Increased Anti-Inflammatory Effects on LPS-Induced Microglia Cells by Spirulina maxima Extract from Ultrasonic Process

Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of the S. maxima Extracts
2.2. Measurement of the Chlorophyll Contents in the Extracts
- C: Chlorophyll (mg/L) = 8.970 × (7.19 × A419 nm + 3.33 × A454 nm)
- S: Weight (mg) of the extracted sample in 2 mL of alkaline pyridine solution
- A419 nm: Absorbance at a wavelength of 419 nm
- A454 nm: Absorbance at a wavelength of 454 nm
2.3. Measurement of Cell Cytotoxicity and the Production of Nitric Oxide (NO) and Prostaglandin E2 (PGE2)
2.4. Reverse Transcription Polymerase Chain Reaction (RT-PCR)
2.5. Measurement of the Secretioin of Pro-Inflammatory Cytokines from BV-2 Cells
2.6. Statistical Analysis
3. Results and Discussion
3.1. Total Chlorophyll Concentrations in the Extracts from Two Different Extraction Processes
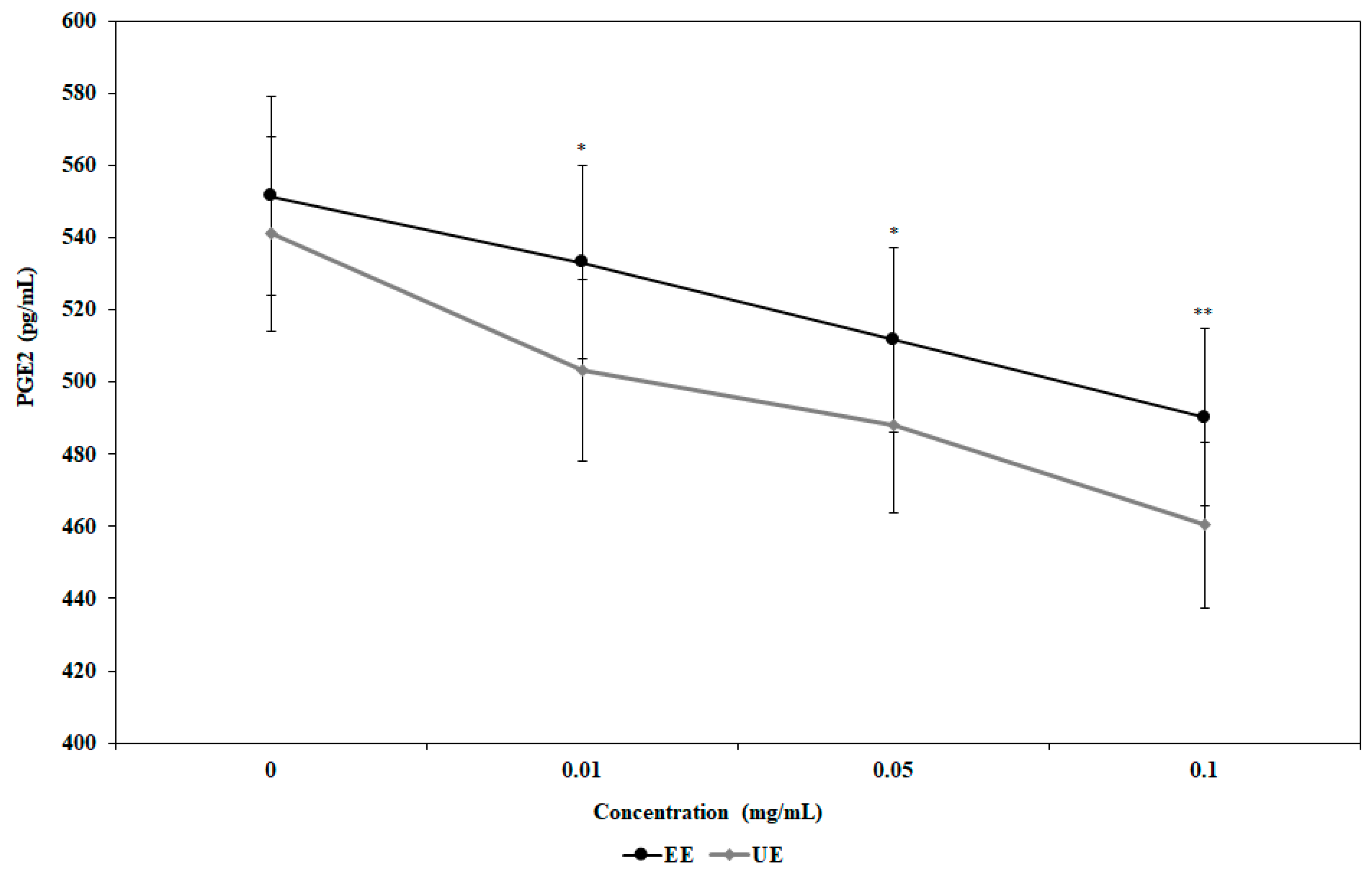
3.2. Inhibition of Nitric Oxide (NO) and Prostaglandin E2 (PGE2) Production by the Extracts
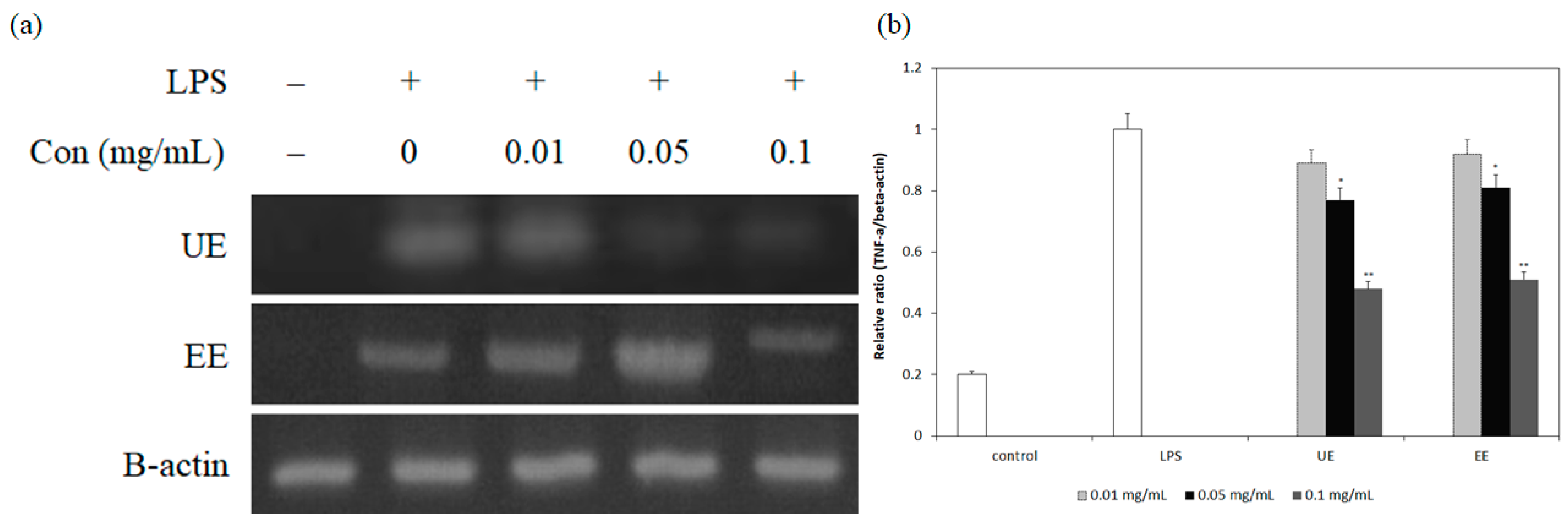
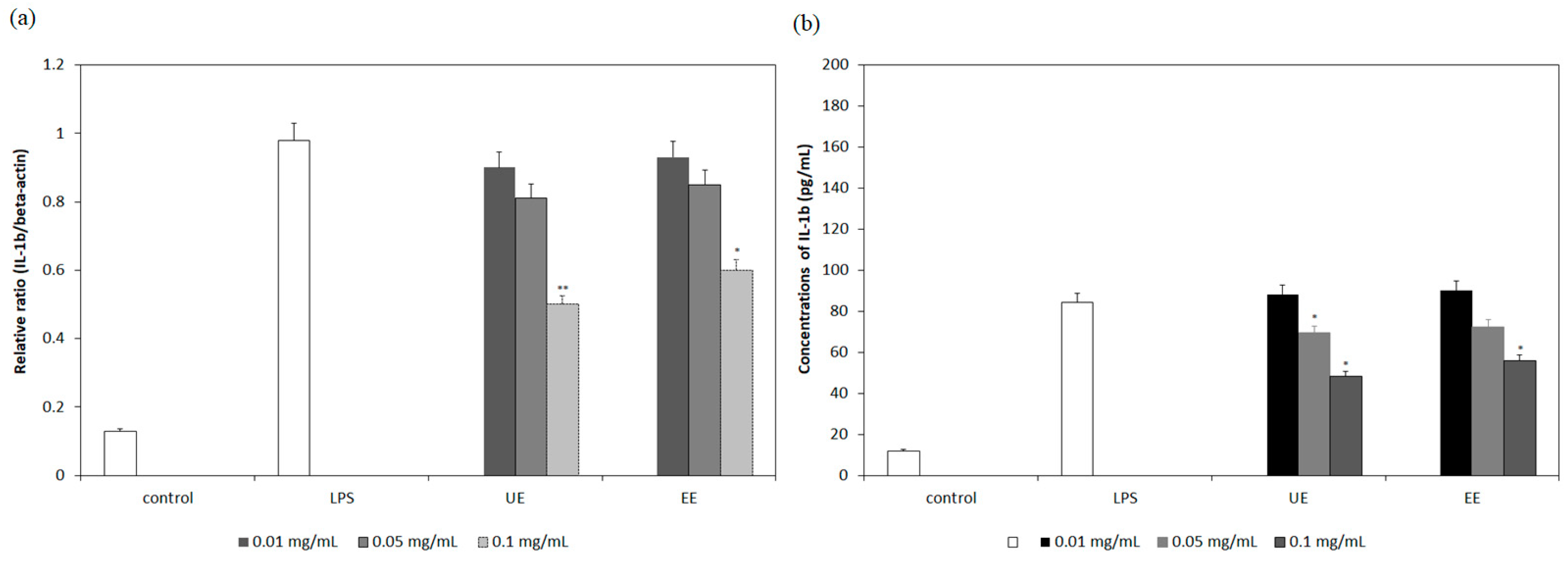
3.3. Inhibition of mRNA Expression and Secretion of Pro-Inflammatory Cytokines by the Extracts
3.4. Correlation Between Gene Down-Regulation and Secretion of Pro-Inflammatory Cytokines
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Bartus, R.T.; Dean, R.L.; Beer, D.; Lipa, A.S. The cholinergic hypothesis of geriatric memory dysfunction. Science 1982, 217, 408–417. [Google Scholar] [CrossRef]
- Citron, M. Alzheimer’s disease: Treatments in discovery and development. Nat. Neurosci. 2002, 5, 1055–1057. [Google Scholar] [CrossRef]
- Squire, L.R. Memory systems of the brain: A brief history and current perspective. Neurobiol. Learn. Mem. 2004, 82, 71–77. [Google Scholar] [CrossRef] [PubMed]
- Minghetti, L. Role of inflammation in neurodegenerative diseases. Curr. Opin. Neurol. 2005, 18, 315–321. [Google Scholar] [CrossRef]
