
The Importance of Soil Elevation and Hydroperiods in Salt Marsh Vegetation Zonation: A Case Study of Ria de Aveiro

 ,
,  , ,
, ,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Field Data Collection and Laboratory and Numerical Analysis
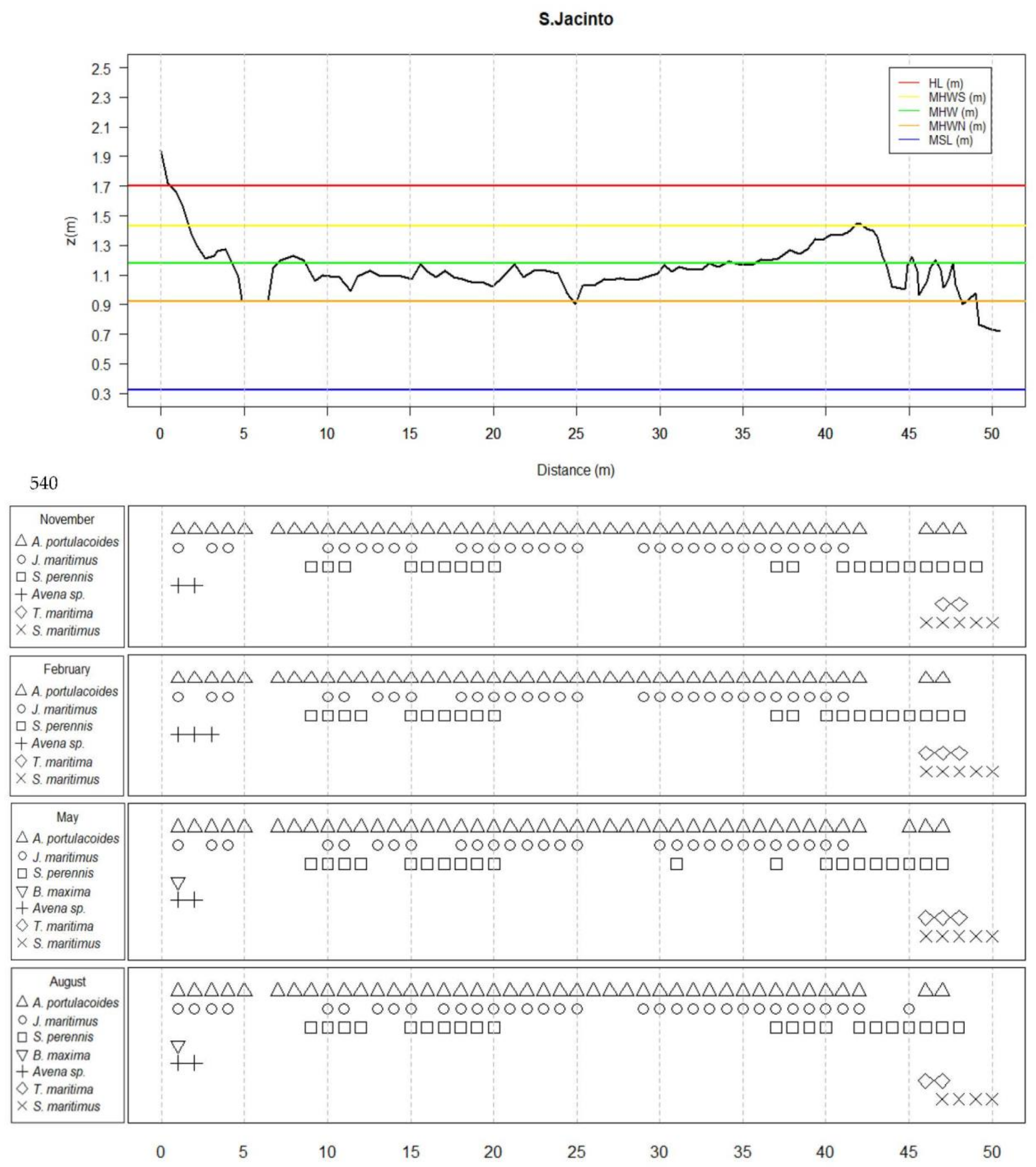
2.2.1. Characterization and Inventorying of the Transects
2.2.2. A Study of the Main Species Monospecific Stands
2.2.3. Numerical Analysis
- 1.
