The Influence of Temperature on Metabolisms of Phosphorus Accumulating Organisms in Biological Wastewater Treatment Plants in the Presence of Cu(II) Toxicity


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sequencing Batch Reactor (SBR) Pilot Plant
2.2. Batch Experiments
2.3. Analytical Methods
3. Results and Discussion
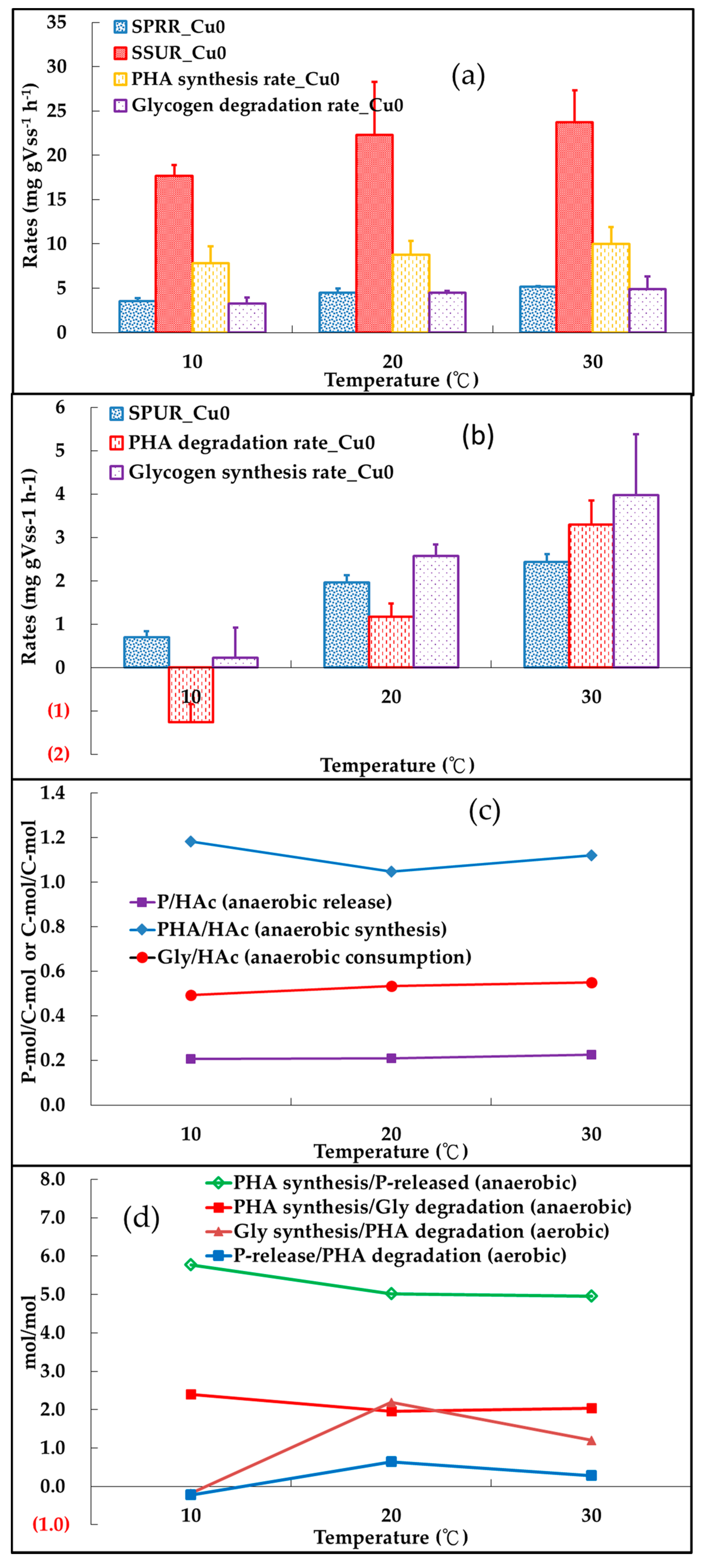
3.1. Background-Temperature Effect on Enhanced Biological Phosphorus Removal (EBPR) without Cu(II)
3.1.1. Anaerobic Metabolisms
3.1.2. Aerobic Metabolisms
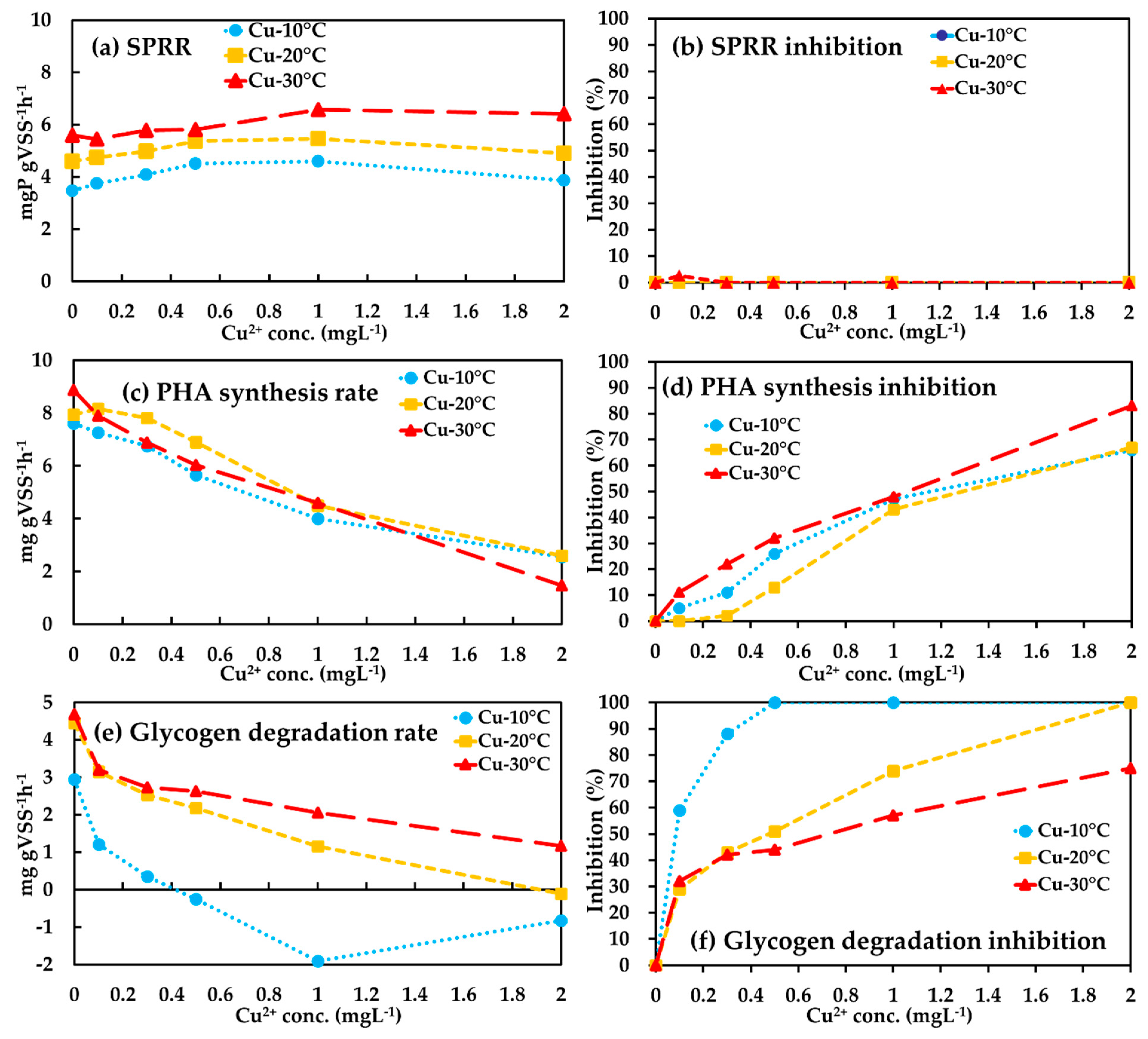
3.2. Temperature Influences on Anaerobic Metabolisms of Phosphorus Accumulating Organisms (PAOs) in Conjunction with Cu(II) Presence
3.2.1. Anaerobic Phosphate Release
3.2.2. Anaerobic Polyhydroxyalkanoate (PHA) Synthesis
3.2.3. Anaerobic Glycogen Degradation
3.3. Temperature Influences on Aerobic Metabolisms of PAOs in Conjunction with Cu(II) Presence
3.3.1. Aerobic Phosphate Uptake
3.3.2. Aerobic PHA Degradation
3.3.3. Aerobic Glycogen Replenishment
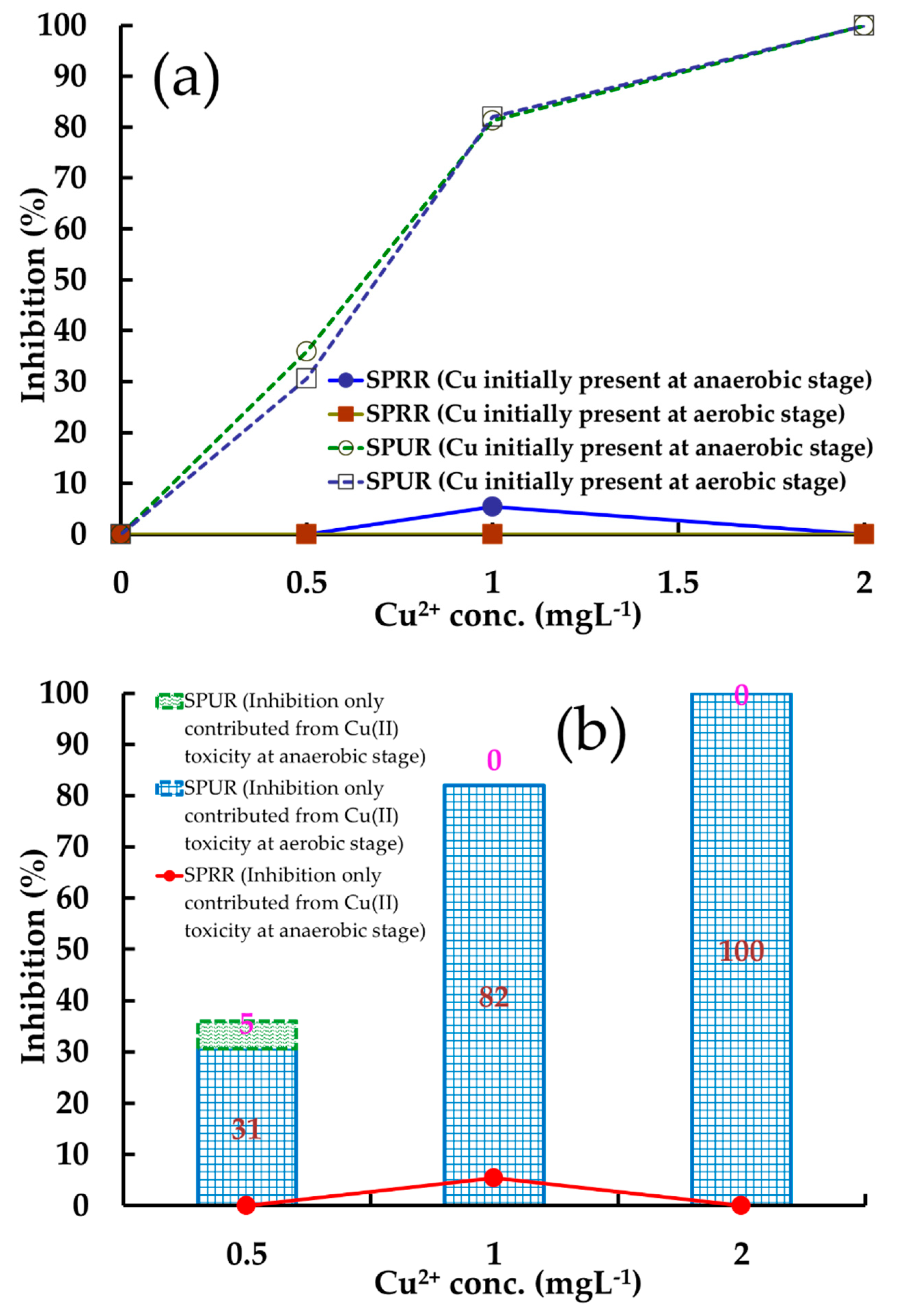
3.4. Identification of Cu(II) Inhibition Extents in Anaerobic and Aerobic Stages
3.4.1. Inhibition Causes of Phosphorus Metabolism
3.4.2. Inhibition Causes in PHA Metabolism
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Gebremariam, S.Y.; Beutel, M.W.; Christian, D.; Hess, T.F. Research Advances and Challenges in the Microbiology of Enhanced Biological Phosphorus Removal—A Critical Review. Water Environ. Res. 2011, 83, 195–219. [Google Scholar] [CrossRef]
