
Potential of Halophytes-Associated Microbes for the Phytoremediation of Metal-Polluted Saline Soils

Abstract
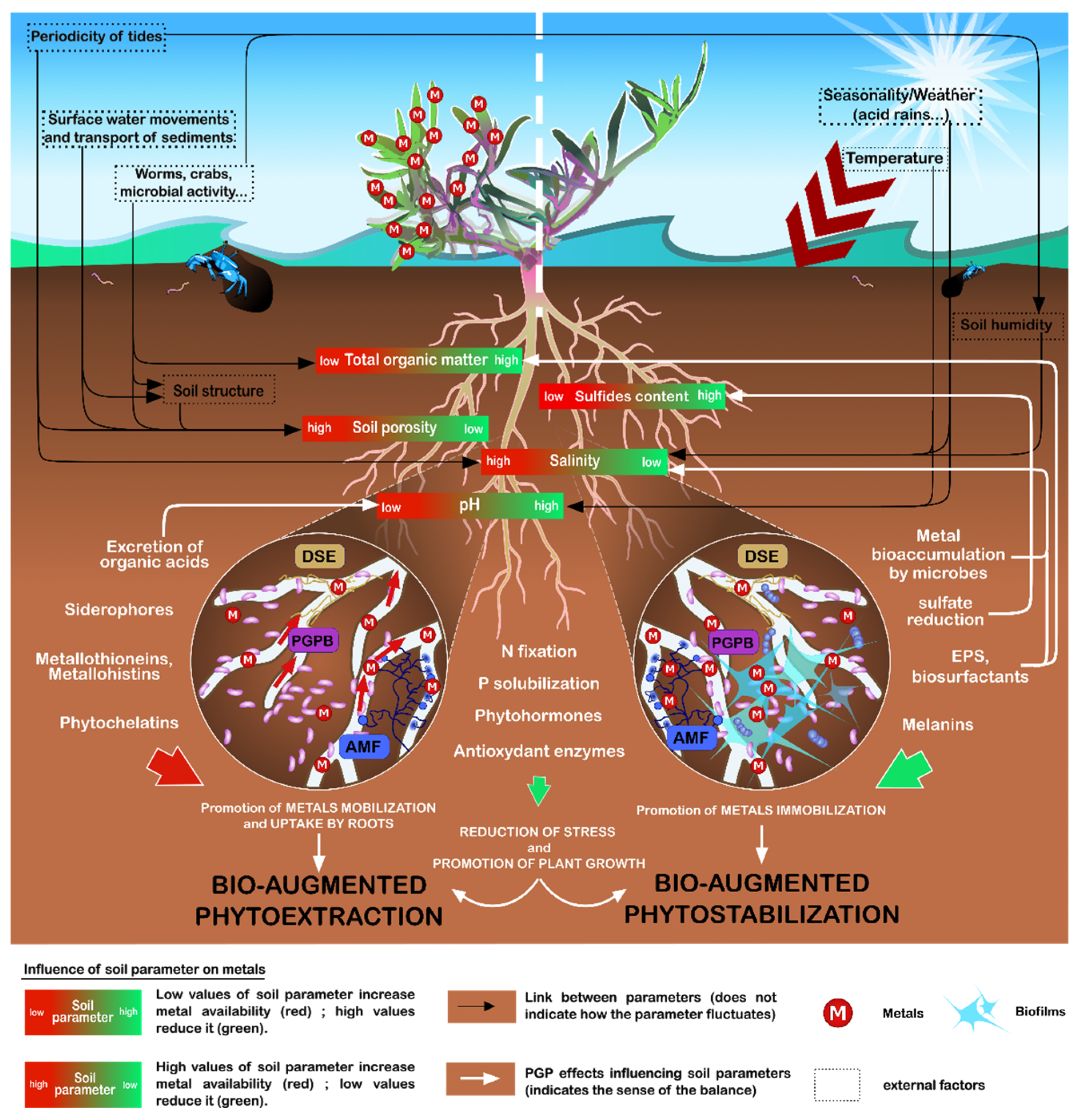
:1. Introduction
2. Potential of Halophytes and Halophytes-Associated Microorganisms for the Bioremediation of Metals in Contaminated Saline Environments
2.1. Potential of Halophytes
2.2. Potential of Halotolerant and Halophilic Microbes in the Mediation of Metal Stress in Plants
3. Selection of Microbes for the Phytoremediation of Metal Contaminated Saline Soils
3.1. Frequently Investigated PGP Properties
3.1.1. Production of Indole-3-Acetic Acid
3.1.2. Production of 1-Aminocyclopropane-1carboxylate Deaminase
3.1.3. Nutrition Improvement
3.1.4. Multiple PGP Effects in Interaction with Metal Stress
3.2. Metal Tolerance and Metal Inactivation Capacities
3.2.1. Metal Tolerance
3.2.2. Production of Exopolysaccharides
3.2.3. Arbuscular Mycorrhizal Fungi
3.2.4. Dark Septate Endophytes
3.2.5. Other Fungi
4. Designing an Appropriate Inoculum for Phytoremediation or Phytostabilization
5. Complementary Effects of Microbial Associations
6. Bioremediation Applications Using Adapted Microbes
6.1. Microbially Assisted Phytoremediation: From the Lab to the Field Applications
6.2. Field Trials
6.2.1. Potential of Applied Phytoremediation
6.2.2. Bioaugmented Phytoremediation Trials
7. Perspectives and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tripathi, P.; Singhal, A.; Jha, P.K. Metal Transport and Its Impact on Coastal Ecosystem. In Coastal Ecosystems: Environmental Importance, Current Challenges and Conservation Measures; Madhav, S., Nazneen, S., Singh, P., Eds.; Coastal Research Library; Springer International Publishing: Cham, Switzerland, 2022; pp. 239–264. ISBN 978-3-030-84255-0. [Google Scholar]
- Bai, J.; Xiao, R.; Zhao, Q.; Lu, Q.; Wang, J.; Reddy, K.R. Seasonal Dynamics of Trace Elements in Tidal Salt Marsh Soils as Affected by the Flow-Sediment Regulation Regime. PLoS ONE 2014, 9, e107738. [Google Scholar] [CrossRef]
- Wang, S.-L.; Xu, X.-R.; Sun, Y.-X.; Liu, J.-L.; Li, H.-B. Heavy Metal Pollution in Coastal Areas of South China: A Review. Mar. Pollut. Bull. 2013, 76, 7–15. [Google Scholar] [CrossRef]
- Qian, Y.; Zhang, W.; Yu, L.; Feng, H. Metal Pollution in Coastal Sediments. Curr. Pollut. Rep. 2015, 1, 203–219. [Google Scholar] [CrossRef] [Green Version]