- Asai, M.; Iwat, N.; Yoshikawa, A.; Aizaki, Y.; Ishiura, S.; Saido, T.C.; Maruyana, K. Berberine alters the processing of Alzheimer’s amyloid precursor protein to decrease Abeta secretion. Bioichem. Biophys. Res. Commun. 2007, 352, 498–502. [Google Scholar] [CrossRef] [PubMed]
- Floyd, R.A. Antioxidants, oxidative stress and degenerative neurological disorders. Proc. Soc. Exp. Biol. Med. 2001, 222, 236–245. [Google Scholar] [CrossRef]
- Halliwell, B.; Gutterridge, J.M. Role of free radicals and catalytic metal ions in human diseases: An overview. Methods Enzymol. 1990, 186, 1–85. [Google Scholar] [CrossRef] [PubMed]
- Boille, S.; Vande Vele, C.; Cleveland, D.W. ALS: A disease of motor neurons and their nonneuronal neighbors. Neuron 2006, 52, 130–144. [Google Scholar] [CrossRef]
- Talhouk, R.S.; Karam, C.; Fostok, S.; El-Jouni, W.; Barbour, E.K. Anti-inflammatory bioactivities in plant extracts. J. Med. Food 2007, 10, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, S.M.; Barone, S. The neurotoxicant trimethyltin induces apoptosis via caspase activation, p38 protein kinase and oxidative stress in PC12 cells. Toxicol. Lett. 2004, 147, 63–72. [Google Scholar] [CrossRef] [PubMed]
- Ali, S.F.; LeBel, C.P.; Bondy, S.C. Reactive oxygen species formation as a biomarker of methylmercury and trimethyltin neurotoxicity. Neurotoxicology 1992, 13, 637648. [Google Scholar] [PubMed]
- Corvino, V.; Marches, E.; Michetti, F.; Geloso, M.C. Neuroprotective strategies in hippocampal neurodegeneration induced by the neurotoxicant trimethyltin. Neurochem. Res. 2013, 38, 240–253. [Google Scholar] [CrossRef] [PubMed]
- Chen, A.; Xiong, L.J.; Tong, Y.; Mao, M. The neuroprotective roles of BDNF in hypoxic ischemic brain injury. Biomed. Rep. 2013, 1, 167–176. [Google Scholar] [CrossRef] [PubMed]
- Dawson, G.R.; Iversen, S.D. The effects of novel cholinesterase inhibitors and selective muscarinic receptor agonists in tests of reference and working memory. Behav. Brain Res. 1993, 57, 143–153. [Google Scholar] [CrossRef]