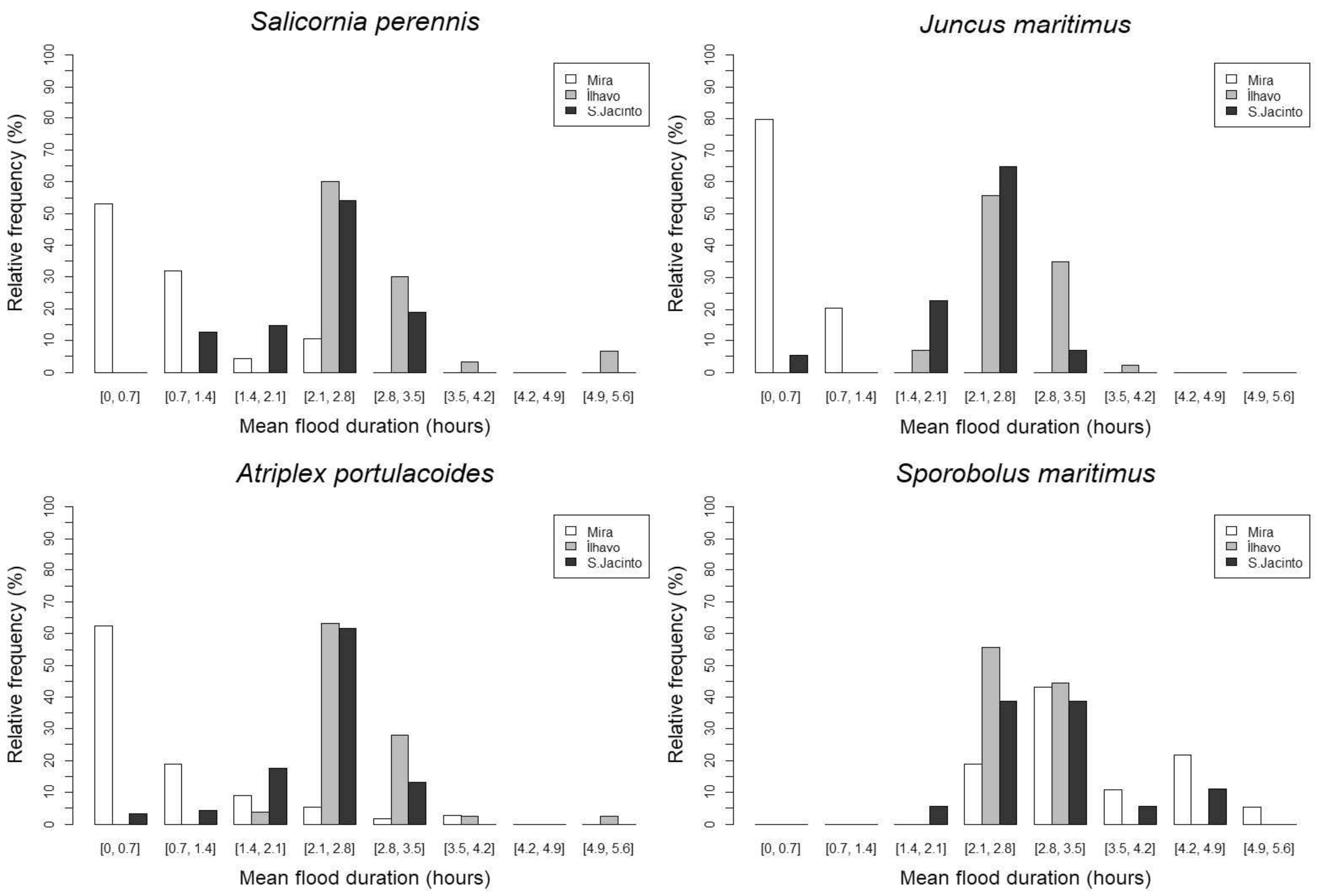
- The mean flood duration (FD), which denotes the time in hours during which the sampling point is flooded [28], was calculated:where is the duration of a single flood and is the number of floods.
- 2.
- The frequency of flooding (F), which estimates the average number of floods that occurred during a reference interval of time [28], was determined:where represents the number of floods and is the time elapsed between surveys.
- 3.
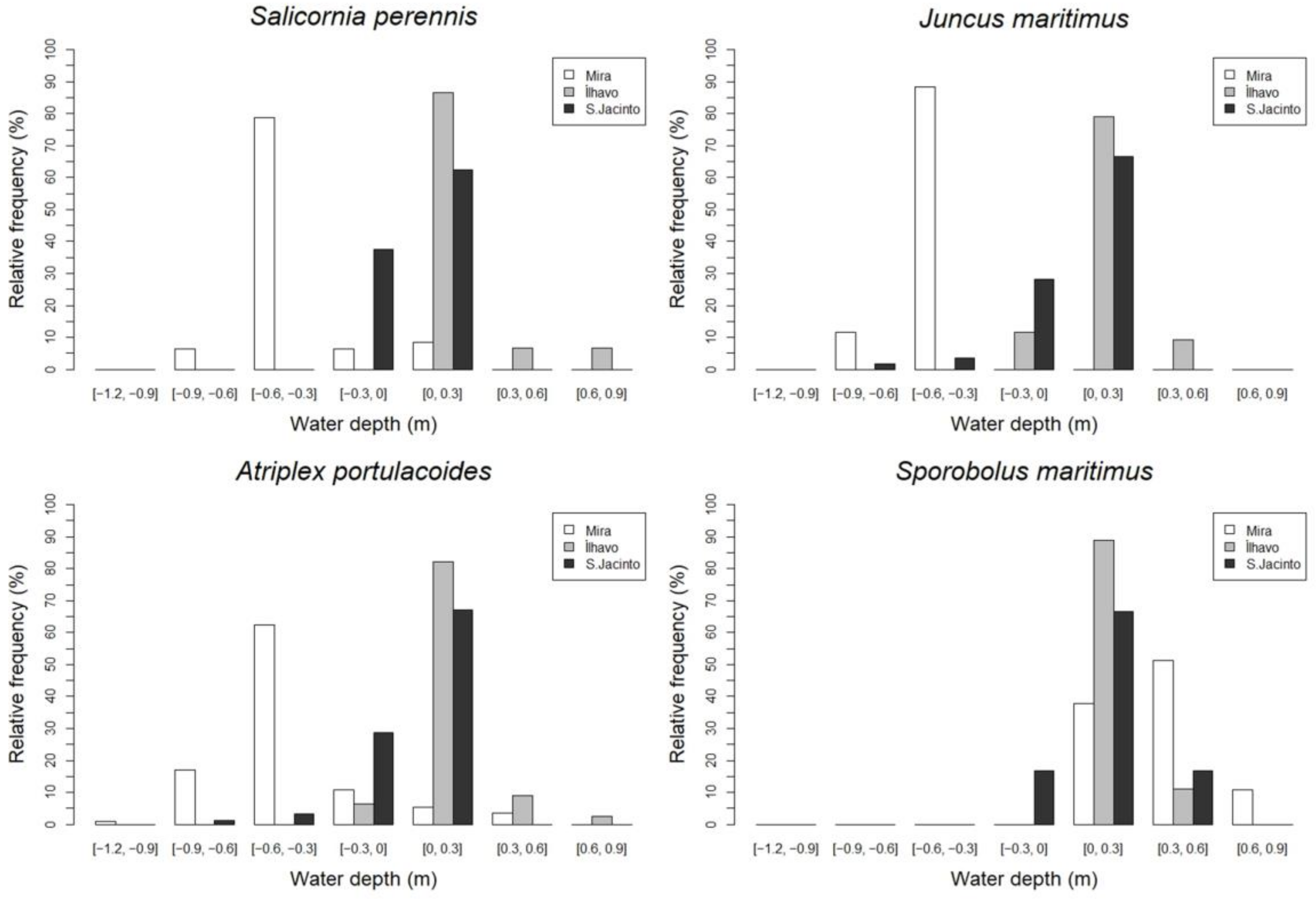
- The water depth (WD), defined as the difference between the level of mean high water (MHW) and soil elevation, z, was calculated [44]:
2.3. Statistical and Data Analysis
3. Results
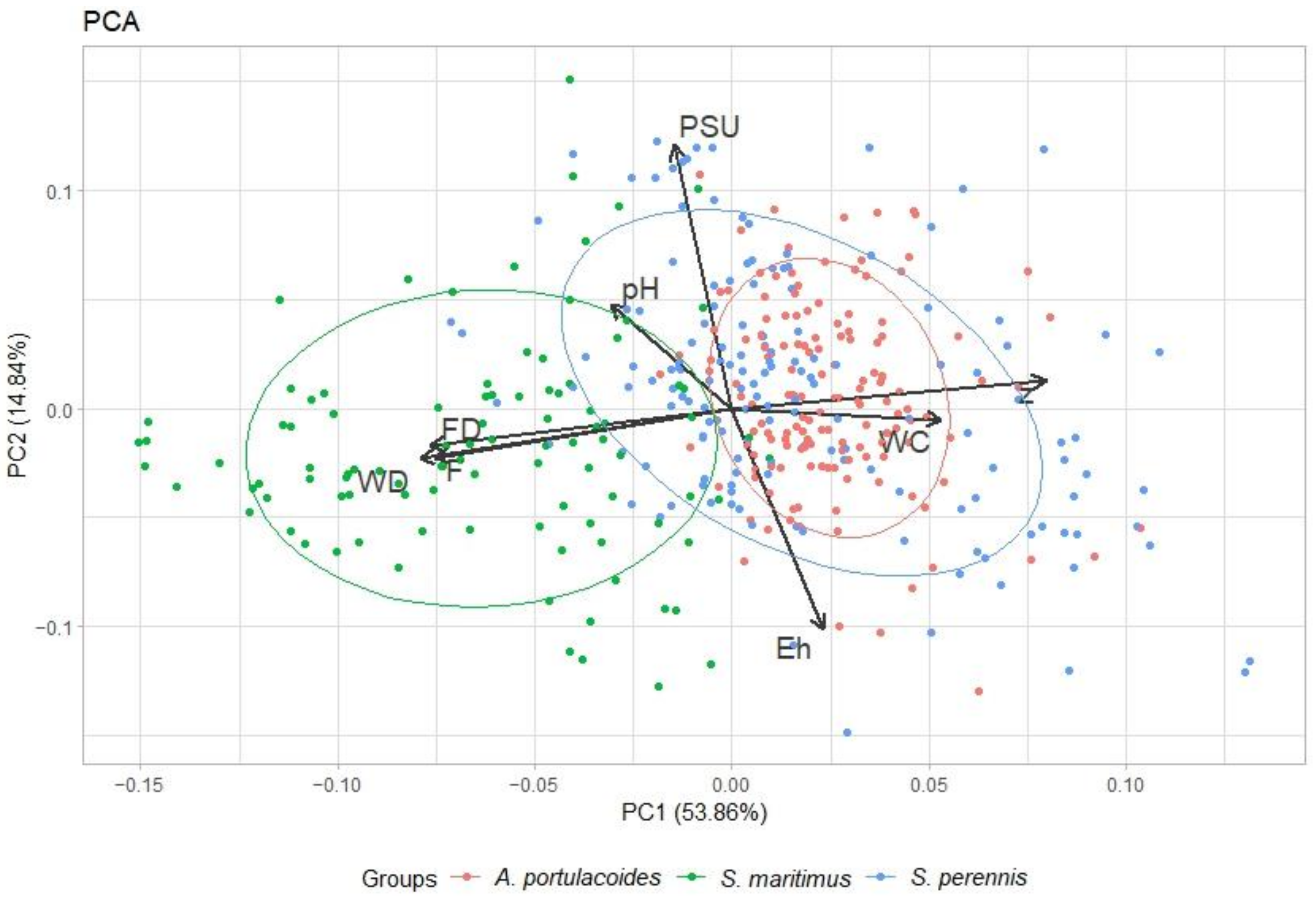
3.1. Species Distribution along the Transects
3.2. Main Species Distribution
4. Discussion
4.1. Salt Marsh Monitoring and Characterization
4.2. Biotic and Abiotic Factors versus Vegetation Monospecific Stands
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Cover-Abundance | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Mira | São Jacinto | Ílhavo | |||||||
| Months | B-B | AC | Months | B-B | AC | Months | B-B | AC | |
| Salicornia perennis | Oct–Aug | 3.33 ± 1.02 | 15.97 ± 2.24 | Oct–Aug | 3.24 ± 1.09 | 19.63 ± 1.13 | Oct–Aug | 3.86 ± 1.01 | 15.99 ± 1.33 |
| Juncus maritimus | Oct–Aug | 3.12 ± 0.97 | 25.33 ± 1.00 | Oct–Aug | 2.33 ± 0.94 | 17.42 ± 0.84 | Oct–Aug | 3.49 ± 0.92 | 15.99 ± 1.62 |
| Atriplex portulacoides | Oct–Aug | 2.86 ± 1.07 | 33.64 ± 2.71 | Oct–Aug | 4.22 ± 0.83 | 78.31 ± 0.50 | Oct–Aug | 3.48 ± 1.11 | 39.7 ± 1.50 |
| Sporobolus maritimus | Oct–Aug | 3.41 ± 0.87 | 20.75 ± 1.52 | Oct–Aug | 2.99 ± 0.68 | 2.66 ± 0.60 | Oct–Aug | + | 0.61 ± 0.16 |
| Sporobolus versicolor | Oct–Aug | 2.90 ± 1.14 | 13.95 ± 2.84 | ||||||
| Elymus repens | Oct–Aug | 3.23 ± 1.25 | 18.91 ± 4.06 | ||||||
| Carpobrotus edulis | Oct–Aug | 3.45 ± 0.70 | 2.41 ± 0.50 | ||||||
| Limbarda crithmoides | Oct–Aug | 1.96 ± 0.51 | 0.61 ± 0.23 | ||||||
| Puccinellia maritima | Nov–Aug | 2.54 ± 0.89 | 2.37 ± 1.13 | ||||||
| Spergularia marina | Nov–Aug | + | 0.12 ± 0.06 | ||||||
| Tripolium pannonicum | Oct–May | + | 0.20 ± 0.15 | ||||||
| Limonium vulgare | May–Aug | + | 0.04 ± 0.01 | ||||||
| Avena sp. | Oct–Aug | 3.56 ± 0.00 | 0.72 ± 0.35 | Oct–Aug | 3.41 ± 0.54 | 3.47 ± 0.29 | |||
| Briza maxima | May–Aug | + | 0.17 ± 0.04 | ||||||
| Triglochin maritima | Nov–Aug | 3.46 ± 0.41 | 1.02 ± 0.23 | Apr–May | 3.00 ± 0.00 | 0.36 ± 0.02 | |||
| Salicornia europaea | Oct–Nov Jul–Aug | 3.5 ± 0.00 | 0.13 ± 0.04 | ||||||
| Atriplex patula | Nov | 3.00 ± 0.00 | 0.06 ± 0.00 | ||||||
| Phragmites australis | Oct Jul–Aug | 3.00 ± 0.00 | 0.30 ± 0.19 | ||||||






References
- Adam, P. Saltmarsh Ecology; Cambridge University Press: Cambridge, UK, 1990; ISBN 0521245087. [Google Scholar]
- Adam, P. Saltmarshes in a Time of Change. Environ. Conserv. 2002, 29, 39–61. [Google Scholar] [CrossRef]
