
Early Spatial Memory Impairment in a Double Transgenic Model of Alzheimer’s Disease TgF-344 AD

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects
2.2. Behavioural Testing
2.2.1. Vision Test
2.2.2. Active Allothetic Place Avoidance (AAPA)
2.2.3. Elevated Plus Maze (EPM)
2.3. Statistics
3. Results
3.1. Visible Platform Water Maze Test
3.2. Elevated Plus Maze
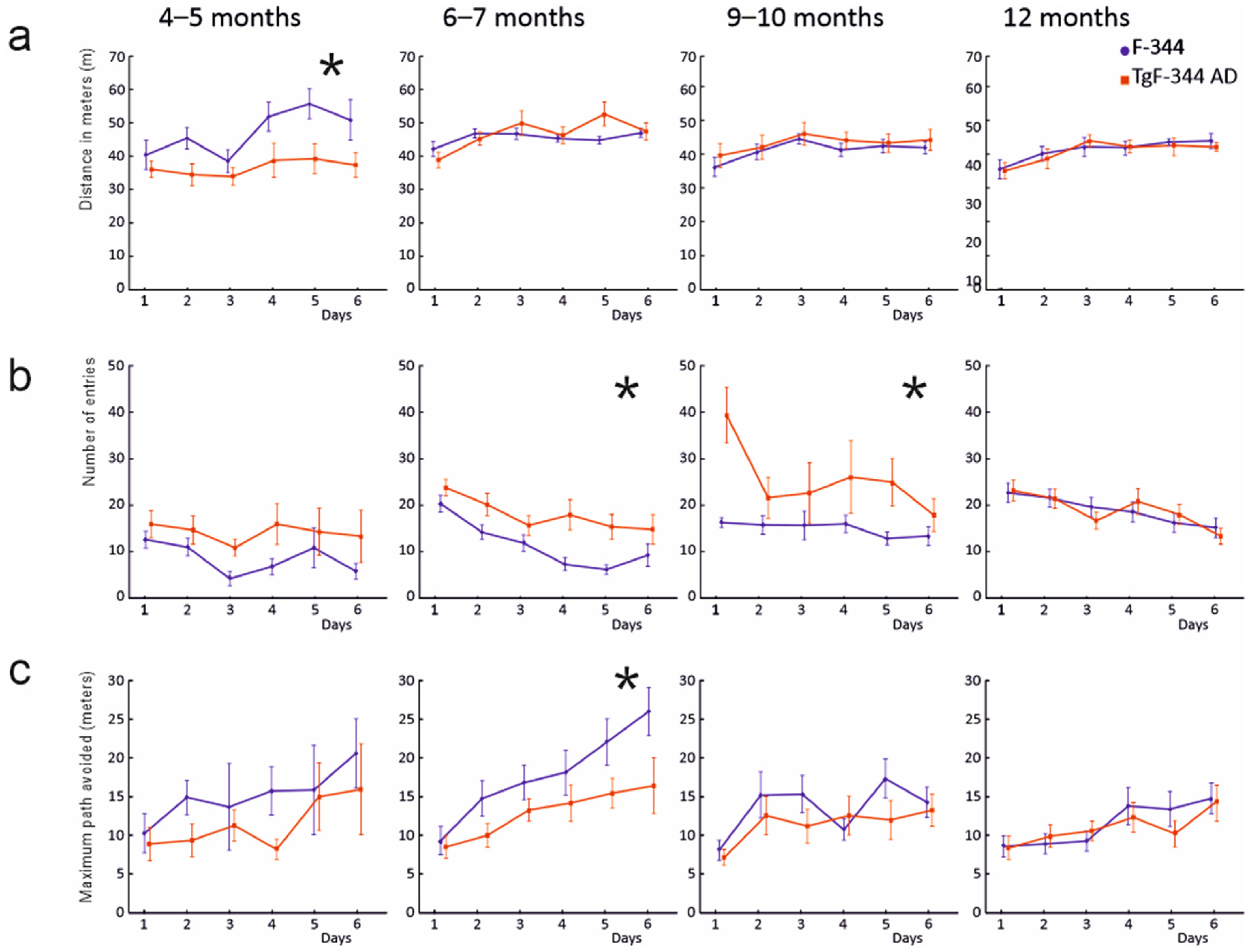
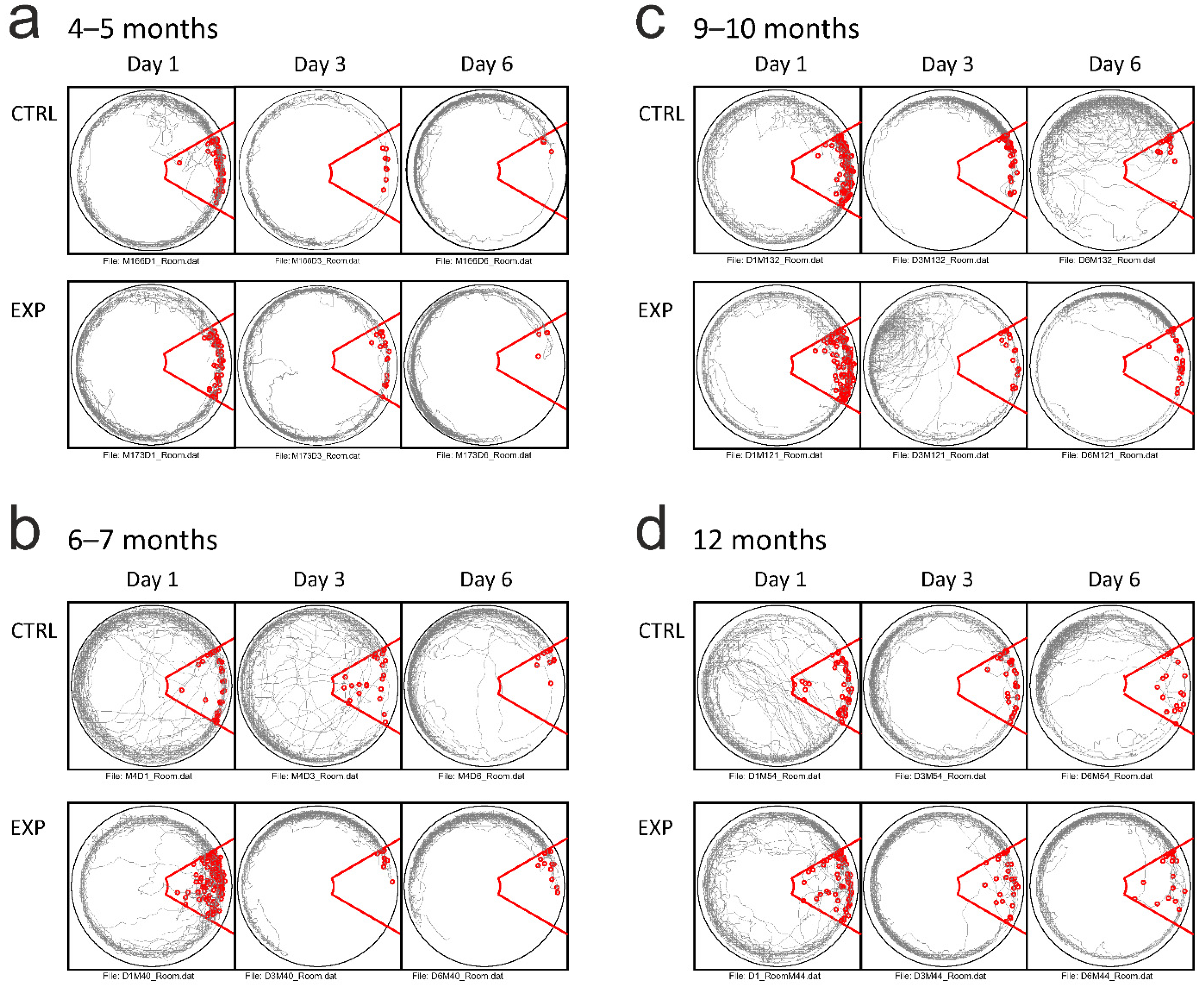
3.3. Active Allothetic Place Avoidance Test
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Querfurth, H.W.; LaFerla, F.M. Alzheimer’s Disease. N. Engl. J. Med. 2010, 362, 329–344. [Google Scholar] [CrossRef] [Green Version]
- Selkoe, D.J. Alzheimer’s Disease: Genes, Proteins, and Therapy. Physiol. Rev. 2001, 81, 741–766. [Google Scholar] [CrossRef]
- Braak, H.; Braak, E. Neuropathological Stageing of Alzheimer-Related Changes. Acta Neuropathol. 1991, 82, 239–259. [Google Scholar] [CrossRef] [PubMed]
- Chambers, J.K.; Tokuda, T.; Uchida, K.; Ishii, R.; Tatebe, H.; Takahashi, E.; Tomiyama, T.; Une, Y.; Nakayama, H. The Domestic Cat as a Natural Animal Model of Alzheimer’s Disease. Acta Neuropathol. Commun. 2015, 3, 78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeTure, M.A.; Dickson, D.W. The Neuropathological Diagnosis of Alzheimer’s Disease. Mol. Neurodegener. 2019, 14, 32. [Google Scholar] [CrossRef] [Green Version]
- Myers, A.; McGonigle, P. Overview of Transgenic Mouse Models for Alzheimer’s Disease. Curr. Protoc. Neurosci. 2019, 89, e81. [Google Scholar] [CrossRef] [PubMed]
- Price, D.L.; Sisodia, S.S. Mutant Genes in Familial Alzheimer’s Disease and Transgenic Models. Annu. Rev. Neurosci. 1998, 21, 479–505. [Google Scholar] [CrossRef]
- Flood, D.G.; Lin, Y.-G.; Lang, D.M.; Trusko, S.P.; Hirsch, J.D.; Savage, M.J.; Scott, R.W.; Howland, D.S. A Transgenic Rat Model of Alzheimer’s Disease with Extracellular Abeta Deposition. Neurobiol. Aging 2009, 30, 1078–1090. [Google Scholar] [CrossRef] [PubMed]
