Enhanced Production of Carboxymethylcellulase by Recombinant Escherichia coli Strain from Rice Bran with Shifts in Optimal Conditions of Aeration Rate and Agitation Speed on a Pilot-Scale


Abstract
:Featured Application
Abstract
1. Introduction
2. Materials and Methods
2.1. Medium and Bacterial Strain
2.2. Production of CMCase by E. coli KACC 91335P
2.3. Analytical Methods
2.4. Statistical Analysis
3. Results and Discussion
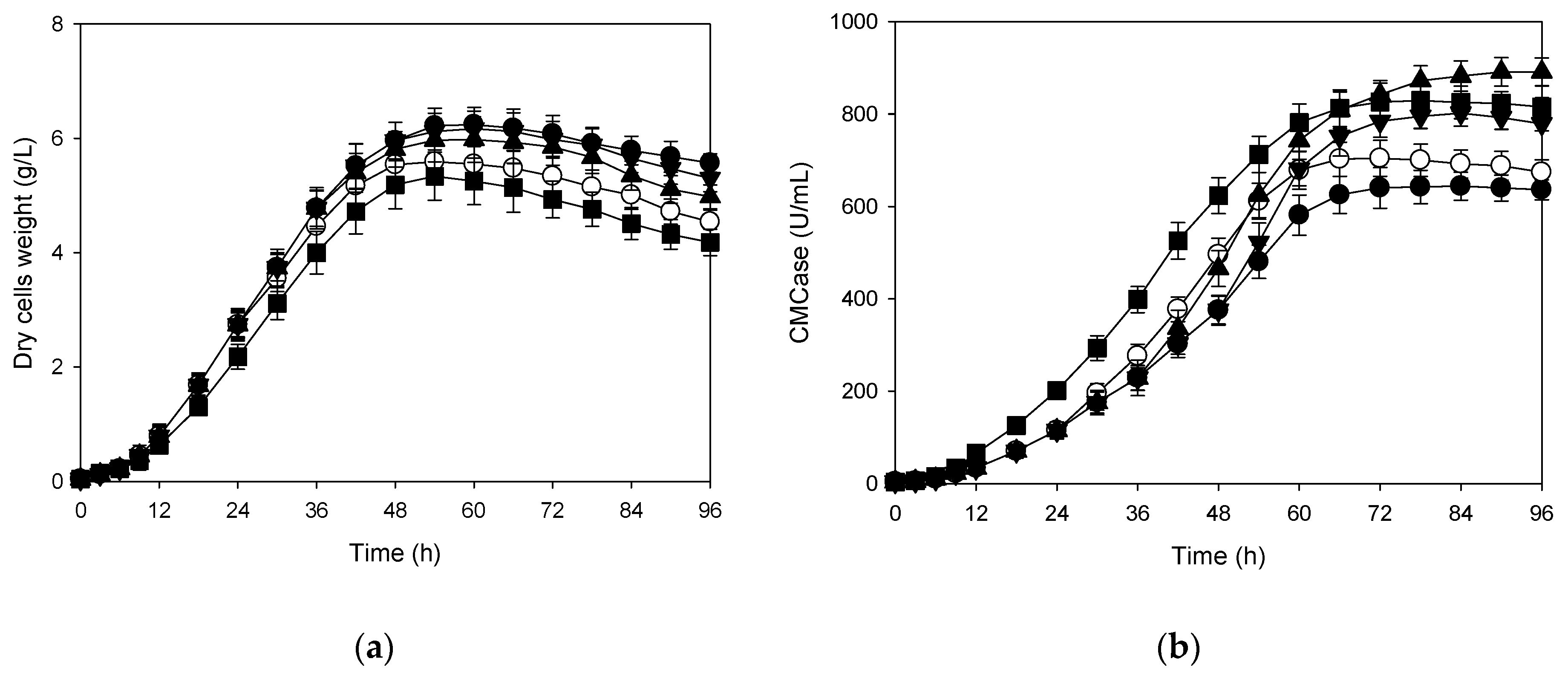
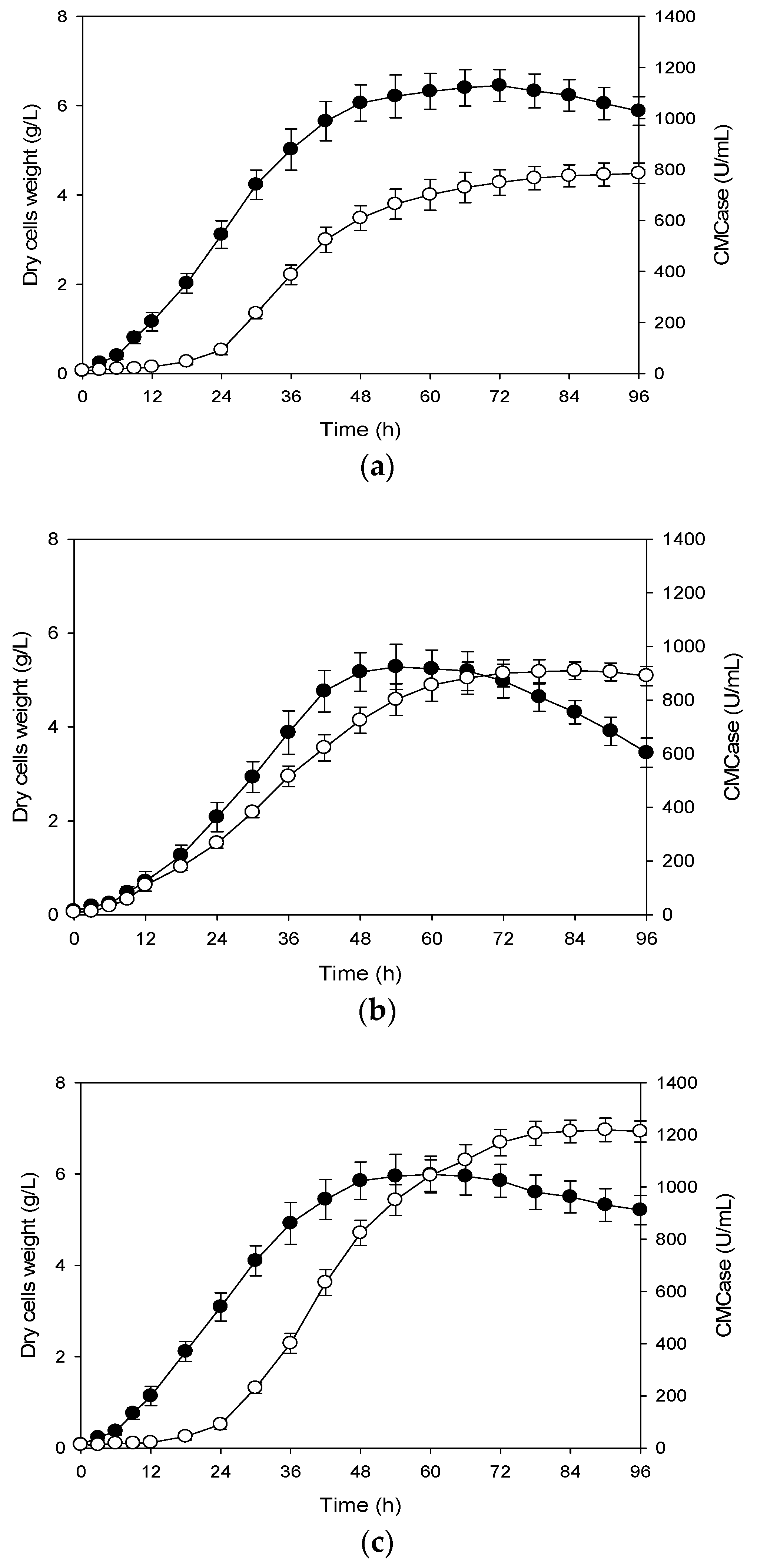
3.1. Effects of Shift in Aeration Rate on Production of CMCase
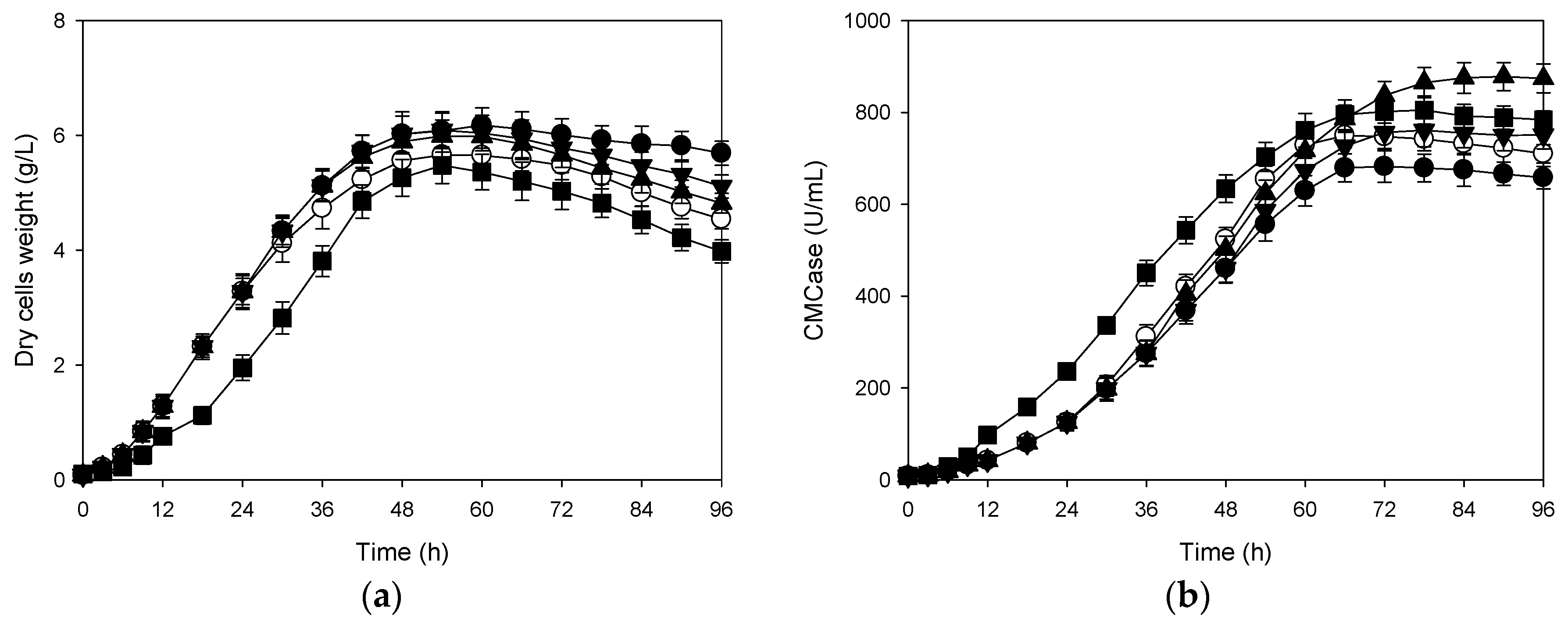
3.2. Effects of Shift in Agitation Speed on Production of CMCase
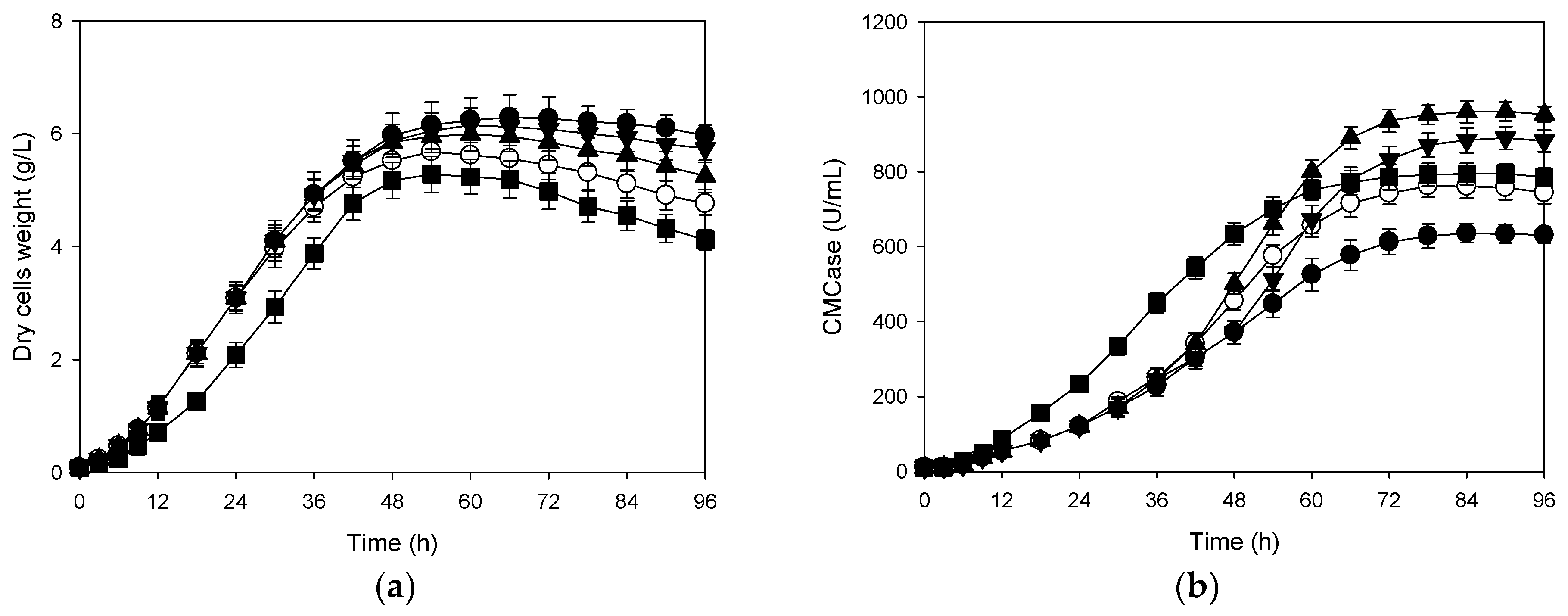
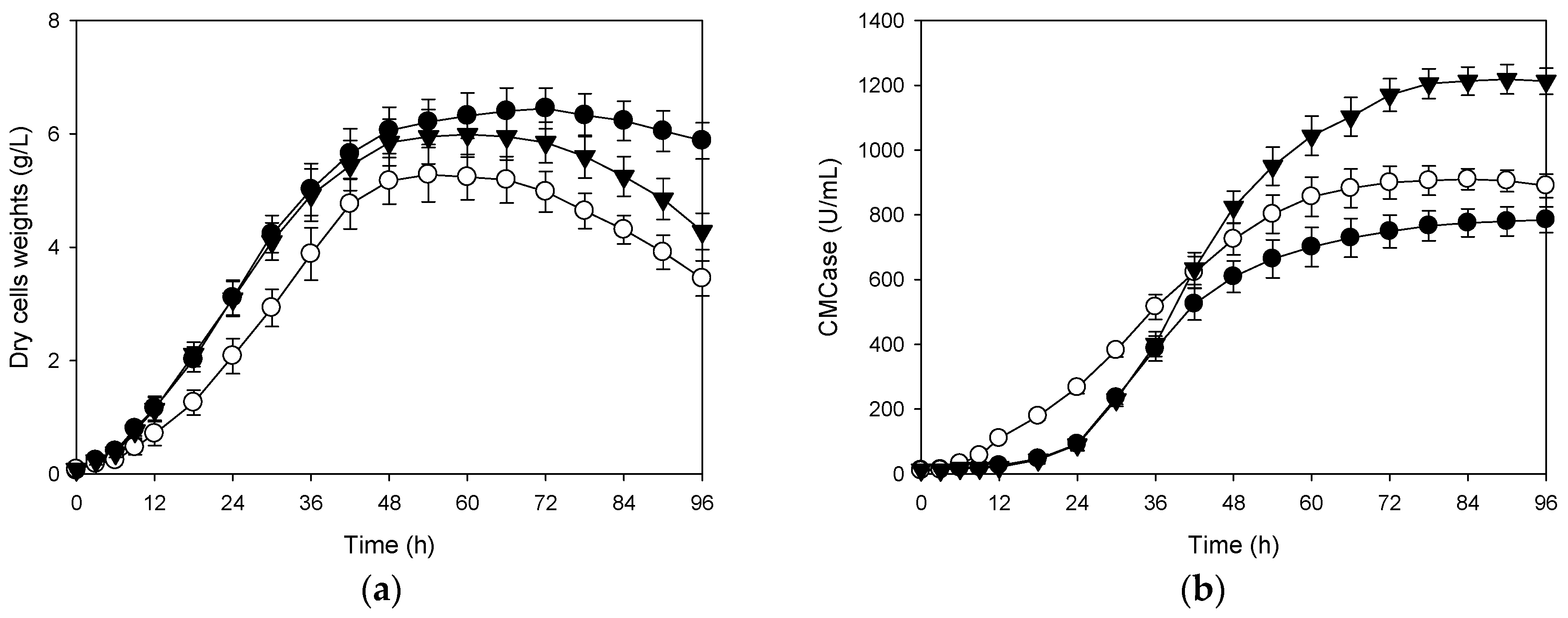
3.3. Effects of Shifts in Aeration Rate and Agitation Speed on Production of CMCase
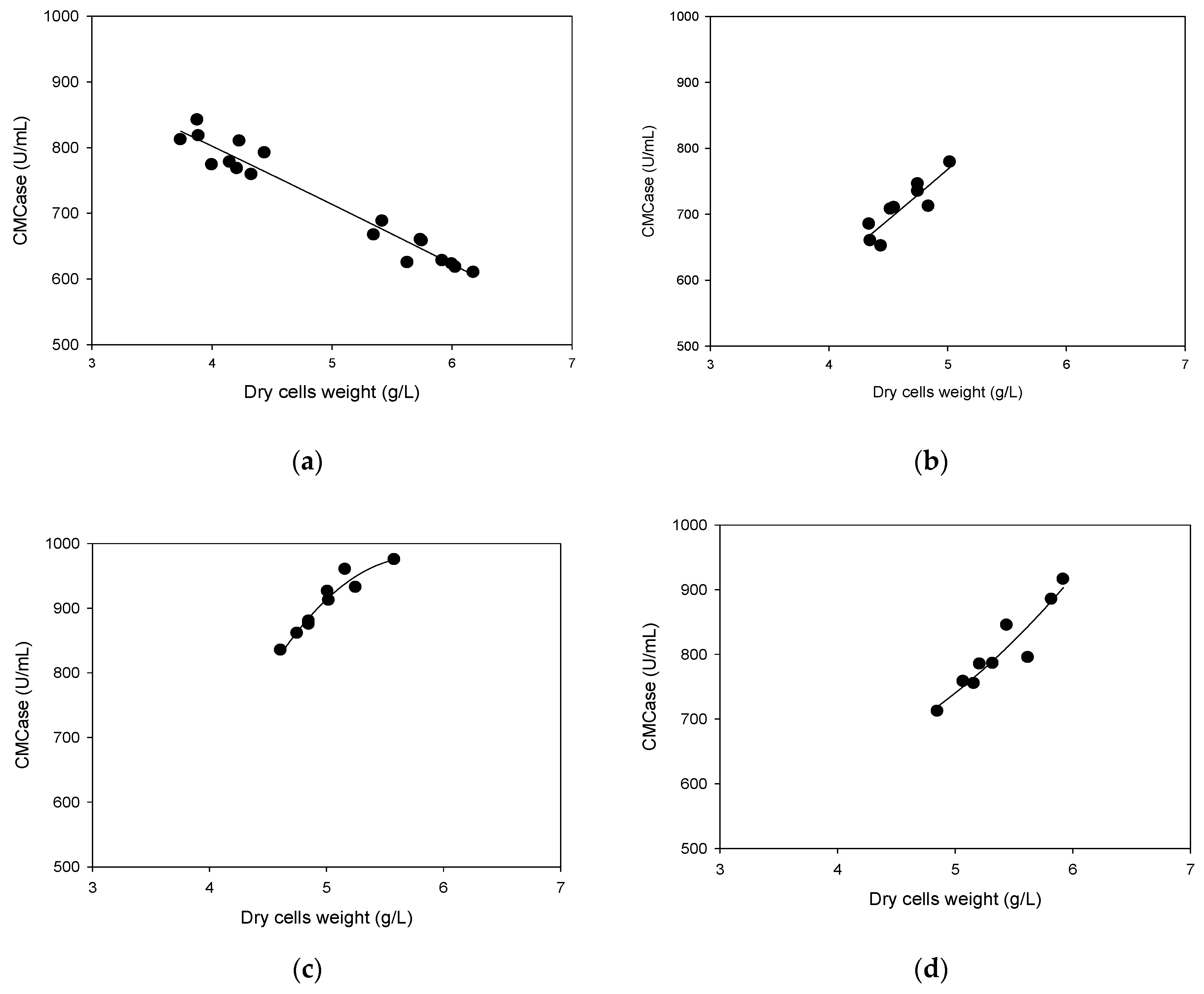
3.4. Correlation Between Cells Growth and Its Production of CMCase
3.5. Production of CMCase with Shifts in Aeration Rate and Agitation Speed of Pilot-Scale Bioreactor
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kim, H.J.; Lee, Y.J.; Gao, W.; Chung, C.H.; Son, C.W.; Lee, J.W. Statistical optimization of fermentation conditions and comparison of their influences on production of cellulases by psychrophilic marine bacterium, Psychrobacter aquimaris LBH-10 using orthogonal array method. Biotechnol. Bioprocess Eng. 2011, 16, 542–548. [Google Scholar] [CrossRef]