- Neethling, J.B. Factors Influencing the Reliability of Enhanced Biological Phosphorus Removal; IWA Publishing: London, UK, 2006. [Google Scholar] [CrossRef]
- You, S.-J.; Tsai, Y.-P.; Cho, B.-C.; Chou, Y.-H. Metabolic influence of lead on polyhydroxyalkanoates (PHA) production and phosphate uptake in activated sludge fed with glucose or acetic acid as carbon source. Bioresour. Technol. 2011, 102, 8165–8170. [Google Scholar] [CrossRef]
- Tsai, Y.P.; Wu, W.M. Estimating Biomass of Heterotrophic and Autotrophic Bacteria by our Batch Tests AU. Environ. Technol. 2005, 26, 601–614. [Google Scholar] [CrossRef]
- Tsai, Y.-P.; You, S.-J.; Pai, T.-Y.; Chen, K.-W. Effect of cadmium on composition and diversity of bacterial communities in activated sludges. Int. Biodeterior. Biodegrad. 2005, 55, 285–291. [Google Scholar] [CrossRef]
- Tsai, Y.P.; You, S.-J.; Pai, T.-Y.; Chen, K.-W. Effect of Cd(II) on Different Bacterial Species Present in a Single Sludge Activated Sludge Process for Carbon and Nutrient Removal. J. Environ. Eng. 2006, 132, 173–180. [Google Scholar] [CrossRef]
- You, S.-J.; Tsai, Y.-P.; Huang, R.-Y. Effect of heavy metals on nitrification performance in different activated sludge processes. J. Hazard. Mater. 2009, 165, 987–994. [Google Scholar] [CrossRef] [PubMed]
- You, S.-J.; Tsai, Y.P.; Huang, R.-Y. Effects of Heavy Metals on the Specific Ammonia and Nitrate Uptake Rates in Activated Sludge. Environ. Eng. Sci. 2009, 26, 1207–1215. [Google Scholar] [CrossRef]
- Rayne, S.; Carey, S.; Forest, K. Evidence for Tin Inhibition of Enhanced Biological Phosphorus Removal at a Municipal Wastewater Treatment Plant AU. J. Environ. Sci. Health Part A 2005, 40, 535–551. [Google Scholar] [CrossRef]
