn. Disabil. 2013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schuchardt, K.; Maehler, C.; Hasselhorn, M. Working memory deficits in children with specific learning disorders. J. Learn. Disabil. 2008, 41, 514–523. [Google Scholar] [CrossRef]
- Baddeley, A.D.; Hitch, G. Working Memory. Psychol. Learn. Motiv. 1974. [Google Scholar] [CrossRef]
- De Weerdt, F.; Desoete, A.; Roeyers, H. Working memory in children with reading disabilities and/or mathematical disabilities. J. Learn. Disabil. 2013, 46, 461–472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siegel, L.S.; Ryan, E.B. The development of working memory in normally achieving and subtypes of learning disabled children. Child Dev. 1989, 60, 973–980. [Google Scholar] [CrossRef] [PubMed]
- Swanson, H.L. Intelligence, Working Memory, and Learning Disabilities; Elsevier Inc.: Amsterdam, The Netherlands, 2015; ISBN 9780124104440. [Google Scholar]
- Schuchardt, K.; Bockmann, A.-K.; Bornemann, G.; Maehler, C. Working Memory Functioning in Children With Learning Disorders and Specific Language Impairment. Top. Lang. Disord. 2013, 33, 298–312. [Google Scholar] [CrossRef]
- Alloway, T.P. Working memory, but not IQ, predicts subsequent learning in children with learning difficulties. Eur. J. Psychol. Assess. 2009, 25, 92–98. [Google Scholar] [CrossRef]
- Peng, P.; Barnes, M.; Wang, C.C.; Wang, W.; Li, S.; Swanson, H.L.; Dardick, W.; Tao, S. Meta-analysis on the relation between reading and working memory. Psychol. Bull. 2018, 144. [Google Scholar] [CrossRef] [PubMed]
- Stanley, P.D.; Dai, Y.; Nolan, R.F. Differences in depression and self-esteem reported by learning disabled and behavior disordered middle school students. J. Adolesc. 1997, 20. [Google Scholar] [CrossRef]
- Francis, D.A.; Caruana, N.; Hudson, J.L.; McArthur, G.M. The association between poor reading and internalising problems: A systematic review and meta-analysis. Clin. Psychol. Rev. 2019, 67. [Google Scholar] [CrossRef] [PubMed]
- Nelson, J.M.; Gregg, N. Depression and anxiety among transitioning adolescents and college students with ADHD, dyslexia, or comorbid ADHD/dyslexia. J. Atten. Disord. 2012, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paszkiel, S. Analysis and Classification of EEG Signals for Brain–Computer Interfaces; Springer: Cham, Switzerland, 2020; Volume 852, ISBN 978-3-030-30581-9. [Google Scholar]
- Roca-Stappung, M.; Fernández, T.; Bosch-Bayard, J.; Harmony, T.; Ricardo-Garcell, J. Electroencephalographic characterization of subgroups of children with learning disorders. PLoS ONE 2017, 12, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernández, T.; Harmony, T.; Fernández-Bouzas, A.; Silva, J.; Herrera, W.; Santiago-Rodríguez, E.; Sánchez, L. Sources of EEG activity in learning disabled children. Clin. EEG Electroencephalogr. 2002, 33, 160–164. [Google Scholar] [CrossRef]
- Fonseca, L.C.; Tedrus, G.M.A.S.; Chiodi, M.G.; Cerqueira, J.N.; Tonelotto, J.M.F. Quantitative EEG in children with learning disabilities: Analysis of band power. Arq. Neuropsiquiatr. 2006. [Google Scholar] [CrossRef] [Green Version]
- Chabot, R.J.; di Michele, F.; Prichep, L.; John, E.R. The clinical role of computerized EEG in the evaluation and treatment of learning and attention disorders in children and adolescents. J. Neuropsychiatry Clin. Neurosci. 2001, 13, 171–186. [Google Scholar] [CrossRef]
- Sternberg, S. Memory-scanning: Mental processes revealed by reaction-time experiments. Am. Sci. 1969. [Google Scholar] [CrossRef]
- Silva-Pereyra, J.; Fernández, T.; Harmony, T.; Bernal, J.; Galán, L.; Díaz-Comas, L.; Fernández-Bouzas, A.; Yáñez, G.; Rivera-Gaxiola, M.; Rodríguez, M.; et al. Delayed P300 during Sternberg and color discrimination tasks in poor readers. Int. J. Psychophysiol. 2001, 40, 17–32. [Google Scholar] [CrossRef]
- Luck, S.J.; Kappenman, E.S. Electroencephalography and Event-Related Brain Potentials. In Handbook of Psychophysiology; Cacioppo, J.T., Tassinary, L.G., Berntson, G.G., Eds.; Cambridge University Press: Cambridge, UK, 2017; pp. 74–100. [Google Scholar]