- Lee, E.J.; Lee, B.H.; Kim, B.K.; Lee, J.W. Enhanced production of carboxymethylcellulase of a marine microorganism, Bacillus subtilis subsp. subtilis A-53 in a pilot-scaled bioreactor by a recombinant Escherichia coli JM109/A-53 from rice bran. Mol. Biol. Rep. 2013, 40, 3609–3621. [Google Scholar] [CrossRef] [PubMed]
- Park, J.W.; Park, K.G.; Lee, N.Y.; Lee, J.H.; Lee, J.W. Enhanced extraction of reducing sugar from fruit of Hovenia dulcis with treatment of cellulase and sequential production of ethanol and acetic acid containing ampelopsin from extracted reducing sugars. Ind. Crops Prod. 2019, 139. in press. [Google Scholar] [CrossRef]
- Wei, G.Y.; Lee, Y.J.; Kim, Y.J.; Jin, I.H.; Lee, J.H.; Chung, C.H.; Lee, J.W. Kinetic study on the pretreatment and enzymatic saccharification on rice hull for the production of fermentable sugars. Appl. Biochem. Biotechnol. 2010, 162, 1471–1482. [Google Scholar] [CrossRef] [PubMed]
- Gao, W.; Lee, E.J.; Lee, S.U.; Lee, J.H.; Chung, C.H.; Lee, J.W. Enhanced carboxymethylcellulases production by a newly isolated marine bacterium Cellulophaga lytica LBH-14 from rice bran using response surface method. J. Microbiol. Biotechnol. 2012, 22, 1415–1425. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Gao, W.; Chung, C.H.; Lee, J.W. Statistical optimization for production of carboxymethylcellulase from rice hulls by a new isolated marine microorganism Bacillus licheniformis LBH-52 using response surface method. J. Life Sci. 2011, 21, 1083–1093. [Google Scholar] [CrossRef]
