
Episodic Memory Encoding and Retrieval in Face-Name Paired Paradigm: An fNIRS Study

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Methods
2.1. Participants
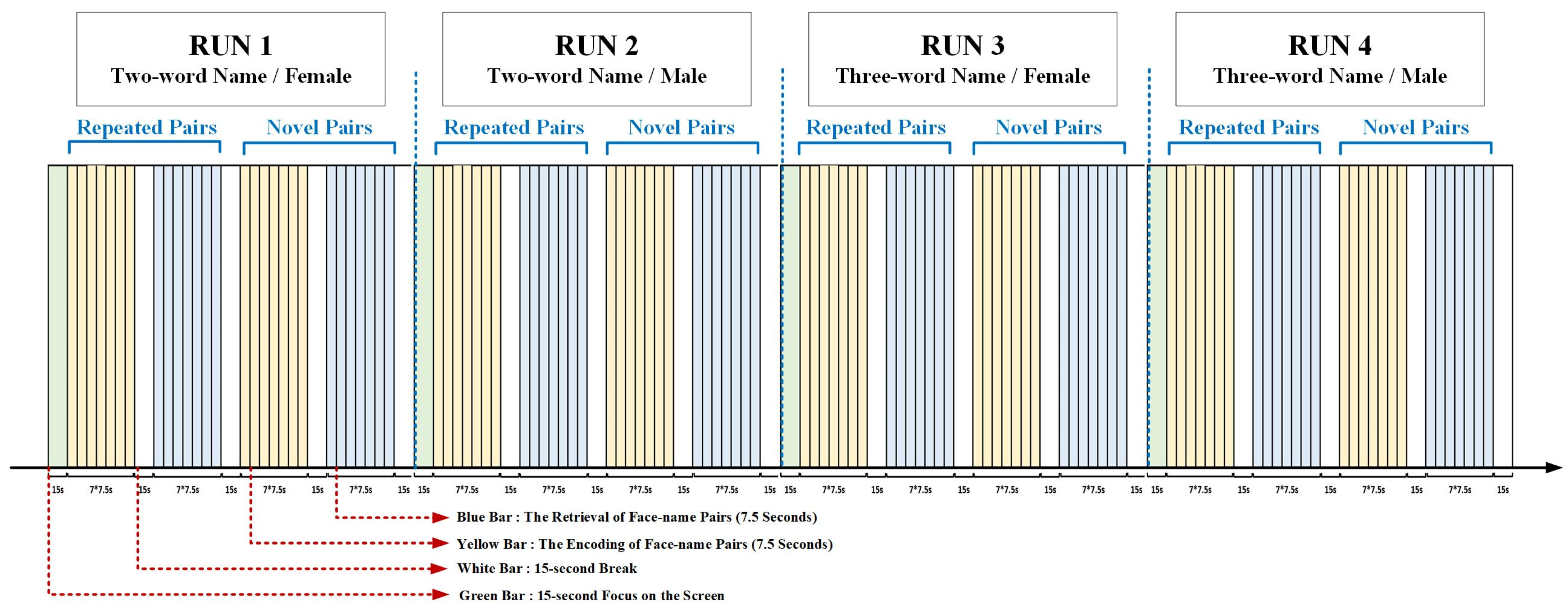
2.2. Experimental Procedure
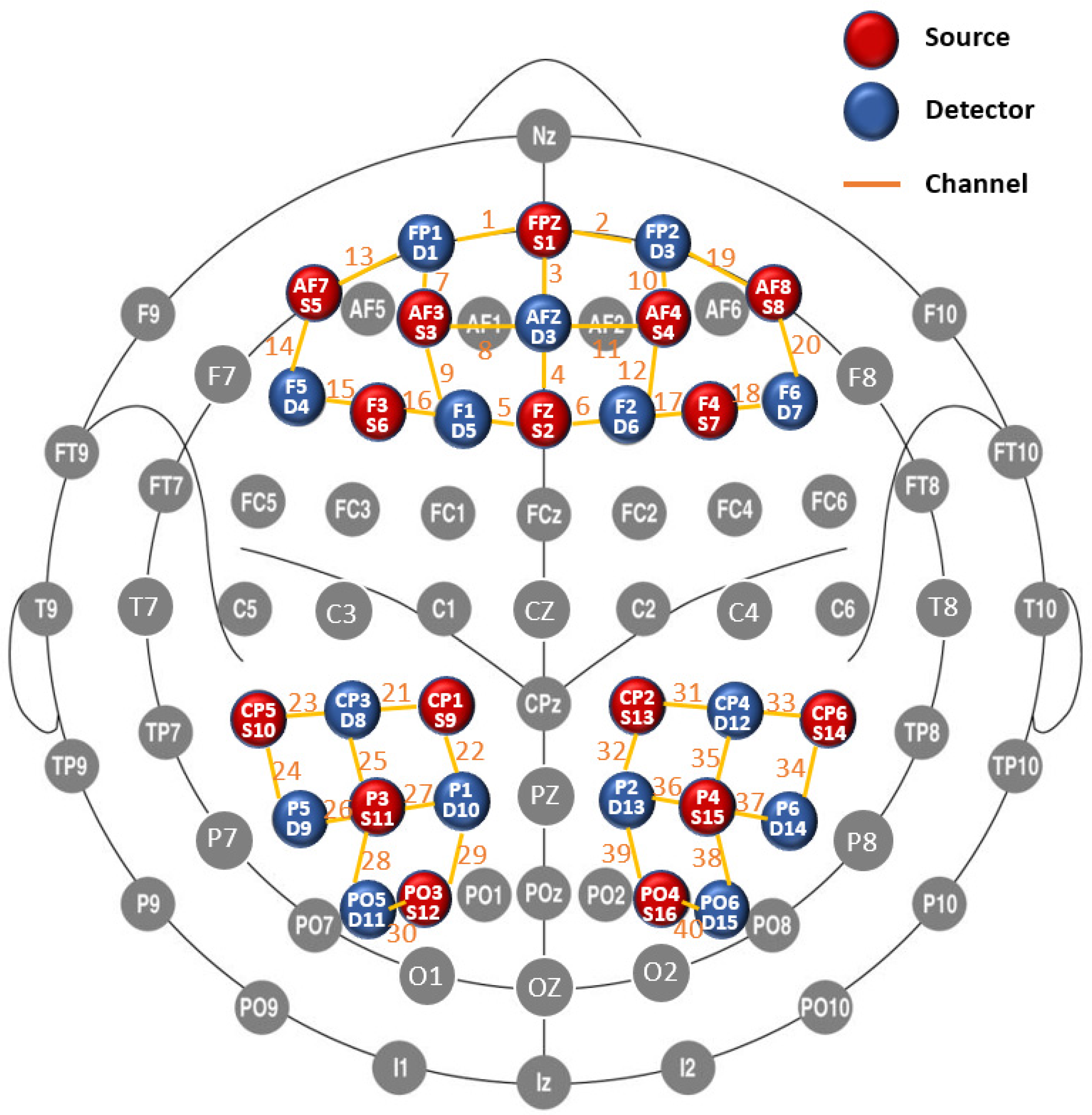
2.2.1. fNIRS Data Acquisition
2.2.2. fNIRS Data Preprocessing
2.3. Data Analysis
3. Results
3.1. Encoding and Retrieval Phases
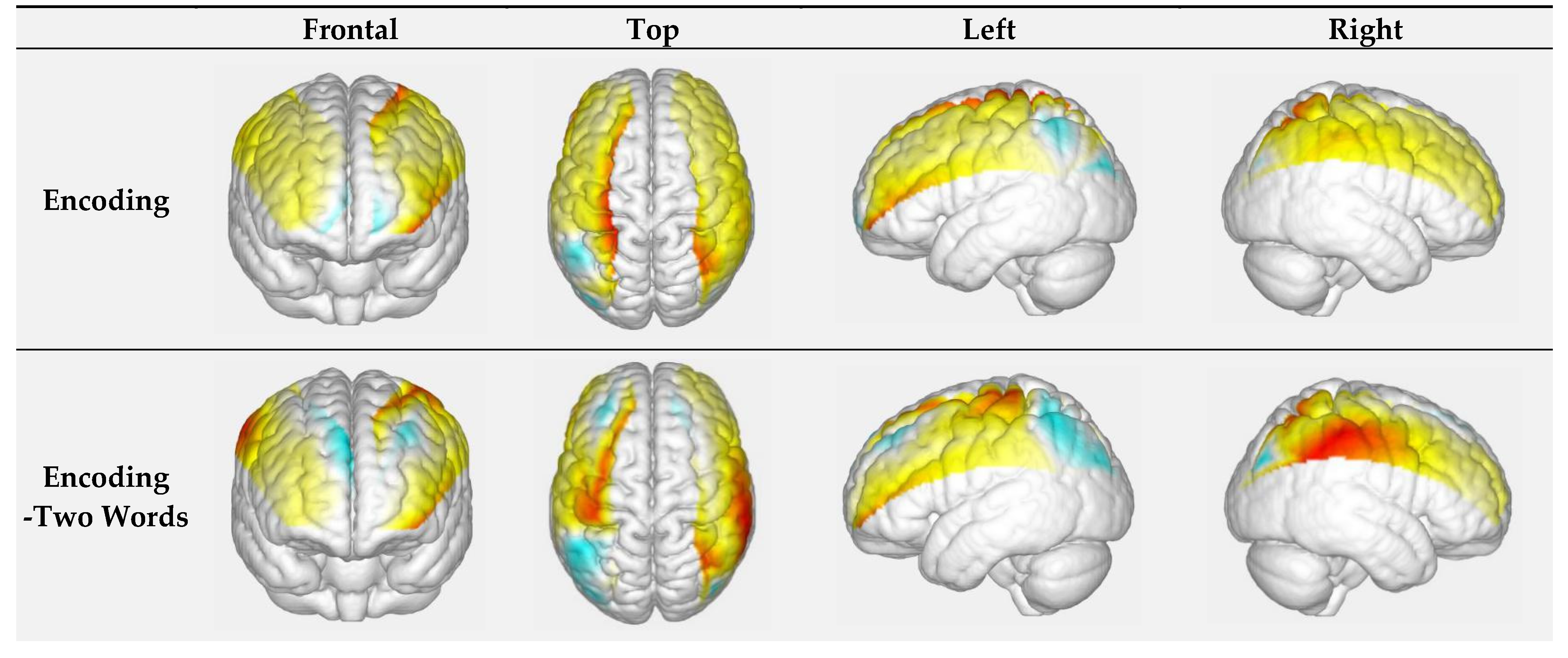
3.1.1. Encoding of Faces with Different Difficulty-Level Names: Two-Word Names vs. Three-Word Names
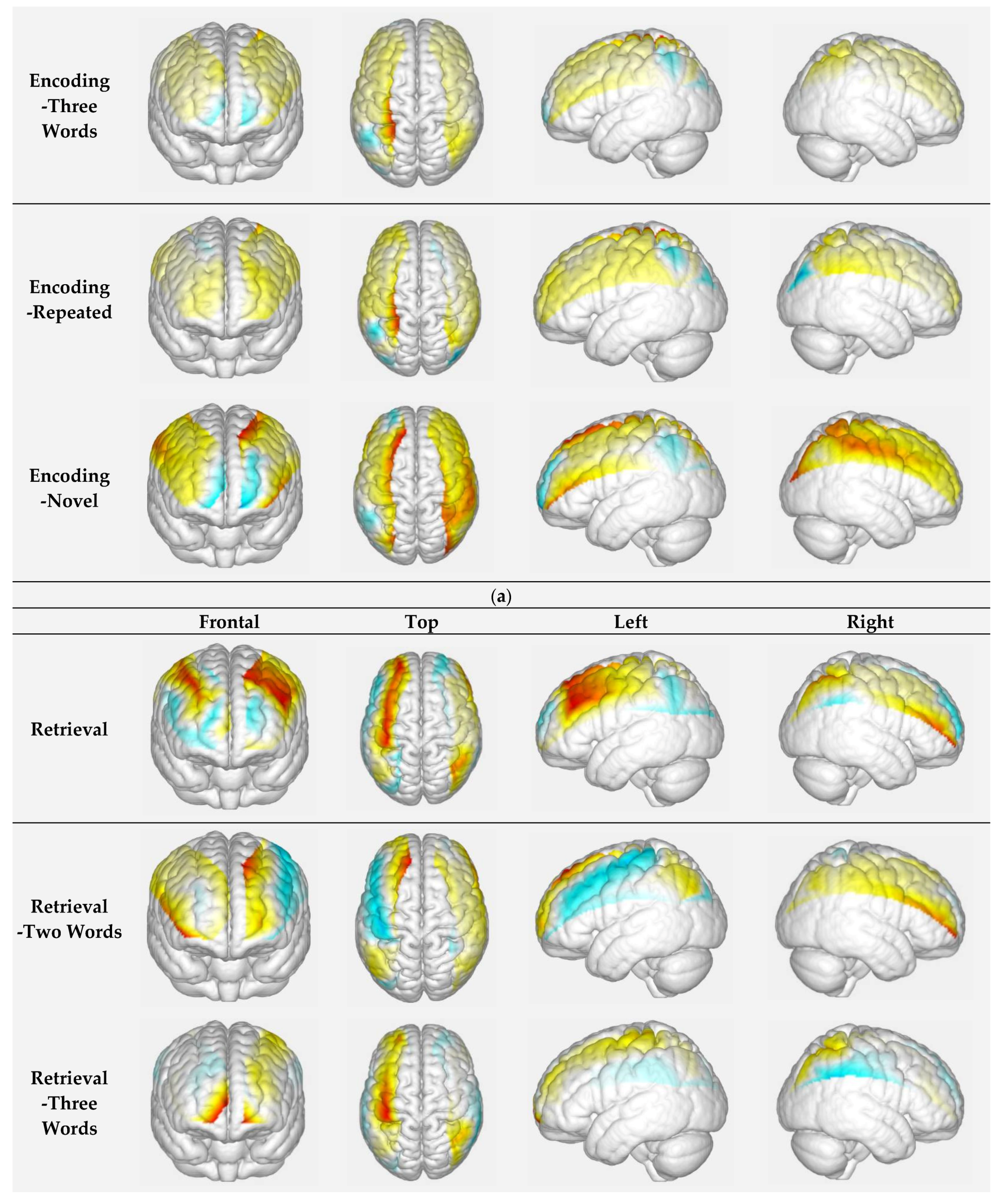
3.1.2. Encoding of Face at Different Novelty Levels: Repeated Faces vs. Novel Faces
3.1.3. Retrieval of Faces with Different Difficulty-Level Names: Two-Word Names vs. Three-Word Names
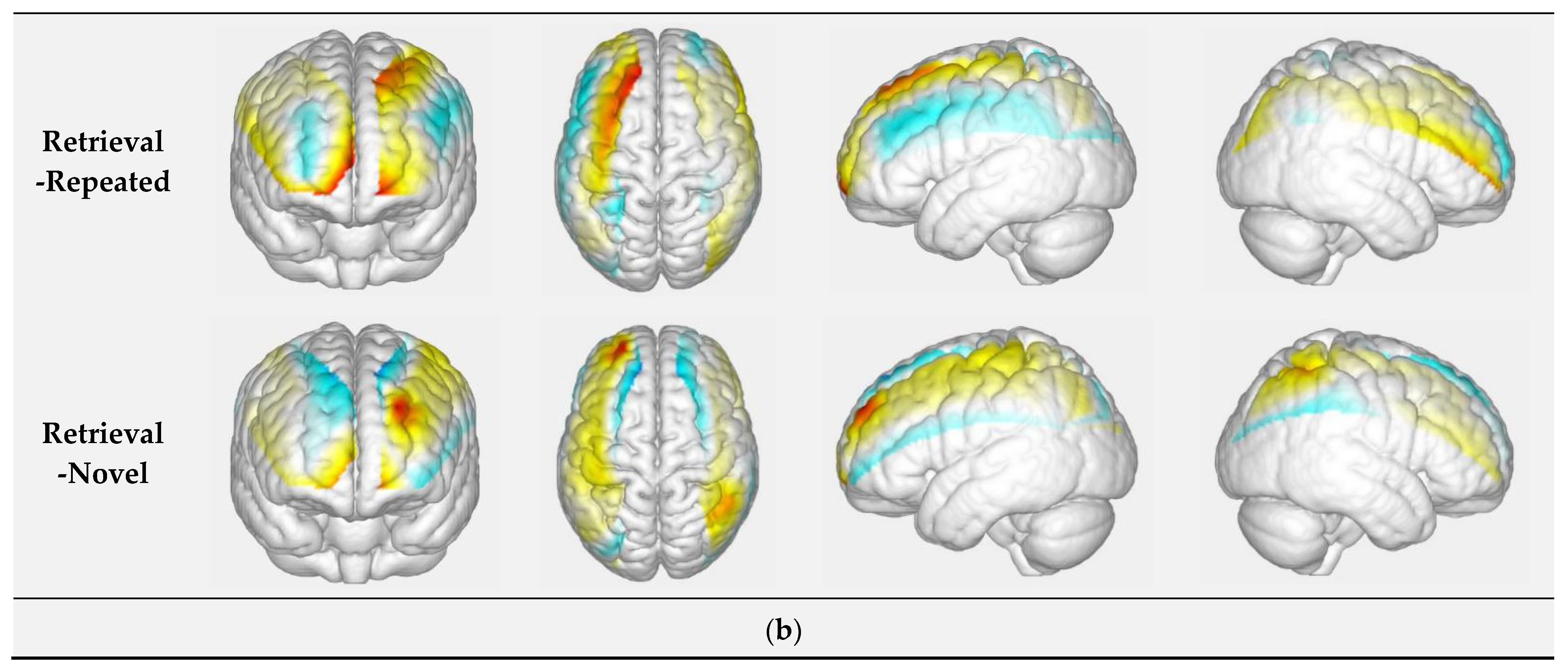
3.1.4. Retrieval of Face at Different Novelty Levels: Repeated Faces vs. Novel Faces
3.2. Correlations between Memory Performance and Brain Activation (Beta Value)
4. Discussion
Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Schacter, D.L.; Gilbert, D.T.; Wegner, D.M. Semantic and Episodic Memory, Psychology. 2009. Worth, New York. Available online: http://www.worthpublishers.com/Catalog/product/psychology-firstedition-s... (accessed on 19 July 2021).
- Tulving, E. Elements of Episodic Memory; Oxford University Press: Oxford, UK, 1983. [Google Scholar]
- Anderson, K.E.; Lynch, K.; Zarahn, E.; Scarmeas, N.; Van Heertum, R.; Sackeim, H.; Moeller, J.R.; Stern, Y. H215O pet study of impairment of nonverbal recognition with normal aging. J. Neuropsychiatry Clin. Neurosci. 2005, 17, 192–200. [Google Scholar] [CrossRef] [PubMed]
- Tromp, D.; Dufour, A.; Lithfous, S.; Pebayle, T.; Després, O. Episodic memory in normal aging and alzheimer disease: Insights from imaging and behavioral studies. Ageing Res. Rev. 2015, 24, 232–262. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.T.; Siddarth, P.; Ercoli, L.M.; Merrill, D.; Torres-Gil, F.; Small, G.W. Modifiable risk factors for alzheimer disease and subjective memory impairment across age groups. PLoS ONE 2014, 9, e98630. [Google Scholar] [CrossRef] [Green Version]
- Conway, M.A.; Gardiner, J.M.; Perfect, T.J.; Anderson, S.J.; Cohen, G.M. Changes in memory awareness during learning: The acquisition of knowledge by psychology undergraduates. J. Exp. Psychol. Gen. 1997, 126, 393–413. [Google Scholar] [CrossRef] [PubMed]
- Herbert, D.B.M.; Burt, J.S. What do students remember? Episodic memory and the development of schematization. Appl. Cogn. Psychol. 2004, 18, 77–88. [Google Scholar] [CrossRef]