- Fernandez, T.; Harmony, T.; Gersenowies, J.; Silva-Pereyra, J.; Fernández-Bouzas, A.; Galán, L.; Díaz-Comas, L. Chapter 41 Sources of EEG activity during a verbal working memory task in adults and children. Suppl. Clin. Neurophysiol. 2002. [Google Scholar] [CrossRef]
- Martínez-Briones, B.J.; Fernández-Harmony, T.; Gómez, N.G.; Biscay-Lirio, R.J.; Bosch-Bayard, J. Working memory in children with learning disorders: An EEG power spectrum analysis. Brain Sci. 2020, 10, 817. [Google Scholar] [CrossRef]
- Schapkin, S.A.; Raggatz, J.; Hillmert, M.; Böckelmann, I. EEG correlates of cognitive load in a multiple choice reaction task. Acta Neurobiol. Exp. 2020, 80, 76–89. [Google Scholar] [CrossRef] [Green Version]
- Dimitriadis, S.I.; Laskaris, N.A.; Tsirka, V.; Vourkas, M.; Micheloyannis, S. What does delta band tell us about cognitive processes: A mental calculation study. Neurosci. Lett. 2010, 483. [Google Scholar] [CrossRef] [PubMed]
- Harmony, T. The functional significance of delta oscillations in cognitive processing. Front. Integr. Neurosci. 2013, 7. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, D.J.; Mcnaughton, N.; Flanagan, D.; Kirk, I.J. Progress in Neurobiology Frontal-midline theta from the perspective of hippocampal “theta”. Prog. Neurobiol. 2008, 86, 156–185. [Google Scholar] [CrossRef] [PubMed]
- Eschmann, K.C.J.; Bader, R.; Mecklinger, A. Topographical differences of frontal-midline theta activity reflect functional differences in cognitive control abilities. Brain Cogn. 2018, 123, 57–64. [Google Scholar] [CrossRef]
- Cavanagh, J.F.; Frank, M.J. Frontal theta as a mechanism for cognitive control. Trends Cogn. Sci. 2014, 18, 414–421. [Google Scholar] [CrossRef] [Green Version]
- Gevins, A.; Smith, M.E.; McEvoy, L.; Yu, D.; Gevins, A.; Smith, M.E.; McEvoy, L.; Yu, D. High-resolution EEG mapping of cortical activation related to working memory: Effects of task difficulty, type of processing, and practice. Cereb. Cortex 1997, 7, 374–385. [Google Scholar] [CrossRef] [Green Version]
- Maurer, U.; Brem, S.; Liechti, M. Frontal Midline Theta Reflects Individual Task Performance in a Working Memory Task. Brain Topogr. 2015, 127–134. [Google Scholar] [CrossRef] [Green Version]
- Fuentes-García, J.P.; Villafaina, S.; Collado-Mateo, D.; Cano-Plasencia, R.; Gusi, N. Chess players increase the theta power spectrum when the difficulty of the opponent increases: An EEG study. Int. J. Environ. Res. Public Health 2020, 17, 46. [Google Scholar] [CrossRef] [Green Version]
- Jokisch, D.; Jensen, O. Modulation of gamma and alpha activity during a working memory task engaging the dorsal or ventral stream. J. Neurosci. 2007, 27, 3244–3251. [Google Scholar] [CrossRef] [Green Version]
- Jensen, O.; Kaiser, J.; Lachaux, J.P. Human gamma-frequency oscillations associated with attention and memory. Trends Neurosci. 2007, 30, 317–324. [Google Scholar] [CrossRef]
- Chen, Y.; Huang, X.; Spironelli, C.; Penolazzi, B.; Vio, C.; Angrilli, A.; Maurer, U.; Brem, S.; Liechti, M.; Maurizio, S.; et al. FFT transformed quantitative EEG analysis of short term memory load. J. Neurosci. 2017, 6, 3788–3801. [Google Scholar] [CrossRef]
- Honkanen, R.; Rouhinen, S.; Wang, S.H.; Palva, J.M.; Palva, S. Gamma oscillations underlie the maintenance of feature-specific information and the contents of visual working memory. Cereb. Cortex 2015, 25, 3788–3801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tallon-Baudry, C.; Bertrand, O.; Peronnet, F.; Pernier, J. Induced γ-band activity during the delay of a visual short-term memory task in humans. J. Neurosci. 1998, 18, 4244–4254. [Google Scholar] [CrossRef]
- Shaywitz, S.; Shaywitz, J. Overcoming Dyslexia: Completely Revised and Updated, 2nd ed.; Knopf, A.A., Ed.; Penguin Random House LLC: New York, NY, USA, 2020. [Google Scholar]
- Swanson, H.L.; Hoskyn, M. Experimental intervention research on students with learning disabilities: A meta-analysis of treatment outcomes. Rev. Educ. Res. 1998, 68. [Google Scholar] [CrossRef]
- NRP Report of the National Reading Panel—Teaching Children to Read: An Evidence-Based Assessment of the Scientific Research Literature on Reading and Its Implications for Reading Instruction; No. 00-4769; NIH: Bethesda, MD, USA, 2000; Volume 7.