- Gao, W.; Kim, H.J.; Chung, C.H.; Lee, J.W. Enhanced production of carboxymethylcellulase by a marine bacterium, Bacillus velezensis A-68 by using rice hulls in pilot-scale bioreactor under optimized conditions for dissolved oxygen. J. Microbiol. 2014, 52, 755–761. [Google Scholar] [CrossRef]
- Lee, N.K.; Seo, H.P.; Cho, Y.B.; Son, C.W.; Gao, W.; Lee, J.W. Enhanced production of gellan by Sphingomonas paucibilis NK-200 with shifts in agitation speed and aeration rate after glucose feeding into the medium. J. Life Sci. 2010, 20, 811–818. [Google Scholar] [CrossRef]
- Jung, D.Y.; Son, C.W.; Kim, S.K.; Gao, W.; Lee, J.W. Enhanced production of heteropolysaccharide-7 by Beijerinckia indica HS-2001 in pilot-scaled bioreactor under optimized conditions involved in dissolved oxygen using sucrose-based medium. Biotechnol. Bioprocess Eng. 2013, 18, 94–103. [Google Scholar] [CrossRef]
- Lee, Y.J.; Kim, H.J.; Gao, W.; Chung, C.H.; Lee, J.W. Statistical optimization for production of carboxymethylcellulase of Bacillus amyloliquefaciens DL-3 by a recombinant Escherichia coli JM109/DL-3 from rice bran using response surface method. Biotechnol. Bioprocess Eng. 2012, 17, 227–235. [Google Scholar] [CrossRef]
- Cao, W.; Lee, S.U.; Li, J.; Lee, J.W. Enhanced production of carboxymethylcellulase by Cellulophaga lytica LBH-14 in pilot-scale bioreactor under optimized conditions involved in dissolved oxygen. Korean J. Chem. Eng. 2013, 30, 1105–1110. [Google Scholar] [CrossRef]