- Barbier, E.B.; Hacker, S.D.; Kennedy, C.; Koch, E.W.; Stier, A.C.; Silliman, B.R. The Value of Estuarine and Coastal Ecosystem Services. Ecol. Monogr. 2011, 81, 169–193. [Google Scholar] [CrossRef]
- Sousa, A.I.; Santos, D.B.; Da Silva, E.F.; Sousa, L.P.; Cleary, D.F.R.; Soares, A.M.V.M.; Lillebø, A.I. “Blue Carbon” and Nutrient Stocks of Salt Marshes at a Temperate Coastal Lagoon (Ria de Aveiro, Portugal). Sci. Rep. 2017, 7, srep41225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chmura, G.L.; Anisfeld, S.C.; Cahoon, D.R.; Lynch, J.C. Global Carbon Sequestration in Tidal, Saline Wetland Soils. Global Biogeochem. Cycles 2003, 17, 1111. [Google Scholar] [CrossRef]
- Owers, C.J.; Woodroffe, C.D.; Mazumder, D.; Rogers, K. Carbon Storage in Coastal Wetlands Is Related to Elevation and How It Changes over Time. Estuar. Coast. Shelf Sci. 2022, 267, 107775. [Google Scholar] [CrossRef]
- Vernberg, F.J. Salt-marsh Processes: A Review. Environ. Toxicol. Chem. 1993, 12, 2167–2195. [Google Scholar] [CrossRef]
- Sarika, M.; Zikos, A. Coastal Salt Marshes. In Handbook of Halophytes; Grigore, M.-N., Ed.; Springer: Cham, Switzerland, 2020; pp. 1–39. [Google Scholar]
- King, S.E.; Lester, J.N. The Value of Salt Marsh as a Sea Defence. Mar. Pollut. Bull. 1995, 30, 180–189. [Google Scholar] [CrossRef]
- Gedan, K.B.; Kirwan, M.L.; Wolanski, E.; Barbier, E.B.; Silliman, B.R. The Present and Future Role of Coastal Wetland Vegetation in Protecting Shorelines: Answering Recent Challenges to the Paradigm. Clim. Change 2011, 106, 7–29. [Google Scholar] [CrossRef]
- Shepard, C.C.; Crain, C.M.; Beck, M.W. The Protective Role of Coastal Marshes: A Systematic Review and Meta-Analysis. PLoS ONE 2011, 6, e27374. [Google Scholar] [CrossRef]
- Anderson, M.E.; Smith, J.M. Wave Attenuation by Flexible, Idealized Salt Marsh Vegetation. Coast. Eng. 2014, 83, 82–92. [Google Scholar] [CrossRef]
- Gedan, K.B.; Silliman, B.R.; Bertness, M.D. Centuries of Human-Driven Change in Salt Marsh Ecosystems. Ann. Rev. Mar. Sci. 2009, 1, 117–141. [Google Scholar] [CrossRef] [Green Version]
- Fitzgerald, D.M.; Hughes, Z. Marsh Processes and Their Response to Climate Change and Sea-Level Rise. Annu. Rev. Earth Planet. Sci. 2019, 47, 481–517. [Google Scholar] [CrossRef] [Green Version]
- Fagherazzi, S.; Mariotti, G.; Leonardi, N.; Canestrelli, A.; Nardin, W.; Kearney, W.S. Salt Marsh Dynamics in a Period of Accelerated Sea Level Rise. J. Geophys. Res. Earth Surf. 2020, 125, e2019JF005200. [Google Scholar] [CrossRef]
- Timmerman, A.; Haasnoot, M.; Middelkoop, H.; Bouma, T.; McEvoy, S. Ecological Consequences of Sea Level Rise and Flood Protection Strategies in Shallow Coastal Systems: A Quick-Scan Barcoding Approach. Ocean Coast. Manag. 2021, 210, 105674. [Google Scholar] [CrossRef]
- Crosby, S.C.; Sax, D.F.; Palmer, M.E.; Booth, H.S.; Deegan, L.A.; Bertness, M.D.; Leslie, H.M. Salt Marsh Persistence Is Threatened by Predicted Sea-Level Rise. Estuar. Coast. Shelf Sci. 2016, 181, 93–99. [Google Scholar] [CrossRef] [Green Version]