- Poljakoff-Mayber, A.; Gale, J. Plants in Saline Environments; Ecological Studies; Poljakoff-Mayber, A., Gale, J., Eds.; Springer: Berlin/Heidelberg, Germany, 1975; Volume 15, ISBN 978-3-642-80931-6. [Google Scholar]
- Andersen, J.; Green, N.; Reker, J.; Harvey, E.T.; Murray, C. Contaminants in Europe’s Seas: Moving towards a Clean, Non-Toxic Marine Environment; European Environment Agency, Ed.; Publications Office of the European Union: Luxembourg, 2019; ISBN 978-92-9480-058-9. [Google Scholar]
- Oyetibo, G.O.; Miyauchi, K.; Huang, Y.; Chien, M.-F.; Ilori, M.O.; Amund, O.O.; Endo, G. Biotechnological Remedies for the Estuarine Environment Polluted with Heavy Metals and Persistent Organic Pollutants. Int. Biodeterior. Biodegrad. 2017, 119, 614–625. [Google Scholar] [CrossRef]
- Yousif, R.; Choudhary, M.I.; Ahmed, S.; Ahmed, Q. Review: Bioaccumulation of Heavy Metals in Fish and Other Aquatic Organisms from Karachi Coast, Pakistan. Nus. Biosci. 2021, 13, 73–84. [Google Scholar] [CrossRef]
- Briand, M.; Bustamante, P.; Bonnet, X.; Churlaud, C.; Letourneur, Y. Tracking Trace Elements into Complex Coral Reef Trophic Networks. Sci. Total Environ. 2018, 612, 1091–1104. [Google Scholar] [CrossRef] [PubMed]
- Hedfi, A.; Mahmoudi, E.; Boufahja, F.; Hamouda, B.; Aïssa, P. Effects of Increasing Levels of Nickel Contamination on Structure of Offshore Nematode Communities in Experimental Microcosms. Bull. Environ. Contam. Toxicol. 2007, 79, 345–349. [Google Scholar] [CrossRef] [PubMed]
- Agoramoorthy, G.; Chen, F.-A.; Hsu, M.J. Threat of Heavy Metal Pollution in Halophytic and Mangrove Plants of Tamil Nadu, India. Environ. Pollut. 2008, 155, 320–326. [Google Scholar] [CrossRef] [PubMed]
- Yi, J.; Lo, L.S.H.; Liu, H.; Qian, P.-Y.; Cheng, J. Study of Heavy Metals and Microbial Communities in Contaminated Sediments Along an Urban Estuary. Front. Mar. Sci. 2021, 8, 741912. [Google Scholar] [CrossRef]
- Li, Z.; Ma, Z.; van der Kuijp, T.J.; Yuan, Z.; Huang, L. A Review of Soil Heavy Metal Pollution from Mines in China: Pollution and Health Risk Assessment. Sci. Total Environ. 2014, 468–469, 843–853. [Google Scholar] [CrossRef]
- Omar, W.A.; Zaghloul, K.H.; Abdel-Khalek, A.A.; Abo-Hegab, S. Risk Assessment and Toxic Effects of Metal Pollution in Two Cultured and Wild Fish Species from Highly Degraded Aquatic Habitats. Arch. Environ. Contam. Toxicol. 2013, 65, 753–764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jain, S.; Choudhary, D.K.; Varma, A. Ecological Perspectives of Halophilic Fungi and Their Role in Bioremediation. In Soil Bioremediation; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2021; pp. 175–192. ISBN 978-1-119-54797-6. [Google Scholar]