- Irish, M.; Lawlor, B.A.; Coen, R.F.; O’Mara, S.M. Everyday episodic memory in amnestic mild cognitive impairment: A preliminary investigation. BMC Neurosci. 2011, 12, 80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paradise, M.; McCade, D.; Hickie, I.B.; Diamond, K.; Lewis, S.; Naismith, S. Caregiver burden in mild cognitive impairment. Aging Ment. Health 2015, 19, 72–78. [Google Scholar] [CrossRef]
- Wong, W. Economic burden of alzheimer disease and managed care considerations. Am. J. Manag. Care. 2020, 26, S177–S183. [Google Scholar] [CrossRef]
- Blackwell, A.D.; Sahakian, B.; Vesey, R.; Semple, J.M.; Robbins, T.W.; Hodges, J.R. Detecting dementia: Novel neuropsychological markers of preclinical alzheimer’s disease. Dement. Geriatr. Cogn. Disord. 2004, 17, 42–48. [Google Scholar] [CrossRef]
- De Jager, C.; Blackwell, A.D.; Budge, M.M.; Sahakian, B.J. Predicting cognitive decline in healthy older adults. Am. J. Geriatr. Psychiatry 2005, 13, 735–740. [Google Scholar] [CrossRef]
- Fowler, K.S.; Saling, M.M.; Conway, E.L.; Semple, J.M.; Louisp, W.J. Paired associate performance in the early detection of dat. J. Int. Neuropsychol. Soc. 2002, 8, 58–71. [Google Scholar] [CrossRef]
- Crystal, J.D. Elements of episodic-like memory in animal models. Behav. Process. 2009, 80, 269–277. [Google Scholar] [CrossRef]
- Eichenbaum, H.; Alvarez, P.; Ramus, S.J. Animal Models of Amnesia; Elsevier: Amsterdam, The Netherlands, 2000. [Google Scholar]
- Glahn, D.C.; Robinson, J.L.; Tordesillas-Gutierrez, D.; Monkul, E.S.; Holmes, M.K.; Green, M.; Bearden, C. Fronto-temporal dysregulation in asymptomatic bipolar i patients: A paired associate functional mri study. Hum. Brain Mapp. 2010, 31, 1041–1051. [Google Scholar] [CrossRef] [Green Version]
- Köhler, S.; Paus, T.; Buckner, R.L.; Milner, B. Effects of left inferior prefrontal stimulation on episodic memory formation: A two-stage fmri-rtms study. J. Cogn. Neurosci. 2004, 16, 178–188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sidhu, M.K.; Stretton, J.; Winston, G.; Bonelli, S.; Centeno, M.; Vollmar, C.; Symms, M.; Thompson, P.J.; Koepp, M.J.; Duncan, J.S. A functional magnetic resonance imaging study mapping the episodic memory encoding network in temporal lobe epilepsy. Brain 2013, 136, 1868–1888. [Google Scholar] [CrossRef] [PubMed]
- Rugg, M.D.; Johnson, J.D.; Uncapher, M.R. Encoding and Retrieval in Episodic Memory: Insights from Fmri; Wiley Blackwell: Hoboken, NJ, USA, 2015. [Google Scholar]