- Gao, W.; Chung, C.H.; Lee, J.H.; Lee, J.W. Enhanced production of cellobiase by marine bacterium Cellulophaga lytica LBH-14 from rice bran under optimized conditions involved in dissolved oxygen. Biotechnol. Bioprocess Eng. 2015, 20, 131–138. [Google Scholar] [CrossRef]
- Song, P.; Chen, C.; Tian, Q.; Lin, M.; Huang, H.; Li, S. Two-stage oxygen supply strategy for enhanced lipase production by Bacillus subtilis based on metabolic analysis. Biochem. Eng. J. 2013, 71, 1–10. [Google Scholar] [CrossRef]
- Wang, Y.H.; Fang, X.L.; Li, Y.P.; Zhang, X. Effects of constant and shifting dissolved oxygen concentration on the growth and antibiotic activity of Xenorhadus nematophila. Bioresour. Technol. 2010, 101, 7529–7536. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.J.; Gao, W.; Lee, J.W. Enhanced production of carboxymethylcellulase of Bacillus subtilis subsp. subtilis A-53 by a recombinant Escherichia coli JM109/A-53 with pH and temperature shifts. Korean J. Chem. Eng. 2015, 32, 113–117. [Google Scholar] [CrossRef]
- Kim, M.H.; Gao, W.; Lee, J.W. Comparison of optimal conditions for mass production of carboxymethylcellulase by Escherichia coli JM109/A-68 with other recombinants in pilot-scale bioreactor. Biotechnol. Bioprocess Eng. 2017, 22, 142–149. [Google Scholar] [CrossRef]
- Kim, M.H.; Kang, D.U.; Lee, J.W. Construction of a recombinant Escherichia coli JM109/A-68 for production of carboxymethylcellulase and comparison of its production with its wild type, Bacillus velezensis A-68 in a pilot-scale bioreactor. Biotechnol. Bioprocess Eng. 2016, 21, 601–611. [Google Scholar] [CrossRef]