- Zhang, W.; Zhang, X.; Zou, K.; Wang, J.L.; Wang, Y. Seabackthorn berry polysaccharide protects against carbon tetrachloride-induced hepatoxicity in mice via anti-oxidative and anti-inflammatory activities. Food Funct. 2017, 8, 3130–3138. [Google Scholar] [CrossRef]
- Yuk, H.J.; Lee, J.W.; Park, H.A.; Seo, K.S.; Oh, S.R.; Ryu, H.W. Protective effects of coumestrol on lipopolysaccharide-induced acute lung injury via the inhibition of pro-inflammatory mediators and NF-kB activation. J. Funct. Food 2017, 34, 181–188. [Google Scholar] [CrossRef]
- Young, I.S.; Woodside, J.V. Antioxidants in health and disease. J. Clin. Pathol. 2001, 54, 176–186. [Google Scholar] [CrossRef] [Green Version]
- Bennett, P.B.; Marquis, R.E.; Demchenko, I. High Pressure Biology and Medicine; Bennett, P.B., Ed.; University of Rochester Press: New York, NY, USA, 1998; pp. 1–23. [Google Scholar]
- Lim, G.P.; Chu, T.; Yang, F.; Beech, W.; Frautschy, S.A.; Cole, G.M. The curry spice curcumin reduces oxidative damage and amyloid pathology in an Alzheimer transgenic mouse. J. Neurosci. 2001, 21, 8370–8377. [Google Scholar] [CrossRef]
- Murabe, Y.; Sano, Y.; Roisen, F.J. Morphological studies on neuralgia. VI. Postnatal development of microglial cell. Tissue Res. 1982, 225, 469–485. [Google Scholar] [CrossRef]
- Nakajima, K.; Kohsaka, D.; Raquel, E.G. Functional roles of microglia in the brain. Neurosci. Res. 1993, 17, 187–203. [Google Scholar] [CrossRef]
- Gonzalez-Scarano, F.; Baltuch, G. Microglia as mediators of inflammatory and degenerative diseases. Annl. Rev. Neurosci. 1999, 22, 219–240. [Google Scholar] [CrossRef]
- Witkamp, R.; Monshouwer, M. Signal transduction in inflammatory processes, current and future therapeutic targets: A mini review. Vet. Q. 2002, 22, 11–16. [Google Scholar] [CrossRef] [PubMed]
- Eikelenboom, P.; van Gool, W.A. Neuroinflammatory perspectives on the two faces of Alzheimer’s disease. J. Neural Transm. 2004, 11, 281–294. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.S.; Hong, J.S. Role of microglia in inflammation-mediated neurodegenerative diseases: Mechanisms and strategies for therapeutic intervention. J. Pharmacol. Exp. Ther. 2003, 304, 1–7. [Google Scholar] [CrossRef]