- Budzynski, T.; Budzynski, H.; Evans, J.; Abarbanel, A. Introduction to Quantitative EEG and Neurofeedback; Academic Press: Burlington, MA, USA, 2009. [Google Scholar]
- Gruzelier, J.H. Neuroscience and Biobehavioral Reviews EEG-neurofeedback for optimising performance. I: A review of cognitive and affective outcome in healthy participants. Neurosci. Biobehav. Rev. 2014, 44, 124–141. [Google Scholar] [CrossRef] [PubMed]
- Sitaram, R.; Ros, T.; Stoeckel, L.; Haller, S.; Scharnowski, F.; Lewis-Peacock, J.; Weiskopf, N.; Blefari, M.L.; Rana, M.; Oblak, E.; et al. Closed-loop brain training: The science of neurofeedback. Nat. Rev. Neurosci. 2017, 18, 86–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thibault, R.T.; Raz, A. When can neurofeedback join the clinical armamentarium? Lancet Psychiatry 2016, 3, 497–498. [Google Scholar] [CrossRef] [Green Version]
- Nazari, M.A.; Mosanezhad, E.; Hashemi, T.; Jahan, A. The Effectiveness of Neurofeedback Training on EEG Coherence and Neuropsychological Functions in Children With Reading Disability. Clin. EEG Neurosci. 2012, 1–8. [Google Scholar] [CrossRef]
- Breteler, M.H.M.; Arns, M.; Peters, S.; Giepmans, I.; Verhoeven, L. Improvements in spelling after QEEG-based neurofeedback in dyslexia: A randomized controlled treatment study. Appl. Psychophysiol. Biofeedback 2010, 35, 5–11. [Google Scholar] [CrossRef] [Green Version]
- Fernández, T.; Herrera, W.; Harmony, T.; Díaz-Comas, L.; Santiago, E.; Sánchez, L.; Bosch, J.; Fernández-Bouzas, A.; Otero, G.; Ricardo-Garcell, J.; et al. EEG and Behavioral Changes following Neurofeedback Treatment in Learning Disabled Children. Clin. EEG Neurosci. 2003, 34. [Google Scholar] [CrossRef]
- Becerra, J.; Fernández, T.; Harmony, T.; Caballero, M.I.; Garcia, F.; Fernández-Bouzas, A.; Santiago-Rodriguez, E.; Prado-Alcalá, R.A. Follow-Up Study of Learning-Disabled Children Treated with Neurofeedback or Placebo. Clin. EEG Neurosci. 2006, 37. [Google Scholar] [CrossRef]
- Association, W.M. World medical association declaration of helsinki: Ethical principles for medical research involving human subjects. JAMA 2013, 310, 2191–2194. [Google Scholar]
- Wechsler, D.; Flanagan, D.P. TEA Ediciones. Departamento I+D. In WISC-IV: Escala de Inteligencia de Wechsler Para Niños-IV; TEA Ediciones: Madrid, Spain, 2007; ISBN 8471748789. [Google Scholar]
- Bosch-Bayard, J.; Galan, L.; Aubert Vazquez, E.; Virues Alba, T.; Valdes-Sosa, P.A. Resting State Healthy EEG: The First Wave of the Cuban Normative Database. Front. Neurosci. 2020, 14. [Google Scholar] [CrossRef] [PubMed]
- Matute, E.; Inozemtseva, O.; Gonzalez, A.L.; Chamorro, Y. La Evaluación Neuropsicológica Infantil (ENI): Historia y fundamentos teóricos de su validación, Un acercamiento práctico a su uso y valor diagnóstico. Rev. Neuropsicol. Neuropsiquiatría Neurocienc. 2014. [Google Scholar] [CrossRef]
- Silva-Pereyra, J.; Rivera-Gaxiola, M.; Fernández, T.; Díaz-Comas, L.; Harmony, T.; Fernández-Bouzas, A.; Rodríguez, M.; Bernal, J.; Marosi, E. Are poor readers semantically challenged? An event-related brain potential assessment. Int. J. Psychophysiol. 2003. [Google Scholar] [CrossRef]
- Holcomb, P.J.; Ackerman, P.T.; Dykman, R.A. Auditory event-related potentials in attention and reading disabled boys. Int. J. Psychophysiol. 1986. [Google Scholar] [CrossRef]