- Jo, K.I.; Lee, Y.J.; Kim, B.K.; Lee, B.H.; Chung, C.H.; Nam, S.W.; Kim, S.K.; Lee, J.W. Pilot-scale production of carboxymethylcellulase from rice hull by Bacillus amyloliquefaciens DL-3. Biotechnol. Bioprocess Eng. 2008, 13, 182–188. [Google Scholar] [CrossRef]
- Lee, S.U.; Gao, W.; Chung, C.H.; Lee, J.W. Construction of recombinant Escherichia coli JM109/LBH-10 and comparison of its optimal conditions for production of carboxymethylcellulase with its wild type, Psychrobacter aquimaris LBH-10. J. Microb. Biochem. Technol. 2014, 3, 135–143. [Google Scholar] [CrossRef]
- Shuler, M.L.; Kargi, F. Bioprocess Engineering Basic Concepts; Prentice Hall: Upper Saddle River, NJ, USA, 2002; pp. 166–168. [Google Scholar]
- Lawrence, R.A. A pocket calculator program for Duncan’s new multiple ranged test and analysis of vatiance. Comput. Biol. Med. 1984, 14, 357–362. [Google Scholar] [CrossRef]
- Fernández, A.; Ocio, M.J.; Fernández, P.S.; Rodrigo, M.; Martinez, A. Application of nonlinear regression analysis to the estimation of kinetic parameters for two enterotoxigenic strains of Bacillus cereus spores. Food Microbiol. 1999, 16, 607–613. [Google Scholar] [CrossRef]
- Zwietering, M.H.; Jongenburger, I.; Rombout, F.M.; van’t Riet, K. Modeling of the bacterial growth curve. Appl. Environ. Microbiol. 1990, 56, 1875–1881. [Google Scholar] [PubMed]
- Yates, G.T.; Smotzer, T. On the lag phase and initial decline of microbial growth curve. J. Theor. Biol. 2007, 244, 511–517. [Google Scholar] [CrossRef]