- Davy, A.J. Development and Structure of Salt Marshes: Community Patterns in Time and Space. In Concepts and Controversies in Tidal Marsh Ecology; Springer: Cham, Switzerland, 2005; pp. 137–156. [Google Scholar] [CrossRef]
- Rogers, K.; Woodroffe, C.D. Tidal Flats and Salt Marshes. In Coastal Environments and Global Change; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2015; pp. 227–250. ISBN 9781119117261. [Google Scholar]
- Pennings, S.C.; He, Q. Community Ecology of Salt Marshes. In Salt Marshes; FitzGerald, D., Hughes, Z., Eds.; Cambridge University Press: Cambridge, UK, 2021; pp. 82–112. [Google Scholar]
- Gao, S. Geomorphology and Sedimentology of Tidal Flats. In Coastal Wetlands: An Integrated Ecosystem Approach; Perillo, G.M.E., Wolanski, E., Cahoon, D.R., Hopkinson, C.S., Eds.; Elsevier B.V.: Amsterdam, The Netherlands, 2018; pp. 359–381. ISBN 9780444638939. [Google Scholar]
- Hughes, Z.J.; FitzGerald, D.M.; Wilson, C.A. Impacts of Climate Change and Sea Level Rise. In Salt Marshes; FitzGerald, D., Hughes, Z., Eds.; Cambridge University Press: Cambridge, UK, 2021; pp. 476–481. [Google Scholar]
- Day, J.; Burdick, D.M.; Ibáñez, C.; Mitsch, W.J.; Elsey-Quirk, T.; Rivaes, S. Restoration of Tidal Marshes. In Salt Marshes; FitzGerald, D., Hughes, Z., Eds.; Cambridge University Press: Cambridge, UK, 2021; pp. 443–475. [Google Scholar]
- Townend, I.; Fletcher, C.; Knappen, M.; Rossington, K. A Review of Salt Marsh Dynamics. Water Environ. J. 2011, 25, 477–488. [Google Scholar] [CrossRef]
- Temmerman, S.; Bouma, T.J.; Govers, G.; Wang, Z.B.; De Vries, M.B.; Herman, P.M.J. Impact of Vegetation on Flow Routing and Sedimentation Patterns: Three-Dimensional Modeling for a Tidal Marsh. J. Geophys. Res. Earth Surf. 2005, 110. [Google Scholar] [CrossRef] [Green Version]
- Costa, C.; Marangoni, J.C.; Azevedo, A.M.G. Plant Zonation in Irregularly Flooded Salt Marshes: Relative Importance of Stress Tolerance and Biological Interactions. J. Ecol. 2003, 91, 951–965. [Google Scholar] [CrossRef]
- Pennings, S.C.; Grant, M.B.; Bertness, M.D. Plant Zonation in Low-Latitude Salt Marshes: Disentangling the Roles of Flooding, Salinity and Competition. J. Ecol. 2005, 93, 159–167. [Google Scholar] [CrossRef]
- Silvestri, S.; Defina, A.; Marani, M. Tidal Regime, Salinity and Salt Marsh Plant Zonation. Estuar. Coast. Shelf Sci. 2005, 62, 119–130. [Google Scholar] [CrossRef]
- Chapman, V. Salt Marshes and Salt Deserts of the World. In Ecology of Halophytes; Reimold, R., Queen, W., Eds.; Academica Press, Inc.: London, UK, 1974; pp. 3–19. ISBN 0-12-586450-7. [Google Scholar]
- Ungar, I.A. Are Biotic Factors Significant in Influencing the Distribution of Halophytes in Saline Habitats? Bot. Rev. 1998, 64, 176–199. [Google Scholar] [CrossRef]