- Wang, Y.; Ren, Z.; Jiang, F.; Geng, J.; He, W.; Yang, J. Effect of copper ion on the anaerobic and aerobic metabolism of phosphorus-accumulating organisms linked to intracellular storage compounds. J. Hazard. Mater. 2011, 186, 313–319. [Google Scholar] [CrossRef]
- Tsai, Y.-P.; Tzeng, H.-F.; Lin, J.-W.; Lu, M.-S.; Lin, J.-Y. Verification of enzymes deterioration due to Cu(II) presence in an enhanced biological phosphorus removal system. Chemosphere 2013, 91, 602–607. [Google Scholar] [CrossRef]
- Tsai, Y.-P.; Chen, H.-T. Influence of sludge retention time on tolerance of copper toxicity for polyphosphate accumulating organisms linked to polyhydroxyalkanoates metabolism and phosphate removal. Bioresour. Technol. 2011, 102, 11043–11047. [Google Scholar] [CrossRef]
- Mulkerrins, D.; Dobson, A.D.W.; Colleran, E. Parameters affecting biological phosphate removal from wastewaters. Environ. Int. 2004, 30, 249–259. [Google Scholar] [CrossRef]
- Oehmen, A.; Lemos, P.C.; Carvalho, G.; Yuan, Z.; Keller, J.; Blackall, L.L.; Reis, M.A.M. Advances in enhanced biological phosphorus removal: From micro to macro scale. Water Res. 2007, 41, 2271–2300. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Ahn, C.; Whang, L.M.; Lee, E.K.; Lee, Y.O.; Probst, T.H.; Reichardt, R.N.; Brauer, J.W. Problems in Dairy Wastewater Treatment; Water Environment Federation: Alexandria, VA, USA, 2001; Volume 2001, pp. 7–31. [Google Scholar]
- Mamais, D.; Jenkins, D. The Effects of MCRT and Temperature on Enhanced Biological Phosphorus Removal. Water Sci. Technol. 1992, 26, 955–965. [Google Scholar] [CrossRef]