- Leon, W.C.; Canneva, F.; Partridge, V.; Allard, S.; Ferretti, M.T.; DeWilde, A.; Vercauteren, F.; Atifeh, R.; Ducatenzeiler, A.; Klein, W.; et al. A Novel Transgenic Rat Model with a Full Alzheimer’s-like Amyloid Pathology Displays Pre-Plaque Intracellular Amyloid-Beta-Associated Cognitive Impairment. J. Alzheimers Dis. 2010, 20, 113–126. [Google Scholar] [CrossRef] [Green Version]
- Yang, S. Patterns of Insertions and Their Covariation with Substitutions in the Rat, Mouse, and Human Genomes. Genome Res. 2004, 14, 517–527. [Google Scholar] [CrossRef] [Green Version]
- Cohen, R.M.; Rezai-Zadeh, K.; Weitz, T.M.; Rentsendorj, A.; Gate, D.; Spivak, I.; Bholat, Y.; Vasilevko, V.; Glabe, C.G.; Breunig, J.J.; et al. A Transgenic Alzheimer Rat with Plaques, Tau Pathology, Behavioral Impairment, Oligomeric Aβ, and Frank Neuronal Loss. J. Neurosci. Off. J. Soc. Neurosci. 2013, 33, 6245–6256. [Google Scholar] [CrossRef]
- Kavkova, M.; Zikmund, T.; Kala, A.; Salplachta, J.; Proskauer Pena, S.L.; Kaiser, J.; Jezek, K. Contrast Enhanced X-Ray Computed Tomography Imaging of Amyloid Plaques in Alzheimer Disease Rat Model on Lab Based Micro CT System. Sci. Rep. 2021, 11, 5999. [Google Scholar] [CrossRef] [PubMed]
- Pentkowski, N.S.; Berkowitz, L.E.; Thompson, S.M.; Drake, E.N.; Olguin, C.R.; Clark, B.J. Anxiety-like Behavior as an Early Endophenotype in the TgF344-AD Rat Model of Alzheimer’s Disease. Neurobiol. Aging 2018, 61, 169–176. [Google Scholar] [CrossRef] [PubMed]
- Saré, R.M.; Cooke, S.K.; Krych, L.; Zerfas, P.M.; Cohen, R.M.; Smith, C.B. Behavioral Phenotype in the TgF344-AD Rat Model of Alzheimer’s Disease. Front. Neurosci. 2020, 14, 601. [Google Scholar] [CrossRef] [PubMed]
- Allison, S.L.; Fagan, A.M.; Morris, J.C.; Head, D. Spatial Navigation in Preclinical Alzheimer’s Disease. J. Alzheimers Dis. 2016, 52, 77–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coughlan, G.; Laczó, J.; Hort, J.; Minihane, A.-M.; Hornberger, M. Spatial Navigation Deficits—Overlooked Cognitive Marker for Preclinical Alzheimer Disease? Nat. Rev. Neurol. 2018, 14, 496–506. [Google Scholar] [CrossRef]