- Guo, H.; Pennings, S.C. Mechanisms Mediating Plant Distributions across Estuarine Landscapes in a Low-Latitude Tidal Estuary. Ecology 2012, 93, 90–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.S.; Kim, J.W. Dynamics of Zonal Halophyte Communities in Salt Marshes in the World. J. Mar. Isl. Cult. 2018, 7, 84–106. [Google Scholar] [CrossRef]
- Lopes, C.L.; Mendes, R.; Caçador, I.; Dias, J.M. Assessing Salt Marsh Loss and Degradation by Combining Long-Term LANDSAT Imagery and Numerical Modelling. Land Degrad. Dev. 2021, 32, 4534–4545. [Google Scholar] [CrossRef]
- Dias, J.M.; Picado, A. Impact of Morphologic Anthropogenic and Natural Changes in Estuarine Tidal Dynamics. J. Coast. Res. 2011, 64, 1490–1494. [Google Scholar]
- Dias, J.M.; Lopes, J.F.; Dekeyser, I. Hydrological Characterisation of Ria de Aveiro, Portugal, in Early Summer. Oceanol. Acta 1999, 22, 473–485. [Google Scholar] [CrossRef] [Green Version]
- Lopes, C.L.; Dias, J.M. Assessment of Flood Hazard during Extreme Sea Levels in a Tidally Dominated Lagoon. Nat. Hazards 2015, 77, 1345–1364. [Google Scholar] [CrossRef]
- Dias, J.M.; Pereira, F.; Picado, A.; Lopes, C.L.; Pinheiro, J.P.; Lopes, S.M.; Pinho, P.G. A Comprehensive Estuarine Hydrodynamics-Salinity Study: Impact of Morphologic Changes on Ria de Aveiro (Atlantic Coast of Portugal). J. Mar. Sci. Eng. 2021, 9, 234. [Google Scholar] [CrossRef]
- Vaz, N.; Miguel Dias, J.; Leitão, P.C. Three-Dimensional Modelling of a Tidal Channel: The Espinheiro Channel (Portugal). Cont. Shelf Res. 2009, 29, 29–41. [Google Scholar] [CrossRef]
- Picado, A.; Dias, J.M.; Fortunato, A.B. Tidal Changes in Estuarine Systems Induced by Local Geomorphologic Modifications. Cont. Shelf Res. 2010, 30, 1854–1864. [Google Scholar] [CrossRef] [Green Version]
- Lopes, C.L.; Silva, P.A.; Dias, J.M.; Rocha, A.; Picado, A.; Plecha, S.; Fortunato, A.B. Local Sea Level Change Scenarios for the End of the 21st Century and Potential Physical Impacts in the Lower Ria de Aveiro (Portugal). Cont. Shelf Res. 2011, 31, 1515–1526. [Google Scholar] [CrossRef]
- Ribeiro, A.S.; Lopes, C.L.; Sousa, M.C.; Gomez-Gesteira, M.; Dias, J.M. Flooding Conditions at Aveiro Port (Portugal) within the Framework of Projected Climate Change. J. Mar. Sci. Eng. 2021, 9, 595. [Google Scholar] [CrossRef]
- Braun-Blanquet, J. Fitosociología. Bases Para El Estudio de Las Comunidades Vegetales; H. Blume: Madrid, Spain, 1979; ISBN 9788472141742. [Google Scholar]
- Wikum, D.A.; Shanholtzer, G.F. Application of the Braun-Blanquet Cover-Abundance Scale for Vegetation Analysis in Land Development Studies. Environ. Manag. 1978, 2, 323–329. [Google Scholar] [CrossRef]
- Kefelegn, H. Mathematical Formulations for Three Components of Hydroperiod in Tidal Wetlands. Wetlands 2019, 39, 349–360. [Google Scholar] [CrossRef]
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer-Verlag: New York, NY, USA, 2016. [Google Scholar]