- McClintock, S.A.; Randall, C.W.; Pattarkine, V.M. Effects of temperature and mean cell residence time on biological phosphorus removal processes. Water Environ. Res. 1993, 65, 110–118. [Google Scholar] [CrossRef]
- Krichten, D.; Hong, S.; Tracy, K. Applied biological phosphorus removal technology for municipal wastewater treatment by the A/O process. In Proceedings of the International Conference on Management Strategies for Phosphorus in the Environment, Lisbon, Portugal, 1–4 July 1985; pp. 1–4. [Google Scholar]
- Vinconneau, J.; Hascoet, M.; Florentz, M. The first applications of biological phosphorus removal in France. In Proceedings of the International Conference on Management Strategies for Phosphorus in the Environment, Lisbon, Portugal, 1–4 July 1985. [Google Scholar]
- Converti, A.; Rovatti, M.; Del Borghi, M. Biological removal of phosphorus from wastewaters by alternating aerobic and anaerobic conditions. Water Res. 1995, 29, 263–269. [Google Scholar] [CrossRef]
- Florentz, M.; Caille, D.; Bourdon, F.; Sibony, J. Biological phosphorus removal in France. Water Sci. Technol. 1987, 19, 1171–1173. [Google Scholar] [CrossRef]
- Lopez-Vazquez, C.M.; Oehmen, A.; Hooijmans, C.M.; Brdjanovic, D.; Gijzen, H.J.; Yuan, Z.; van Loosdrecht, M.C.M. Modeling the PAO–GAO competition: Effects of carbon source, pH and temperature. Water Res. 2009, 43, 450–462. [Google Scholar] [CrossRef]
- Panswad, T.; Doungchai, A.; Anotai, J. Temperature effect on microbial community of enhanced biological phosphorus removal system. Water Res. 2003, 37, 409–415. [Google Scholar] [CrossRef]