- Gao, H.M.; Liu, B.; Zhang, W.; Hong, J.S. Novel anti-inflammatory therapy for Parkinson’s disease. Trends Pharmacol. Sci. 2003, 24, 395–401. [Google Scholar] [CrossRef]
- Saeid, A.; Chojnacka, K.; Korczyński, M.; Korniewicz, D.; Dobrzański, Z. Biomass of Spirulina maxima enriched by biosorption process as a new feed supplement for swine. J. Appl. Phycol. 2013, 25, 667–675. [Google Scholar] [CrossRef]
- Son, M.H.; Park, K.H.; Choi, A.R.; Yoo, G.; In, M.J.; Kim, D.H.; Chae, H.J. Investigation of biological activities of enzymatic hydrolysate of Spirulina. J. Korean Soc. Food Sci. Nutr. 2009, 38, 136–141. [Google Scholar] [CrossRef]
- Hernandez, A.C.; Nieves, I.; Meckes, M.; Chamorro, G.; Barron, B.L. Antiviral activity of Spirulina maxima against Herpes simplex virus type 2. Antivir. Res. 2002, 56, 279–285. [Google Scholar] [PubMed]
- Zao, X.; Yang, Y.H.; Cho, Y.S.; Chun, H.K.; Song, K.B.; Kim, M.R. Quality characteristics of Spirulina maxima added salad dressing. East Asian Soc. Diet. Life 2005, 15, 292–299. [Google Scholar]
- Deng, R.; Chow, T.J. Hypolipidemic, antioxidant, and anti-inflammatory activities of microalgae Spirulina. Cardiovasc. Ther. 2010, 28, 33–45. [Google Scholar] [CrossRef]
- Koh, E.J.; Seo, Y.J.; Choi, J.; Lee, H.Y.; Kang, D.H.; Kim, K.J.; Lee, B.Y. Spirulina maxima extract prevents neurotoxicity via promoting activation of BDNF/CREB signaling pathways in neuronal cells and mice. Molecules 2017, 22, 1363. [Google Scholar] [CrossRef] [PubMed]
- Bhat, S. R Chlorophyll: The wonder pigment. Sci Rep. 2005, 42, 29–32. [Google Scholar]
- Hosikian, A.; Lim, S.; Halim, R.; Danquah, M.K. Chlorophyll Extraction from Microalgae: A Review on the Process Engineering Aspects. Int. J. Chem. Eng. 2010, 29, 1–11. [Google Scholar] [CrossRef]