- Hort, J.; Laczo, J.; Vyhnalek, M.; Bojar, M.; Bures, J.; Vlcek, K. Spatial Navigation Deficit in Amnestic Mild Cognitive Impairment. Proc. Natl. Acad. Sci. USA 2007, 104, 4042–4047. [Google Scholar] [CrossRef] [Green Version]
- Serino, S.; Morganti, F.; Di Stefano, F.; Riva, G. Detecting Early Egocentric and Allocentric Impairments Deficits in Alzheimer’s Disease: An Experimental Study with Virtual Reality. Front. Aging Neurosci. 2015, 7, 88. [Google Scholar] [CrossRef] [Green Version]
- Eichenbaum, H. The Role of the Hippocampus in Navigation Is Memory. J. Neurophysiol. 2017, 117, 1785–1796. [Google Scholar] [CrossRef] [Green Version]
- Morris, R.G.M.; Garrud, P.; Rawlins, J.N.P.; O’Keefe, J. Place Navigation Impaired in Rats with Hippocampal Lesions. Nature 1982, 297, 681–683. [Google Scholar] [CrossRef]
- O’Keefe, J.; Nadel, L. The Hippocampus as a Cognitive Map; Oxford University Press: Oxford, NY, USA, 1978; ISBN 978-0-19-857206-0. [Google Scholar]
- Fox, N.C.; Warrington, E.K.; Freeborough, P.A.; Hartikainen, P.; Kennedy, A.M.; Stevens, J.M.; Rossor, M.N. Presymptomatic Hippocampal Atrophy in Alzheimer’s Disease: A Longitudinal MRI Study. Brain 1996, 119, 2001–2007. [Google Scholar] [CrossRef]
- Hyman, B.; Van Hoesen, G.; Damasio, A.; Barnes, C. Alzheimer’s Disease: Cell-Specific Pathology Isolates the Hippocampal Formation. Science 1984, 225, 1168–1170. [Google Scholar] [CrossRef]
- Stoiljkovic, M.; Kelley, C.; Stutz, B.; Horvath, T.L.; Hajós, M. Altered Cortical and Hippocampal Excitability in TgF344-AD Rats Modeling Alzheimer’s Disease Pathology. Cereb. Cortex 2019, 29, 2716–2727. [Google Scholar] [CrossRef]
- Berkowitz, L.E.; Harvey, R.E.; Drake, E.; Thompson, S.M.; Clark, B.J. Progressive Impairment of Directional and Spatially Precise Trajectories by TgF344-AD Rats in the Morris Water Task. BioRxiv 2018, 8, 282392. [Google Scholar]
- Smith, L.A.; McMahon, L.L. Deficits in Synaptic Function Occur at Medial Perforant Path-Dentate Granule Cell Synapses Prior to Schaffer Collateral-CA1 Pyramidal Cell Synapses in the Novel TgF344-Alzheimer’s Disease Rat Model. Neurobiol. Dis. 2018, 110, 166–179. [Google Scholar] [CrossRef]