- Whang, L.-M.; Park, J.K. Competition between polyphosphate-and glycogen-accumulating organisms in enhanced-biological-phosphorus-removal systems: Effect of temperature and sludge age. Water Environ. Res. 2006, 78, 4–11. [Google Scholar] [CrossRef]
- Ren, N.; Kang, H.; Wang, X.; Li, N. Short-term effect of temperature variation on the competition between PAOs and GAOs during acclimation period of an EBPR system. Front. Environ. Sci. Eng. 2011, 5, 277–282. [Google Scholar] [CrossRef]
- Ahn, Y.H.; Speece, R.E. Elutriated acid fermentation of municipal primary sludge. Water Res. 2006, 40, 2210–2220. [Google Scholar] [CrossRef]
- American Public Health Association. Standard Methods for the Examination of Water and Wastewater; American Public Health Association: Washington, DC, USA, 1998. [Google Scholar]
- Chuang, S.H.; Ouyang, C.F.; Yuang, H.C.; You, S.J. Phosphorus and polyhydroxyalkanoates variation in a combined process with activated sludge and biofilm. Water Sci. Technol. 1998, 37, 593–597. [Google Scholar] [CrossRef]
- Sudiana, I.M.; Mino, T.; Satoh, H.; Nakamura, K.; Matsuo, T. Metabolism of enhanced biological phosphorus removal and non-enhanced biological phosphorus removal sludge with acetate and glucose as carbon source. Water Sci. Technol. 1999, 39, 29–35. [Google Scholar] [CrossRef]
- Smolders, G.J.F.; van der Meij, J.; van Loosdrecht, M.C.M.; Heijnen, J.J. Model of the anaerobic metabolism of the biological phosphorus removal process: Stoichiometry and pH influence. Biotechnol. Bioeng. 1994, 43, 461–470. [Google Scholar] [CrossRef]