- Lee, B.H.; Kim, B.K.; Lee, Y.J.; Chung, C.H.; Lee, J.W. Industrial scale of optimization for the production of carboxymethylcellulase from rice bran by a marine bacterium, Bacillus subtilis subsp. subtilis A-53. Enzym. Microb. Technol. 2010, 46, 38–42. [Google Scholar] [CrossRef]
- Kim, B.K.; Kim, H.J.; Lee, J.W. Rapid statistical optimization of cultural conditions for mass production of carboxymethylcellulase by a newly isolated marine bacterium, Bacillus velezensis A-68 from rice hulls. J. Life Sci. 2013, 23, 757–769. [Google Scholar] [CrossRef]
- Heathman, T.R.J.; Nienow, A.W.; Rafiq, Q.A.; Coopman, K.; Kara, B.; Hewitt, C.J. Agitation and aeration of stirred-bioreactors for the microcarrier culture of human mesenchymal stem cells and potential implications for large-scale bioprocess development. Biochem. Eng. J. 2018, 136, 9–17. [Google Scholar] [CrossRef] [Green Version]
- Clement, T.; Perez, M.; Mouret, J.R.; Sablayrolles, J.M.; Camarasa, C. Use of a continuous multistage bioreactors to mimic winemaking fermentation. Int. J. Food Microbiol. 2011, 150, 42–49. [Google Scholar] [CrossRef]
- Wu, W.J.; Zhang, A.H.; Peng, C.; Song, P.; Yu, Y.D.; Huang, H.; Ji, X.J. An efficient multi-stage fermentation strategy for the production of microbial oil rich in arachidonic acid in Mortierella alpine. Bioresour. Bioprocess. 2017, 4, 8. [Google Scholar] [CrossRef]
- Lee, J.H.; Lama, S.; Kim, J.R.; Park, S.H. Production of 1,3-propanedilo from glucose by recombinant Escherichia coli BL21 (DE3). Biotechnol. Bioprocess Eng. 2018, 23, 250–258. [Google Scholar] [CrossRef]