- Andersson, U.; Lyxell, B. Working memory deficit in children with mathematical difficulties: A general or specific deficit? J. Exp. Child Psychol. 2007, 96. [Google Scholar] [CrossRef] [PubMed]
- Jeffries, S.; Everatt, J. Working memory: Its role in dyslexia and other specific learning difficulties. Dyslexia 2004, 10. [Google Scholar] [CrossRef] [PubMed]
- Bosch-Bayard, J.; Valdés-Sosa, P.; Virues-Alba, T.; Aubert-Vázquez, E.; John, E.R.; Harmony, T.; Riera-Díaz, J.; Trujillo-Barreto, N. 3D Statistical Parametric Mapping of EEG Source Spectra by Means of Variable Resolution Electromagnetic Tomography (VARETA). Clin. EEG Neurosci. 2001, 32, 47–61. [Google Scholar] [CrossRef] [PubMed]
- Bosch-Bayard, J.; Aubert-Vazquez, E.; Brown, S.T.; Rogers, C.; Kiar, G.; Glatard, T.; Scaria, L.; Galan-Garcia, L.; Bringas-Vega, M.L.; Virues-Alba, T.; et al. A Quantitative EEG Toolbox for the MNI Neuroinformatics Ecosystem: Normative SPM of EEG Source Spectra. Front. Neuroinform. 2020, 14, 33. [Google Scholar] [CrossRef] [PubMed]
- Angelakis, E.; Stathopoulou, S.; Frymiare, J.L.; Green, D.L.; Lubar, J.F.; Kounios, J.; Angelakis, E.; Stathopoulou, S.; Frymiare, J.L.; Angelakis, E.; et al. EEG Neurofeedback: A Brief Overview and an Example of Peak Alpha Frequency Training for Cognitive Enhancement in the Elderly. Clin. Neuropsychol. 2007, 4046. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Barros, D.; Savio, G.; Pérez, M. Evaluación de la percepción auditiva con el sistema Medicid 3E. Rev. CENIC Cienc. Biol. 2002, 33, 93–99. [Google Scholar]
- Pascual-Marqui, R.D. Discrete, 3D distributed, linear imaging methods of electric neuronal activity. Part 1: Exact, zero error localization. Clin. Neurophysiol. 2007. [Google Scholar] [CrossRef]
- Pascual-Marqui, R.D.; Faber, P.L.; Kinoshita, T.; Kochi, K.; Milz, P.; Nishida, K.; Yoshimura, M. Comparing EEG/MEG neuroimaging methods based on localization error, false positive activity, and false positive connectivity. bioRxiv 2018, 269753. [Google Scholar] [CrossRef]
- Nunez, P.L.; Srinivasan, R. Electric Fields of the Brain: The Neurophysics of EEG, 2nd ed.; Oxford University Press: New York, NY, USA, 2009; ISBN 9780199865673. [Google Scholar]
- Biscay, R.J.; Bosch-Bayard, J.F.; Pascual-Marqui, R.D. Unmixing EEG Inverse solutions based on brain segmentation. Front. Neurosci. 2018, 12. [Google Scholar] [CrossRef]
- Lohmann, G.; Margulies, D.S.; Horstmann, A.; Pleger, B.; Lepsien, J.; Schloegl, H.; Stumvoll, M.; Villringer, A.; Turner, R. Eigenvector Centrality Mapping for Analyzing Connectivity Patterns in fMRI Data of the Human Brain. PLoS ONE 2010, 5, e10232. [Google Scholar] [CrossRef]
- Worsley, K.; Taylor, J.; Carbonell, F.; Chung, M.; Duerden, E.; Bernhardt, B.; Lyttelton, O.; Boucher, M.; Evans, A. SurfStat: A Matlab toolbox for the statistical analysis of univariate and multivariate surface and volumetric data using linear mixed effects models and random field theory. Neuroimage 2009, 47, S102. [Google Scholar] [CrossRef]
- Suckling, J.; Bullmore, E. Permutation Tests for Factorially Designed Neuroimaging Experiments. Hum. Brain Mapp. 2004, 22, 193–205. [Google Scholar] [CrossRef]
- Galán, L.; Biscay, R.; Rodríguez, J.L.; Pérez-Abalo, M.C.; Rodríguez, R. Testing topographic differences between event related brain potentials by using non-parametric combinations of permutation tests. Electroencephalogr. Clin. Neurophysiol. 1997, 102. [Google Scholar] [CrossRef]