- Adorni, R.; Gatti, A.; Brugnera, A.; Sakatani, K.; Compare, A. Could fNIRS Promote Neuroscience Approach in Clinical Psychology? Front. Psychol. 2016, 7, 456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yücel, M.A.; Lühmann, A.V.; Scholkmann, F.; Gervain, J.; Dan, I.; Ayaz, H.; Boas, D.; Cooper, R.J.; Culver, J.; Elwell, C.E.; et al. Best practices for fnirs publications. Neurophotonics 2021, 8, 012101. [Google Scholar] [CrossRef]
- Jahani, S.; Fantana, A.L.; Harper, D.; Ellison, J.M.; Boas, D.A.; Forester, B.P.; Yücel, M.A. Fnirs can robustly measure brain activity during memory encoding and retrieval in healthy subjects. Sci. Rep. 2017, 7, 9533. [Google Scholar] [CrossRef] [PubMed]
- Cabeza, R.; Nyberg, L. Imaging cognition ii: An empirical review of 275 pet and fmri studies. J. Cogn. Neurosci. 2000, 12, 1–47. [Google Scholar] [CrossRef] [PubMed]
- Gui, P.; Ku, Y.; Li, L.; Li, X.; Bodner, M.; Lenz, F.A.; Wang, L.; Zhou, Y.-D. Neural correlates of visuo-tactile crossmodal paired-associate learning and memory in humans. Neuroscience 2017, 362, 181–195. [Google Scholar] [CrossRef]
- Preston, A.R.; Eichenbaum, H. Interplay of hippocampus and prefrontal cortex in memory. Curr. Biol. 2013, 23, R764–R773. [Google Scholar] [CrossRef] [Green Version]
- Hunkin, N.M.; Mayes, A.R.; Gregory, L.J.; Nicholas, A.K.; Nunn, J.A.; Brammer, M.J.; Bullmore, E.T.; Williams, S.C. Novelty-related activation within the medial temporal lobes. Neuropsychologia 2002, 40, 1456–1464. [Google Scholar] [CrossRef]
- Nyberg, L.; Tulving, E.; Habib, R.; Nilsson, L.G.; Kapur, S.; Houle, S.; Cabeza, R.; McIntosh, A.R. Functional brain maps of retrieval mode and recovery of episodic information. Neuroreport 1995, 7, 249–252. [Google Scholar] [CrossRef]
- Schacter, D.L.; Reiman, E.; Uecker, A.; Roister, M.R.; Yun, L.S.; Cooper, L.A. Brain regions associated with retrieval of structurally coherent visual information. Nature 1995, 376, 587–590. [Google Scholar] [CrossRef]
- Tulving, E.; Markowitsch, H.J.; Craik, F.I.M.; Habib, R.; Houle, S. Novelty and familiarity activations in pet studies of memory encoding and retrieval. Cereb. Cortex. 1996, 6, 71–79. [Google Scholar] [CrossRef] [Green Version]
- Sperling, R.A.; Bates, J.F.; Cocchiarella, A.J.; Schacter, D.L.; Rosen, B.R.; Albert, M.S. Encoding novel face-name associations: A functional mri study. Hum. Brain Mapp. 2001, 14, 129–139. [Google Scholar] [CrossRef]