- Brdjanovic, D.; van Loosdrecht, M.; Hooijmans, C.M.; Alaerts, G.; Heijnen, S. Temperature effects on physiology of biological phosphorus removal. J. Environ. Eng. 1997, 123, 144–154. [Google Scholar] [CrossRef]
- Mino, T.; van Loosdrecht, M.; Heijnen, S. Microbiology and biochemistry of the enhanced biological phosphate removal process. Water Res. 1998, 32, 3193–3207. [Google Scholar] [CrossRef]
- Matias, H.; Lemos, P.; Reis, M.; Crespo, J.; Carrondo, M.; Santos, H. Model for carbon metabolism in biological phosphorus removal processes based on in vivo 13C-NMR labelling experiments. Water Res. 1996, 30, 2128–2138. [Google Scholar]





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chou, C.-C.; Yang, C.-C.; Lu, M.-S.; Hu, L.-Y.; Chen, K.-F.; Tsai, Y.-P. The Influence of Temperature on Metabolisms of Phosphorus Accumulating Organisms in Biological Wastewater Treatment Plants in the Presence of Cu(II) Toxicity. Appl. Sci. 2019, 9, 1126. https://doi.org/10.3390/app9061126
Chou C-C, Yang C-C, Lu M-S, Hu L-Y, Chen K-F, Tsai Y-P. The Influence of Temperature on Metabolisms of Phosphorus Accumulating Organisms in Biological Wastewater Treatment Plants in the Presence of Cu(II) Toxicity. Applied Sciences. 2019; 9(6):1126. https://doi.org/10.3390/app9061126
Chicago/Turabian StyleChou, Chen-Chiang, Chih-Chi Yang, Meng-Shan Lu, Li-Yuan Hu, Ku-Fan Chen, and Yung-Pin Tsai. 2019. "The Influence of Temperature on Metabolisms of Phosphorus Accumulating Organisms in Biological Wastewater Treatment Plants in the Presence of Cu(II) Toxicity" Applied Sciences 9, no. 6: 1126. https://doi.org/10.3390/app9061126