- Bayrock, D.P.; Ingledew, W.M. Application of multistage continuous fermentation for production of fuel alcohol by very-high-gravity fermentation technology. J. Ind. Microbiol. Biotechnol. 2001, 27, 87–93. [Google Scholar] [CrossRef]
- Lee, N.K. Statistical optimization of medium and fermentation conditions of recombinant Pichia pastoris for the production of xylanase. Biotechnol. Bioprocess Eng. 2018, 23, 55–63. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Condition | Aeration (vvm) | Agitation (rpm) | Time for Shift (h) | Dry Cells Weight (g/L) | CMCase (U/mL) |
|---|---|---|---|---|---|
| Shift in agitation | 0.95 | 500 | - | 5.69 ± 0.21 ae1,2 | 658 ± 25 ad |
| 0.95 | 500 to 416 | 24 | 4.54 ± 0.17 bf | 710 ± 20 ag | |
| 0.95 | 500 to 416 | 36 | 4.82 ± 0.17 bc | 874 ± 31 b | |
| 0.95 | 500 to 416 | 48 | 5.11 ± 0.20 cg | 751 ± 30 cg | |
| 0.95 | 416 | - | 3.98 ± 0.20 d | 784 ± 20 ce | |
| Shift in aeration | 0.50 | 416 | - | 5.67 ± 0.16 ae | 636 ± 22 d |
| 0.50 to 0.95 | 416 | 24 | 4.54 ± 0.21 bf | 674 ± 27 ad | |
| 0.50 to 0.95 | 416 | 36 | 4.95 ± 0.15 bcg | 891 ± 30 b | |
| 0.50 to 0.95 | 416 | 48 | 5.30 ± 0.23 ag | 779 ± 16 ce | |
| 0.95 | 416 | - | 4.18 ± 0.23 df | 815 ± 20.1 e | |
| Shifts in agitation and aeration | 0.50 | 500 | - | 5.97 ± 0.18 e | 631 ± 21 d |
| 0.50 to 0.95 | 500 to 416 | 24 | 4.76 ± 0.20 bc | 744 ± 29 cg | |
| 0.50 to 0.95 | 500 to 416 | 36 | 5.25 ± 0.24 ag | 953 ± 21 cf | |
| 0.50 to 0.95 | 500 to 416 | 48 | 5.74 ± 0.21 ae | 882 ± 29 b | |