- Zimeo Morais, G.A.; Balardin, J.B.; Sato, J.R. fNIRS Optodes’ Location Decider (fOLD): A toolbox for probe arrangement guided by brain regions-of-interest. Sci. Rep. 2018, 8, 3341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinti, P.; Scholkmann, F.; Hamilton, A.; Burgess, P.; Tachtsidis, I. Current status and issues regarding pre-processing of fnirs neuroimaging data: An investigation of diverse signal filtering methods within a general linear model framework. Front. Hum. Neurosci. 2018, 12, 505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, F.; Kozel, F.A.; Yennu, A.S.; Croarkin, P.E.; McClintock, S.M.; Mapes, K.S.; Husain, M.M.; Liu, H. Test-retest assessment of cortical activation induced by repetitive transcranial magnetic stimulation with brain atlas-guided optical topography. J. Biomed. Opt. 2012, 17, 116020. [Google Scholar] [CrossRef] [PubMed]
- Tian, F.; Lin, Z.J.; Liu, H. Easytopo: A toolbox for rapid diffuse optical topography based on a standard template of brain atlas. In Proceedings of the SPIE. 8578: 85782J, San Francisco, CA, USA, 25 March 2013. [Google Scholar] [CrossRef]
- Strangman, G.; Culver, J.P.; Thompson, J.H.; Boas, D.A. A quantitative comparison of simultaneous bold fmri and nirs recordings during functional brain activation. Neuroimage 2002, 17, 719–731. [Google Scholar] [CrossRef] [PubMed]
- Ghasemi, A.; Zahediasl, S. Normality tests for statistical analysis: A guide for non-statisticians. Int. J. Endocrinol. Metab. 2012, 10, 486–489. [Google Scholar] [CrossRef] [Green Version]
- Shapiro, S.S.; Wilk, M.B.; Chen, H.J. A comparative study of various tests for normality. J. Am. Stat. Assoc. 1968, 63, 1343–1372. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. Ser. B (Methodol.) 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Singh, A.K.; Dan, I. Exploring the false discovery rate in multichannel nirs. Neuroimage 2006, 33, 542–549. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W. P < 0.05, < 0.01, < 0.001, < 0.0001, < 0.00001, < 0.000001, or < 0.0000001. J. Sport Health Sci. 2016, 5, 77–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blumenfeld, R.S.; Ranganath, C. Prefrontal cortex and long-term memory encoding: An integrative review of findings from neuropsychology and neuroimaging. Neuroscientist 2007, 13, 280–291. [Google Scholar] [CrossRef] [PubMed]
- Demb, J.B.; Desmond, J.E.; Wagner, A.D.; Vaidya, C.J.; Glover, G.H.; Gabrieli, J.D. Semantic encoding and retrieval in the left inferior prefrontal cortex: A functional mri study of task difficulty and process specificity. J. Neurosci. 1995, 15, 5870–5878. [Google Scholar] [CrossRef]