- Enriquez-geppert, S.; Huster, R.J.; Herrmann, C.S. EEG-Neurofeedback as a Tool to Modulate Cognition and Behavior: A Review Tutorial. Front. Hum. Neurosci. 2017, 11, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Kable, J.W.; Caulfield, M.K.; Falcone, M.; McConnell, M.; Bernardo, L.; Parthasarathi, T.; Cooper, N.; Ashare, R.; Audrain-McGovern, J.; Hornik, R.; et al. No effect of commercial cognitive training on brain activity, choice behavior, or cognitive performance. J. Neurosci. 2017, 37, 7390–7402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Melby-Lervåg, M.; Hulme, C. There is no convincing evidence that working memory training is effective: A reply to Au et al. (2014) and Karbach and Verhaeghen (2014). Psychon. Bull. Rev. 2016, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Redick, T.S.; Shipstead, Z.; Harrison, T.L.; Hicks, K.L.; Fried, D.E.; Hambrick, D.Z.; Kane, M.J.; Engle, R.W. No evidence of intelligence improvement after working memory training: A randomized, placebo-controlled study. J. Exp. Psychol. Gen. 2013, 142. [Google Scholar] [CrossRef]
- Ritchie, S.J.; Tucker-Drob, E.M. How Much Does Education Improve Intelligence? A Meta-Analysis. Psychol. Sci. 2018, 095679761877425. [Google Scholar] [CrossRef] [PubMed]
- Schönenberg, M.; Weingärtner, A.L.; Weimer, K.; Scheeff, J. Believing is achieving—On the role of treatment expectation in neurofeedback applications. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2021, 105. [Google Scholar] [CrossRef] [PubMed]
- Leuchter, A.F.; Cook, I.A.; Witte, E.A.; Morgan, M.; Abrams, M. Changes in brain function of depressed subjects during treatment with placebo. Am. J. Psychiatry 2002, 159. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Zhang, H.; Huang, J.; Duan, C.; Kim, J.J.; Ferrari, M.; Hu, C.S. Stronger resting-state neural oscillations associated with wiser advising from the 2nd- but not the 3rd-person perspective. Sci. Rep. 2020, 10. [Google Scholar] [CrossRef]
- Jensen, O.; Tesche, C.D. Frontal theta activity in humans increases with memory load in a working memory task. Neuroscience 2002. [Google Scholar] [CrossRef] [PubMed]
- Sarnthein, J.; Petsche, H.; Rappelsberger, P.; Shaw, G.L.; von Stein, A. Synchronization between prefrontal and posterior association cortex during human working memory. Proc. Natl. Acad. Sci. USA 1998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwang, G.; Jacobs, J.; Geller, A.; Danker, J.; Sekuler, R.; Kahana, M.J. EEG correlates of verbal and nonverbal working memory. Behav. Brain Funct. 2005, 1, 20. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.J.; Kulubya, E.; Goldin, P.; Goodarzi, A.; Girgis, F. Review of the neural oscillations underlying meditation. Front. Neurosci. 2018, 12. [Google Scholar] [CrossRef]
- Schabus, M.; Heib, D.P.J.; Lechinger, J.; Griessenberger, H.; Klimesch, W.; Pawlizki, A.; Kunz, A.B.; Sterman, B.M.; Hoedlmoser, K. Enhancing sleep quality and memory in insomnia using instrumental sensorimotor rhythm conditioning. Biol. Psychol. 2014, 95, 126–134. [Google Scholar] [CrossRef]
- Hinterberger, T.; Schmidt, S.; Kamei, T.; Walach, H. Decreased electrophysiological activity represents the conscious state of emptiness in meditation. Front. Psychol. 2014, 5. [Google Scholar] [CrossRef] [Green Version]