- Colgin, L.L.; Denninger, T.; Fyhn, M.; Hafting, T.; Bonnevie, T.; Jensen, O.; Moser, M.-B.; Moser, E.I. Frequency of Gamma Oscillations Routes Flow of Information in the Hippocampus. Nature 2009, 462, 353–357. [Google Scholar] [CrossRef] [PubMed]
- Jezek, K.; Henriksen, E.J.; Treves, A.; Moser, E.I.; Moser, M.-B. Theta-Paced Flickering between Place-Cell Maps in the Hippocampus. Nature 2011, 478, 246–249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zitricky, F.; Jezek, K. Retrieval of Spatial Representation on Network Level in Hippocampal CA3 Accompanied by Overexpression and Mixture of Stored Network Patterns. Sci. Rep. 2019, 9, 11512. [Google Scholar] [CrossRef] [Green Version]
- Cimadevilla, J.M.; Wesierska, M.; Fenton, A.A.; Bures, J. Inactivating One Hippocampus Impairs Avoidance of a Stable Room-Defined Place during Dissociation of Arena Cues from Room Cues by Rotation of the Arena. Proc. Natl. Acad. Sci. USA 2001, 98, 3531–3536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stuchlík, A.; Petrásek, T.; Prokopová, I.; Holubová, K.; Hatalová, H.; Valeš, K.; Kubík, S.; Dockery, C.; Wesierska, M. Place Avoidance Tasks as Tools in the Behavioral Neuroscience of Learning and Memory. Physiol. Res. 2013, 62, S1–S19. [Google Scholar] [CrossRef] [PubMed]
- Wesierska, M.; Dockery, C.; Fenton, A.A. Beyond Memory, Navigation, and Inhibition: Behavioral Evidence for Hippocampus-Dependent Cognitive Coordination in the Rat. J. Neurosci. 2005, 25, 2413–2419. [Google Scholar] [CrossRef] [Green Version]
- Muñoz-Moreno, E.; Tudela, R.; López-Gil, X.; Soria, G. Early Brain Connectivity Alterations and Cognitive Impairment in a Rat Model of Alzheimer’s Disease. Alzheimers Res. Ther. 2018, 10, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lester, A.W.; Moffat, S.D.; Wiener, J.M.; Barnes, C.A.; Wolbers, T. The Aging Navigational System. Neuron 2017, 95, 1019–1035. [Google Scholar] [CrossRef] [PubMed]
- Tsai, Y.; Lu, B.; Ljubimov, A.V.; Girman, S.; Ross-Cisneros, F.N.; Sadun, A.A.; Svendsen, C.N.; Cohen, R.M.; Wang, S. Ocular Changes in TgF344-AD Rat Model of Alzheimer’s Disease. Investig. Ophthalmol. Vis. Sci. 2014, 55, 523–534. [Google Scholar] [CrossRef] [PubMed]