- Tang, Y.; Horikoshi, M.; Li, W. Ggfortify: Unified Interface to Visualize Statistical Results of Popular R Packages. R J. 2016, 8, 478–489. [Google Scholar] [CrossRef] [Green Version]
- Caçador, I.; Neto, J.M.; Duarte, B.; Barroso, D.V.; Pinto, M.; Marques, J.C. Development of an Angiosperm Quality Assessment Index (AQuA-Index) for Ecological Quality Evaluation of Portuguese Water Bodies—A Multi-Metric Approach. Ecol. Indic. 2013, 25, 141–148. [Google Scholar] [CrossRef]
- Costa, J.; Arsénio, P.; Monteiro-Henriques, T.; Neto, C.; Pereira, E.; Almeida, T.; Izco, J. Finding the Boundary between Eurosiberian and Mediterranean Salt Marshes. J. Coast. Res. 2009, 1340–1344. [Google Scholar]
- Silva, H. Aspectos Morfológicos e Ecofisiológicos de Algumas Halófitas Do Sapal Da Ria de Aveiro. Ph.D. Thesis, Universidade de Aveiro, Aveiro, Portugal, 2000. [Google Scholar]
- Bertness, M.D.; Ellison, A.M. Determinants of Pattern in a New England Salt Marsh Plant Community. Ecol. Monogr. 1987, 57, 129–147. [Google Scholar] [CrossRef]
- Hacker, S.D.; Gaines, S.D. Some Implications of Direct Positive Interactions for Community Species Diversity. Ecology 1997, 78, 1990–2003. [Google Scholar] [CrossRef]
- Bertness, M.D. Interspecific Interactions among High Marsh Perennials in a New England Salt Marsh. Ecology 1991, 72, 125–137. [Google Scholar] [CrossRef] [Green Version]
- Pennings, S.C.; Callaway, R.M. Salt Marsh Plant Zonation: The Relative Importance of Competition and Physical Factors. Ecology 1992, 73, 681–690. [Google Scholar] [CrossRef]
- Crain, C.M.; Silliman, B.R.; Bertness, S.L.; Bertness, M.D. Physical and Biotic Drivers of Plant Distribution across Estuarine Salinity Gradients. Ecology 2004, 85, 2539–2549. [Google Scholar] [CrossRef] [Green Version]
- Gallego-Tévar, B.; Grewell, B.J.; Figueroa, E.; Castillo, J.M. The Role of Exotic and Native Hybrids during Ecological Succession in Salt Marshes. J. Exp. Mar. Bio. Ecol. 2020, 523, 151282. [Google Scholar] [CrossRef]
- Bockelmann, A.C.; Neuhaus, R. Competitive Exclusion of Elymus Athericus from a High-Stress Habitat in a European Salt Marsh. J. Ecol. 1999, 87, 503–513. [Google Scholar] [CrossRef]
- Davy, A.J.; Bishop, G.F.; Mossman, H.; Redondo-Gómez, S.; Castillo, J.M.; Castellanos, E.M.; Luque, T.; Figueroa, M.E. Biological Flora of the British Isles: Sarcocornia Perennis (Miller) A.J. Scott. J. Ecol. 2006, 94, 1035–1048. [Google Scholar] [CrossRef]
- Duarte, B.; Mateos-Naranjo, E.; Goméz, S.R.; Marques, J.C.; Caçador, I. Cordgrass Invasions in Mediterranean Marshes: Past, Present and Future. In Environmental History (Netherlands); Springer Science and Business Media B.V.: Amsterdam, The Netherlands, 2018; Volume 8, pp. 171–193. [Google Scholar]
- Armstrong, W.; Wright, E.J.; Lythe, S.; Gaynard, T.J. Plant Zonation and the Effects of the Spring-Neap Tidal Cycle on Soil Aeration in a Humber Salt Marsh. J. Ecol. 1985, 73, 323. [Google Scholar] [CrossRef]