- Schoenberg, P.L.A.; David, A.S. Biofeedback for psychiatric disorders: A systematic review. Appl. Psychophysiol. Biofeedback 2014, 39, 109–135. [Google Scholar] [CrossRef] [PubMed]
- Fernández, T.; Bosch-Bayard, J.; Harmony, T.; Caballero, M.I.; Díaz-Comas, L.; Galán, L.; Ricardo-Garcell, J.; Aubert, E.; Otero-Ojeda, G. Neurofeedback in Learning Disabled Children: Visual versus Auditory Reinforcement. Appl. Psychophysiol. Biofeedback 2016, 41. [Google Scholar] [CrossRef]
- Fernández, T.; Harmony, T.; Fernández-Bouzas, A.; Díaz-Comas, L.; Prado-Alcalá, R.A.; Valdés-Sosa, P.; Otero, G.; Bosch, J.; Galán, L.; Santiago-Rodríguez, E.; et al. Changes in EEG Current Sources Induced by Neurofeedback in Learning Disabled Children. An Exploratory Study. Appl. Psychophysiol. Biofeedback 2007, 32, 169–183. [Google Scholar] [CrossRef]
- Thatcher, R.W.; Krause, P.J.; Hrybyk, M. Cortico-cortical associations and EEG coherence: A two-compartmental model. Electroencephalogr. Clin. Neurophysiol. 1986, 64. [Google Scholar] [CrossRef]
- Lubar, J.F. Neocortical Dynamics: Implications for Understanding the Role of Neurofeedback and Related Techniques for the Enhancement of Attention. Appl. Psychophysiol. Biofeedback 1997, 22, 111–126. [Google Scholar] [CrossRef] [PubMed]
- Lubar, J.F.; Swartwood, M.O.; Swartwood, J.N.; O’Donnell, P.H. Evaluation of the effectiveness of EEG neurofeedback training for ADHD in a clinical setting as measured by changes in T.O.V.A. scores, behavioral ratings, and WISC-R performance. Biofeedback Self. Regul. 1995. [Google Scholar] [CrossRef] [PubMed]
- Sterman, M.B.; Egner, T. Foundation and Practice of Neurofeedback for the Treatment of Epilepsy. Appl. Psychophysiol. Biofeedback 2006, 31, 21–35. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| NFB Group n = 10 | Sham Group n = 8 | Statistical Differences between Groups | |||
|---|---|---|---|---|---|
| Mean | sd | Mean | sd | ||
| Age | 10.4 | 1.0 | 10.1 | 0.8 | U = 26, p = 0.37 |
| WISC-4 test: | |||||
| Full scale IQ | 90.1 | 12.4 | 89 | 8.5 | U = 30, p = 0.6 |
| WM index | 85 | 11.7 | 94.5 | 13.7 | U = 23, p = 0.23 |
| Female/male ratio | 3/7 | 4/4 | p = 0.63, Fisher’s exact test | ||
| z Theta/alpha ratio * | 2.6 | 0.8 | 2.2 | 0.6 | U = 22, p = 0.34 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martínez-Briones, B.J.; Bosch-Bayard, J.; Biscay-Lirio, R.J.; Silva-Pereyra, J.; Albarrán-Cárdenas, L.; Fernández, T. Effects of Neurofeedback on the Working Memory of Children with Learning Disorders—An EEG Power-Spectrum Analysis. Brain Sci. 2021, 11, 957. https://doi.org/10.3390/brainsci11070957
Martínez-Briones BJ, Bosch-Bayard J, Biscay-Lirio RJ, Silva-Pereyra J, Albarrán-Cárdenas L, Fernández T. Effects of Neurofeedback on the Working Memory of Children with Learning Disorders—An EEG Power-Spectrum Analysis. Brain Sciences. 2021; 11(7):957. https://doi.org/10.3390/brainsci11070957
Chicago/Turabian StyleMartínez-Briones, Benito J., Jorge Bosch-Bayard, Rolando J. Biscay-Lirio, Juan Silva-Pereyra, Lucero Albarrán-Cárdenas, and Thalía Fernández. 2021. "Effects of Neurofeedback on the Working Memory of Children with Learning Disorders—An EEG Power-Spectrum Analysis" Brain Sciences 11, no. 7: 957. https://doi.org/10.3390/brainsci11070957