- Donovan, N.J.; Locascio, J.J.; Marshall, G.A.; Gatchel, J.; Hanseeuw, B.J.; Rentz, D.M.; Johnson, K.A.; Sperling, R.A. Harvard Aging Brain Study Longitudinal Association of Amyloid Beta and Anxious-Depressive Symptoms in Cognitively Normal Older Adults. Am. J. Psychiatry 2018, 175, 530–537. [Google Scholar] [CrossRef] [PubMed]
- Lyketsos, C.G.; Carrillo, M.C.; Ryan, J.M.; Khachaturian, A.S.; Trzepacz, P.; Amatniek, J.; Cedarbaum, J.; Brashear, R.; Miller, D.S. Neuropsychiatric Symptoms in Alzheimer’s Disease. Alzheimers Dement 2011, 7, 532–539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kromer Vogt, L.J.; Hyman, B.T.; Van Hoesen, G.W.; Damasio, A.R. Pathological Alterations in the Amygdala in Alzheimer’s Disease. Neuroscience 1990, 37, 377–385. [Google Scholar] [CrossRef]
- España, J.; Giménez-Llort, L.; Valero, J.; Miñano, A.; Rábano, A.; Rodriguez-Alvarez, J.; LaFerla, F.M.; Saura, C.A. Intraneuronal Beta-Amyloid Accumulation in the Amygdala Enhances Fear and Anxiety in Alzheimer’s Disease Transgenic Mice. Biol. Psychiatry 2010, 67, 513–521. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Proskauer Pena, S.L.; Mallouppas, K.; Oliveira, A.M.G.; Zitricky, F.; Nataraj, A.; Jezek, K. Early Spatial Memory Impairment in a Double Transgenic Model of Alzheimer’s Disease TgF-344 AD. Brain Sci. 2021, 11, 1300. https://doi.org/10.3390/brainsci11101300
Proskauer Pena SL, Mallouppas K, Oliveira AMG, Zitricky F, Nataraj A, Jezek K. Early Spatial Memory Impairment in a Double Transgenic Model of Alzheimer’s Disease TgF-344 AD. Brain Sciences. 2021; 11(10):1300. https://doi.org/10.3390/brainsci11101300
Chicago/Turabian StyleProskauer Pena, Stephanie L., Konstantinos Mallouppas, Andre M. G. Oliveira, Frantisek Zitricky, Athira Nataraj, and Karel Jezek. 2021. "Early Spatial Memory Impairment in a Double Transgenic Model of Alzheimer’s Disease TgF-344 AD" Brain Sciences 11, no. 10: 1300. https://doi.org/10.3390/brainsci11101300