- Opitz, B.; Mecklinger, A.; Friederici, A.D. Functional asymmetry of human prefrontal cortex: Encoding and retrieval of verbally and nonverbally coded information. Learn. Mem. 2000, 7, 85–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lundstrom, B.N.; Petersson, K.M.; Andersson, J.; Johansson, M.; Fransson, P.; Ingvar, M. Isolating the retrieval of imagined pictures during episodic memory: Activation of the left precuneus and left prefrontal cortex. Neuroimage 2003, 20, 1934–1943. [Google Scholar] [CrossRef] [PubMed]
- Friederici, A.D. Towards a neural basis of auditory sentence processing. Trends Cogn. Sci. 2002, 6, 78–84. [Google Scholar] [CrossRef] [Green Version]
- Hagoort, P.; Hald, P.; Petersson, K.M. Semantic vs. world knowledge integration during sentence comprehension. J. Cogn. Neurosci. Suppl. 2002, 159. [Google Scholar]
- Wagner, A.D.; Paré-Blagoev, E.J.; Clark, J.; Poldrack, R.A. Recovering meaning: Left prefrontal cortex guides controlled semantic retrieval. Neuron 2001, 31, 329–338. [Google Scholar] [CrossRef] [Green Version]
- Sestieri, C.; Shulman, G.L.; Corbetta, M. The contribution of the human posterior parietal cortex to episodic memory. Nat. Rev. Neurosci. 2017, 18, 183–192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buckner, R.L.; Andrews-Hanna, J.R.; Schacter, D.L. The brain’s default network: Anatomy, function, and relevance to disease. Ann. N. Y. Acad. Sci. 2008, 1124, 1–38. [Google Scholar] [CrossRef] [Green Version]
- White, T.P.; Jansen, M.; Doege, K.; Mullinger, K.; Park, S.B.; Liddle, E.B.; Gowland, P.; Francis, S.T.; Bowtell, R.; Liddle, P.F. Theta power during encoding predicts subsequent-memory performance and default mode network deactivation. Hum. Brain Mapp. 2013, 34, 2929–2943. [Google Scholar] [CrossRef]
- Sestieri, C.; Corbetta, M.; Romani, G.L.; Shulman, G.L. Episodic memory retrieval, parietal cortex, and the default mode network: Functional and topographic analyses. J. Neurosci. 2011, 31, 4407–4420. [Google Scholar] [CrossRef] [PubMed]
- Wagner, A.D.; Shannon, B.J.; Kahn, I.; Buckner, R.L. Parietal lobe contributions to episodic memory retrieval. Trends Cogn. Sci. 2005, 9, 445–453. [Google Scholar] [CrossRef]
- Nolde, S.F.; Johnson, M.K.; Raye, C.L. The role of prefrontal cortex during tests of episodic memory. Trends Cogn. Sci. 1998, 2, 399–406. [Google Scholar] [CrossRef]
- Fletcher, P.C.; Henson, R.N. Frontal lobes and human memory: Insights from functional neuroimaging. Brain 2001, 124, 849–881. [Google Scholar] [CrossRef] [Green Version]
- Pinti, P.; Merla, A.; Aichelburg, C.; Lind, F.; Power, S.; Swingler, E.; Hamilton, A.; Gilbert, S.; Burgess, P.W.; Tachtsidis, I. A novel glm-based method for the automatic identification of functional events (aide) in fnirs data recorded in naturalistic environments. Neuroimage 2017, 155, 291–304. [Google Scholar] [CrossRef] [PubMed]