- Koh, E.J. The Improvement of Cognitive Impairment and Neuroprotection Effects of Spirulina maxima Extract. Ph.D. Thesis, Cha University, Pangyo, Korea, 2018; pp. 1–125. [Google Scholar]
- Kay, R.A.; Barton, L.L. Microalgae as food and supplement. Crit. Rev. Food Sci. Nutr. 1991, 30, 555–573. [Google Scholar] [CrossRef] [PubMed]
- McCarty, M.F. The chlorophyll metabolite phytanic acid is a natural rexinoid—Potential for treatment and prevention of diabetes. Med. Hypotheses 2001, 56, 217–219. [Google Scholar] [CrossRef]
- Lee, Y.J.; Kim, S.H.; Kim, J.S.; Han, J.A.; Seo, H.J.; Lim, H.J.; Choi, S.Y. Studies on Simultaneous Determination of Chlorophyll a and b, Pheophorbide a, and β-Carotene in Chlorella and Spirulina Products. J. Food Hyg. Saf. 2005, 20, 141–146. [Google Scholar]
- Kim, W.I.; Chung, K.W.; Hong, I.K.; Park, K.A. Ultrasound Energy Effect on Solvent Extraction of Amaranth Seed Oil. J. Korean Ind. Eng. Chem. 2001, 12, 307–311, OSTI ID:20268268. [Google Scholar]
- Oh, S.H.; Ahn, J.H.; Kang, D.H.; Lee, H.Y. The effect of ultrasonificated extracts of Spirulina maxima on the anticancer activity. Mar. Biotechnol. 2011, 13, 205–214. [Google Scholar] [CrossRef]
- Lee, D.H.; Hong, J.H. Antioxidant activities of chlorella extracts and physicochemical characteristics of spray-dried Chlorella powders. Korean J. Food Preserv. 2015, 22, 591–597. [Google Scholar] [CrossRef]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Met. 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Lee, Y.H.; Jeon, S.H.; Kim, S.H.; Kim, C.Y.; Lee, S.J.; Koh, D.S.; Lim, Y.H.; Ha, K.S.; Shin, S.Y. A new synthetic chalcone derivative, 2-hydroxy-3′,5,5′-trimethoxychalcone (DK-139), suppresses the Toll-like receptor 4-mediated inflammatory response through inhibition of the Akt/NF-κB pathway in BV-2 microglial cells. Exp. Mol. Med. 2012, 44, 369–377. [Google Scholar] [CrossRef]
- Kim, J.S.; Seo, Y.C.; Choi, W.Y.; Kim, H.S.; Kim, B.H.; Shin, D.H.; Lee, H.Y. Enhancement of antioxidant activities and whitening effect of Acer mono Sap through nano-encapsulation processes. Korean J. Med. Corp. Sci. 2011, 19, 191–197. [Google Scholar] [CrossRef]