| 0.95 | 416 | - | 4.12 ± 0.18 df | 785 ± 25 ce |
| Conditions and Results | Without Shift | With Shifts at 36 h | ||
|---|---|---|---|---|
| Conditions | Aeration (rpm) | 180 | 150 | 180 to 150 rpm |
| Agitation (vvm) | 0.50 | 0.95 | 0.50 to 0.95 vvm | |
| Results | DCW (g/L) | 5.88 | 3.45 | 5.21 |
| Cellulase (U/mL) | 785 | 890 | 1212 | |
| μ (/h) | 0.059 | 0.044 | 0.052 | |
| μmax (/h) | 0.092 | 0.083 | 0.103 | |
| dx/dt (g/L·h) | 0.061 | 0.035 | 0.054 | |
| dp/dt (U/mL·h) | 8.177 | 9.271 | 12.625 | |
| dp/dx (/U/g) | 133.50 | 257.97 | 232.63 | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, C.-I.; Lee, J.-H.; Li, J.; Lee, J.-W. Enhanced Production of Carboxymethylcellulase by Recombinant Escherichia coli Strain from Rice Bran with Shifts in Optimal Conditions of Aeration Rate and Agitation Speed on a Pilot-Scale. Appl. Sci. 2019, 9, 4083. https://doi.org/10.3390/app9194083
Park C-I, Lee J-H, Li J, Lee J-W. Enhanced Production of Carboxymethylcellulase by Recombinant Escherichia coli Strain from Rice Bran with Shifts in Optimal Conditions of Aeration Rate and Agitation Speed on a Pilot-Scale. Applied Sciences. 2019; 9(19):4083. https://doi.org/10.3390/app9194083
Chicago/Turabian StylePark, Chung-Il, Jae-Hong Lee, Jianhong Li, and Jin-Woo Lee. 2019. "Enhanced Production of Carboxymethylcellulase by Recombinant Escherichia coli Strain from Rice Bran with Shifts in Optimal Conditions of Aeration Rate and Agitation Speed on a Pilot-Scale" Applied Sciences 9, no. 19: 4083. https://doi.org/10.3390/app9194083