- Thompson-Schill, S.L.; D’Esposito, M.; Kan, I.P. Effects of repetition and competition on activity in left prefrontal cortex during word generation. Neuron 1999, 23, 513–522. [Google Scholar] [CrossRef] [Green Version]
- Huff, T.; Mahabadi, N.; Tadi, P. Neuroanatomy, Visual Cortex; StatPearls: Treasure Island, FL, USA, 2020. [Google Scholar]
- Qiao, F.; Zheng, L.; Li, L.; Zhu, L.; Wang, Q. Reduced repetition suppression in the occipital visual cortex during repeated negative chinese personality-trait word processing. Scand. J. Psychol. 2014, 55, 533–537. [Google Scholar] [CrossRef]
- Grill-Spector, K.; Henson, R.; Martin, A. Repetition and the brain: Neural models of stimulus-specific effects. Trends Cogn. Sci. 2006, 10, 14–23. [Google Scholar] [CrossRef]
- Utzerath, C.; John-Saaltink, E.S.; Buitelaar, J.; De Lange, F.P. Repetition suppression to objects is modulated by stimulus-specific expectations. Sci. Rep. 2017, 7, 8781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franks, J.J.; Bilbrey, C.W.; Lien, K.G.; McNamara, T.P. Transfer-appropriate processing (tap) and repetition priming. Mem. Cognit. 2000, 28, 1140–1151. [Google Scholar] [CrossRef] [Green Version]
- Morris, C.D.; Bransford, J.D.; Franks, J.J. Levels of processing versus transfer appropriate processing. J. Verbal Learn. Verbal Behav. 1977, 16, 519–533. [Google Scholar] [CrossRef]
- Tulving, E.; Thomson, D.M. Encoding specificity and retrieval processes in episodic memory. Psychol. Rev. 1973, 80, 353–373. [Google Scholar] [CrossRef] [Green Version]
- Morcom, A.M. Re-engaging with the past: Recapitulation of encoding operations during episodic retrieval. Front. Hum. Neurosci. 2014, 8, 351. [Google Scholar] [CrossRef] [Green Version]
- Herold, F.; Wiegel, P.; Scholkmann, F.; Müller, N.G. Applications of functional near-infrared spectroscopy (fnirs) neuroimaging in exercise-cognition science: A systematic, methodology- focused review. J. Clin. Med. 2018, 7, 466. [Google Scholar] [CrossRef] [Green Version]
- Tachtsidis, I.; Scholkmann, F. False positives and false negatives in functional near-infrared spectroscopy: Issues, challenges, and the way forward. Neurophotonics 2016, 3, 031405. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Conditions | Channel | Brodmann Area | Location | Baseline (Rest) | Task | FDR Corrected p-Value | ||
|---|---|---|---|---|---|---|---|---|
| Mean | SD | Mean | SD | |||||