- Green, L.C.; Wagner, D.A.; Glogowski, J.; Skipper, P.L.; Wishnok, J.S.; Tannenbaum, S.R. Analysis of nitrate, nitrite, and [15N] nitrate in biological fluids. Anal. Biochem. 1982, 126, 131–138. [Google Scholar] [CrossRef] [PubMed]
- Deck, L.M.; Hunsaker, L.A.; Gonzales, A.M.; Orlando, R.A.; Vander, D.L. Substituted trans-stilbenes can inhibit or enhance the TPA-induced up-regulation of activator protein-1. BMC Pharmacol. 2008, 8, 19–25. [Google Scholar] [CrossRef]
- Kim, M.R.; Kang, O.H.; Kim, S.B.; Kang, H.J.; Kim, J.E.; Hwang, H.C.; Kim, I.W.; Kwon, D.Y. The study of anti-inflammatory effect of Hwanggeumjakyak-tang extract in RAW 264.7 Macrophage. Korean J. Herbol. 2013, 28, 43–50. [Google Scholar] [CrossRef]
- Zheng, H.; Yin, J.; Gao, Z.; Huang, H.; Ji, X.; Dou, C. Disruption of Chlorella vulgaris Cells for the release of biodiesel-producing lipids: A Comparison of grinding, ultrasonication, bead milling, enzymatic lysis, and microwaves. Appl. Biochem. Biotechnol. 2011, 164, 1215–1224. [Google Scholar] [CrossRef]
- Okamoto, K.; Kihira, T.; Kobashi, G.; Sasaki, S.; Yokoyama, T.; Inaba, Y.; Nagai, M. Fruit and vegetable intake and risk of amyotrophic lateral sclerosis in Japan. Neuroepidemiology 2009, 3, 251–258. [Google Scholar] [CrossRef]
- Gruskin, B. Chlorophyll—Its therapeutic place in acute and suppurative disease: Preliminary report of clinical use and rationale. Am. J. Surg. 1940, 49, 49–55. [Google Scholar] [CrossRef]
- Ju, M.K. Protorpine, and Alkaloid Isolated from Croydalis bugeana Turca, Attenuate Neuro-Inflammation in LPS-Induced BV-2 Microglia Cells. Master’s Thesis, Kyungbuk National University, Daegu, Korea, 2017; pp. 1–56. [Google Scholar]
- Akhter, M.H.; Sabir, M.; Bhide, N.K. Anti-inflammatory effect of berberine in rats injected locally with cholera toxin. Indian J. Med. Res. 1977, 65, 133–141. [Google Scholar] [PubMed]