- Anastasiou, C.J.; Brooks, J.R. Effects of Soil PH, Redox Potential, and Elevation on Survival of Spartina Patens Planted at a West Central Florida Salt Marsh Restoration Site. Wetlands 2003, 23, 845–859. [Google Scholar] [CrossRef]
- Pennings, S.C.; Moore, D.J. Zonation of Shrubs in Western Atlantic Salt Marshes. Oecologia 2001, 126, 587–594. [Google Scholar] [CrossRef]
- Giblin, A.E.; Fulweiler, R.W.; Hopkinson, C.S. The Role of Marshes in Coastal Nutrient Dynamics and Loss. In Salt Marshes; Cambridge University Press: Cambridge, UK, 2021; pp. 113–154. [Google Scholar]
- Xin, P.; Kong, J.; Li, L.; Barry, D.A. Modelling of Groundwater-Vegetation Interactions in a Tidal Marsh. Adv. Water Resour. 2013, 57, 52–68. [Google Scholar] [CrossRef]
- Anderson, F.E.; Howell, B.A. Dewatering of an Unvegetated Muddy Tidal Flat during Exposure-Desiccation or Drainage? Estuaries 1984, 7, 225–232. [Google Scholar] [CrossRef]
- Anjum, N.A.; Ahmad, I.; Válega, M.; Pacheco, M.; Figueira, E.; Duarte, A.C.; Pereira, E. Impact of Seasonal Fluctuations on the Sediment-Mercury, Its Accumulation and Partitioning in Halimione Portulacoides and Juncus Maritimus Collected from Ria de Aveiro Coastal Lagoon (Portugal). Water. Air. Soil Pollut. 2011, 222, 1–15. [Google Scholar] [CrossRef]
- Sorrell, B.K.; Mendelssohn, I.A.; McKee, K.L.; Woods, R.A. Ecophysiology of Wetland Plant Roots: A Modelling Comparison of Aeration in Relation to Species Distribution. Ann. Bot. 2000, 86, 675–685. [Google Scholar] [CrossRef] [Green Version]
- Sundby, B.; Vale, C.; Caetano, M.; Luther, G.W. Redox Chemistry in the Root Zone of a Salt Marsh Sediment in the Tagus Estuary, Portugal. Aquat. Geochem. 2003, 9, 257–271. [Google Scholar] [CrossRef]
- Howes, B.L.; Howarth, R.W.; Teal, J.M.; Valiela, I. Oxidation-Reduction Potentials in a Salt Marsh: Spatial Patterns and Interactions with Primary Production1. Limnol. Oceanogr. 1981, 26, 350–360. [Google Scholar] [CrossRef]
- Dacey, J.W.H.; Howes, B.L. Water Uptake by Roots Controls Water Table Movement and Sediment Oxidation in Short Spartina Marsh. Science 1984, 224, 487–489. [Google Scholar] [CrossRef]







| Strategy | Variables | Frequency | Analysis | Description |
|---|---|---|---|---|
| Transect | Elevation (z) | Once | Field | Measured directly with a GPS-RTK on average every 0.6 m |
| Cover abundance | Every month | All species were identified and had their occupied area in an m2 registered along the transect using a 1 × 1 m quadrat | ||
| Species diversity | ||||
| Main species monospecific stands (n = 5) | Elevation (z) | Every month | Field | Measured directly with a GPS-RTK |
| Redox potential (Eh) | Measured directly with a WTW pH 330/mV meter at 4 cm depth for 10 min | |||
| pH | Laboratory | Measured with a WTW pH 330/mV meter in the porewater obtained from sediment centrifugation 1 | ||
| Salinity (PSU) | Measured with a WTW Cond 3110 SET 1 in the porewater obtained from sediment centrifugation 1 | |||