| Encoding | C1 | 11 | Left Prefrontal Cortex | −0.001 | 0.011 | −0.052 | 0.337 | 0.048 |
| C3 | / | Prefrontal Cortex | −0.002 | 0.012 | −0.050 | 0.314 | 0.048 | |
| C17 | 9 | Right Prefrontal Cortex | −0.001 | 0.016 | −0.102 | 0.451 | 0.012 | |
| C18 | 45 | 0.000 | 0.009 | −0.069 | 0.455 | 0.048 | ||
| C20 | 46 | 0.000 | 0.008 | −0.049 | 0.258 | 0.029 | ||
| C22 | / | Left Superior Parietal Cortex | 0.003 | 0.028 | 0.345 | 2.070 | 0.038 | |
| C25 | 40 | Left Inferior Parietal Cortex | −0.002 | 0.014 | −0.219 | 0.815 | 0.000 | |
| C26 | 39 | −0.002 | 0.013 | −0.198 | 0.868 | 0.012 | ||
| Encoding -Two Words | C15 | 45 | Left Prefrontal Cortex | 0.000 | 0.008 | −0.135 | 0.468 | 0.026 |
| C17 | 9 | Right Prefrontal Cortex | 0.000 | 0.009 | −0.104 | 0.425 | 0.034 | |
| C25 | 40 | Left Inferior Parietal Cortex | 0.000 | 0.012 | −0.268 | 0.748 | 0.010 | |
| C26 | 39 | −0.004 | 0.012 | −0.305 | 0.895 | 0.012 | ||
| C30 | 18 | Left Middle Occipital Cortex | −0.002 | 0.012 | −0.184 | 0.842 | 0.048 | |
| Encoding -Three Words | C19 | 11 | Right Prefrontal Cortex | 0.000 | 0.005 | −0.012 | 0.298 | 0.035 |
| C20 | 46 | 0.000 | 0.004 | −0.042 | 0.283 | 0.035 | ||
| C23 | 40 | Left Inferior Parietal Cortex | 0.001 | 0.018 | −0.045 | 1.745 | 0.010 | |
| C33 | 40 | Right Inferior Parietal Cortex | 0.002 | 0.022 | 0.222 | 2.251 | 0.035 | |
| C37 | 39 | 0.000 | 0.016 | −0.023 | 1.460 | 0.048 | ||
| Encoding -Repeated | C18 | 45 | Right Prefrontal Cortex | 0.000 | 0.008 | −0.106 | 0.383 | 0.026 |
| C22 | / | Left Superior Parietal Cortex | 0.006 | 0.028 | 0.558 | 2.224 | 0.035 | |
| C25 | 40 | Left Inferior Parietal Cortex | 0.000 | 0.013 | −0.252 | 0.730 | 0.010 | |
| Encoding -Novel | C2 | 11 | Right Prefrontal Cortex | −0.002 | 0.011 | −0.085 | 0.383 | 0.048 |
| C17 | 9 | −0.003 | 0.020 | −0.135 | 0.394 | 0.012 | ||
| C23 | 40 | Left Inferior Parietal Cortex | −0.004 | 0.018 | −0.199 | 0.692 | 0.026 | |
| C26 | 39 | −0.004 | 0.015 | −0.233 | 0.930 | 0.035 | ||
| Retrieval | C1 | 11 | Left Prefrontal Cortex | 0.002 | 0.018 | 0.237 | 1.551 | 0.048 |
| Retrieval -Two words | / | / | / | / | / | / | / | |
| Retrieval -Three words | C7 | 11 | Left Prefrontal Cortex | 0.001 | 0.012 | 0.121 | 0.456 | 0.031 |
| C19 | 11 | Right Prefrontal Cortex | 0.000 | 0.006 | 0.091 | 0.374 | 0.035 | |
| C20 | 46 | −0.001 | 0.005 | 0.084 | 0.320 | 0.031 | ||
| Retrieval -Repeated | / | / | / | / | / | / | / | |
| Retrieval -Novel | / | / | / | / | / | / | / | |
| (1) Encoding vs. Retrieval | ||||||
| Channel | Location | Encoding | Retrieval | FDR Corrected p-Value | ||