- Yu, M.M.; Chen, Y.F.; Pan, Y.M.; Liu, Y.C.; Tu, J.; Wang, K.; Hu, F.L. Royal jelly attenuate LPS-induced inflammation in BV-2 microglia cells through modulating NF-kB and p38/JNK signaling pathways. Med. Inflamm. 2018, 2018, 7834381. [Google Scholar] [CrossRef]
- Abramson, S.B.; Attur, M.; Amin, A.R.; Clancy, R. Nitric oxide and inflammatory mediators in the perpetuation of osteoarthritis. Curr. Rheumatol. Rep. 2001, 3, 535–564. [Google Scholar] [CrossRef] [PubMed]
- Kuo, C.L.; Chi, C.W.; Liu, T.Y. The anti-inflammatory potential of berberine in vitro and in vivo. Cancer Lett. 2004, 203, 127–137. [Google Scholar] [CrossRef] [PubMed]
- Jhun, B.S.; Jin, Q.; Kin, Y.T.; Kong, S.S.; Cho, Y.; Ha, Y.H.; Baik, H.H.; Kang, I. 5-aminoinidazole-4-carboximide riboside suppresses LPS-induced TNF-alpha production through inhibition of posphophatidylinositol-3-kinase/A activation in RAW264.7 murine macrophage. Biochem. Biophys. Res. Commun. 2004, 318, 372–380. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Extraction Process | Total Chlorophyll Contents (mg/g) | Extraction Yields (%) |
|---|---|---|
| Conventional ethanol extraction (EE) * | 6.24 ± 0.92 | 8.9 ± 1.66 |
| Ultrasonic extraction (UE) ** | 17.56 ± 1.86 | 11.6 ± 2.02 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choi, W.Y.; Sim, J.-H.; Lee, J.-Y.; Kang, D.H.; Lee, H.Y. Increased Anti-Inflammatory Effects on LPS-Induced Microglia Cells by Spirulina maxima Extract from Ultrasonic Process. Appl. Sci. 2019, 9, 2144. https://doi.org/10.3390/app9102144
Choi WY, Sim J-H, Lee J-Y, Kang DH, Lee HY. Increased Anti-Inflammatory Effects on LPS-Induced Microglia Cells by Spirulina maxima Extract from Ultrasonic Process. Applied Sciences. 2019; 9(10):2144. https://doi.org/10.3390/app9102144
Chicago/Turabian StyleChoi, Woon Yong, Jae-Hun Sim, Jung-Youl Lee, Do Hyung Kang, and Hyeon Yong Lee. 2019. "Increased Anti-Inflammatory Effects on LPS-Induced Microglia Cells by Spirulina maxima Extract from Ultrasonic Process" Applied Sciences 9, no. 10: 2144. https://doi.org/10.3390/app9102144