| Water content (WC) | Determined by drying 50 g of the collected sediment at 105 °C until a constant weight was obtained | |||
| Water depth (WD) | Every month | Numerical model | Determined using a hydrodynamic model developed and calibrated for the Ria de Aveiro lagoon [41] 2 | |
| Frequency of flooding (F) | ||||
| Mean flood duration (FD) |
| Cover-Abundance | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Salt Marsh | Month | S. perennis | J. maritimus | A. portulacoides | S. maritimus | ||||
| B-B | AC (%) | B-B | AC (%) | B-B | AC (%) | B-B | AC (%) | ||
| Mira | November | 3.26 | 13.89 | 3.04 | 24.43 | 2.59 | 32.49 | 3.50 | 22.43 |
| February | 3.03 | 14.80 | 2.98 | 25.53 | 2.82 | 33.33 | 3.86 | 21.09 | |
| May | 3.50 | 16.58 | 3.19 | 25.41 | 3.06 | 34.58 | 3.46 | 18.33 | |
| August | 3.30 | 19.36 | 3.15 | 26.88 | 3.11 | 37.40 | 3.18 | 19.50 | |
| São Jacinto | November | 3.37 | 20.18 | 2.26 | 16.40 | 4.00 | 78.60 | 3.33 | 2.84 |
| February | 3.11 | 20.74 | 2.52 | 19.10 | 4.02 | 78.16 | 2.75 | 2.94 | |
| May | 3.00 | 18.20 | 2.19 | 17.28 | 4.16 | 78.42 | 2.25 | 2.48 | |
| August | 3.21 | 19.50 | 2.42 | 16.70 | 4.48 | 79.10 | 2.75 | 2.72 | |
| Ílhavo | November | 3.94 | 16.94 | 3.04 | 16.00 | 3.48 | 39.30 | 1.00 | 0.61 |
| February | 4.00 | 17.09 | 3.57 | 17.36 | 3.22 | 39.30 | 1.00 | 0.60 | |
| May | 3.75 | 13.01 | 3.55 | 15.16 | 3.77 | 41.46 | 1.00 | 0.59 | |
| August | 3.86 | 16.33 | 3.86 | 16.76 | 3.60 | 39.46 | + | 0.46 | |
| PERMANOVA Table of Results | |||||||
|---|---|---|---|---|---|---|---|
| Source | df | SS | MS | Pseudo-F | P (Perm) | Unique Perms | |
| Sp | 2 | 990.13 | 495.06 | 169.34 | 0.001 * | 998 | |
| Sa | 2 | 476.63 | 238.31 | 81.515 | 0.001 * | 999 | |
| Mo | 9 | 232.15 | 25.795 | 8.8231 | 0.001 * | 997 | |
| SpxSa ** | 3 | 214.84 | 71.615 | 24.496 | 0.001 * | 999 | |
| SpxMo | 18 | 61.64 | 3.4244 | 1.1713 | 0.106 | 998 | |
| SaxMo | 18 | 177.01 | 9.8337 | 3.3636 | 0.001 * | 996 | |
| SpxSaxMo ** | 27 | 89.979 | 3.3326 | 1.1399 | 0.103 | 993 | |
| Res | 320 | 935.54 | 2.9236 | ||||
| Total | 399 | 3192 | |||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gouveia, M.M.; Magni, N.N.; Lopes, C.L.; Ribeiro, A.S.; Dias, J.M.; Silva, H. The Importance of Soil Elevation and Hydroperiods in Salt Marsh Vegetation Zonation: A Case Study of Ria de Aveiro. Appl. Sci. 2023, 13, 4605. https://doi.org/10.3390/app13074605
Gouveia MM, Magni NN, Lopes CL, Ribeiro AS, Dias JM, Silva H. The Importance of Soil Elevation and Hydroperiods in Salt Marsh Vegetation Zonation: A Case Study of Ria de Aveiro. Applied Sciences. 2023; 13(7):4605. https://doi.org/10.3390/app13074605
Chicago/Turabian StyleGouveia, Marisa Marques, Natasha Nonemacher Magni, Carina Lurdes Lopes, Américo Soares Ribeiro, João Miguel Dias, and Helena Silva. 2023. "The Importance of Soil Elevation and Hydroperiods in Salt Marsh Vegetation Zonation: A Case Study of Ria de Aveiro" Applied Sciences 13, no. 7: 4605. https://doi.org/10.3390/app13074605