| Mean | SD | Mean | SD | |||
| C1 | Left Prefrontal Cortex | −0.179 | 0.926 | 0.881 | 1.936 | 0.021 |
| C10 | Right Prefrontal Cortex | 0.217 | 0.291 | −0.206 | 0.723 | 0.022 |
| C12 | 0.181 | 0.283 | −0.199 | 0.549 | 0.022 | |
| C40 | Right Middle Occipital Cortex | 0.066 | 0.843 | 0.109 | 0.818 | 0.021 |
| (2) Encoding (Two Words) vs. Encoding (Three Words) | ||||||
| Channel | Location | Encoding (Two Words) | Encoding (Three Words) | FDR Corrected p-Value | ||
| Mean | SD | Mean | SD | |||
| C4 | Prefrontal Cortex | −0.390 | 1.174 | 0.456 | 1.002 | 0.032 |
| (3) Encoding (Repeated) vs. Encoding (Novel) | ||||||
| Channel | Location | Encoding (Repeated) | Encoding (Novel) | FDR Corrected p-Value | ||
| Mean | SD | Mean | SD | |||
| C11 | Right Prefrontal Cortex | 0.223 | 0.476 | −0.002 | 0.274 | 0.022 |
| C38 | Right Middle Occipital Cortex | −0.713 | 1.530 | 0.342 | 1.199 | 0.034 |
| (4) Retrieval (Two Words) vs. Retrieval (Three Words) | ||||||
| Channel | Location | Retrieval (Two Words) | Retrieval (Three Words) | FDR Corrected p-Value | ||
| Mean | SD | Mean | SD | |||
| / | / | / | / | / | / | / |
| (5) Retrieval (Repeated) vs. Retrieval (Novel) | ||||||
| Channel | Location | Retrieval (Repeated) | Retrieval (Novel) | FDR Corrected p-Value | ||
| Mean | SD | Mean | SD | |||
| C5 | Left Prefrontal Cortex | 0.956 | 2.210 | −0.618 | 2.242 | 0.021 |
| C30 | Left Middle Occipital Cortex | −0.273 | 1.005 | 0.289 | 0.826 | 0.036 |
| C31 | Right Inferior Parietal Cortex | −0.052 | 0.344 | 0.178 | 0.310 | 0.046 |
| C32 | Right Superior Parietal Cortex | −0.150 | 1.183 | 0.120 | 1.015 | 0.022 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, Q.; Cheval, B.; Becker, B.; Herold, F.; Chan, C.C.H.; Delevoye-Turrell, Y.N.; Guérin, S.M.R.; Loprinzi, P.; Mueller, N.; Zou, L. Episodic Memory Encoding and Retrieval in Face-Name Paired Paradigm: An fNIRS Study. Brain Sci. 2021, 11, 951. https://doi.org/10.3390/brainsci11070951
Yu Q, Cheval B, Becker B, Herold F, Chan CCH, Delevoye-Turrell YN, Guérin SMR, Loprinzi P, Mueller N, Zou L. Episodic Memory Encoding and Retrieval in Face-Name Paired Paradigm: An fNIRS Study. Brain Sciences. 2021; 11(7):951. https://doi.org/10.3390/brainsci11070951
Chicago/Turabian StyleYu, Qian, Boris Cheval, Benjamin Becker, Fabian Herold, Chetwyn C. H. Chan, Yvonne N. Delevoye-Turrell, Ségolène M. R. Guérin, Paul Loprinzi, Notger Mueller, and Liye Zou. 2021. "Episodic Memory Encoding and Retrieval in Face-Name Paired Paradigm: An fNIRS Study" Brain Sciences 11, no. 7: 951. https://doi.org/10.3390/brainsci11070951