- Li, C.; Wang, H.; Liao, X.; Xiao, R.; Liu, K.; Bai, J.; Li, B.; He, Q. Heavy Metal Pollution in Coastal Wetlands: A Systematic Review of Studies Globally over the Past Three Decades. J. Hazard. Mater. 2022, 424, 127312. [Google Scholar] [CrossRef]
- Origo, N.; Wicherek, S.; Hotyat, M. Réhabilitation des sites pollués par phytoremédiation. VertigO Rev. Électronique Sci. Environ. 2012, 12, 2. [Google Scholar] [CrossRef]
- Koptsik, G.N. Problems and Prospects Concerning the Phytoremediation of Heavy Metal Polluted Soils: A Review. Eurasian Soil Sci. 2014, 47, 923–939. [Google Scholar] [CrossRef]
- Sladkovska, T.; Wolski, K.; Bujak, H.; Radkowski, A.; Sobol, Ł. A Review of Research on the Use of Selected Grass Species in Removal of Heavy Metals. Agronomy 2022, 12, 2587. [Google Scholar] [CrossRef]
- Schwitzguébel, J. Hype or Hope: The Potential of Phytoremediation as an Emerging Green Technology. Remediat. J. 2001, 11, 63–78. [Google Scholar] [CrossRef]
- Sun, W.; Cheng, K.; Sun, K.Y.; Ma, X. Microbially Mediated Remediation of Contaminated Sediments by Heavy Metals: A Critical Review. Curr. Pollution. Rep. 2021, 7, 201–212. [Google Scholar] [CrossRef]
- Westphal, L.M.; Isebrands, J.G. Phytoremediation of Chicago’s Brownfields: Consideration of Ecological Approaches and Social Issues. In Proceedings of the Brownfields 2001, Chicago, IL, USA, 2 November 2001; Volume 9. [Google Scholar]
- Khan, M.; Shaheen, S.; Ali, S.; Yi, Z.; Cheng, L.; Samrana, S.; Khan, D.; Azam, M.; Rizwan, M.; Afzal, M.; et al. In Situ Phytoremediation of Metals. In Phytoremediation In-Situ Applications; Shmaefsky, B.R., Ed.; Springer: Berlin/Heidelberg, Germany, 2020; pp. 103–121. ISBN 978-3-030-00098-1. [Google Scholar]
- Chen, W.; Li, H. Cost-Effectiveness Analysis for Soil Heavy Metal Contamination Treatments. Water Air Soil Pollut. 2018, 229, 126. [Google Scholar] [CrossRef]
- Ma, Y.; Prasad, M.N.V.; Rajkumar, M.; Freitas, H. Plant Growth Promoting Rhizobacteria and Endophytes Accelerate Phytoremediation of Metalliferous Soils. Biotechnol. Adv. 2011, 29, 248–258. [Google Scholar] [CrossRef]
- García-Sánchez, M.; Košnář, Z.; Mercl, F.; Aranda, E.; Tlustoš, P. A Comparative Study to Evaluate Natural Attenuation, Mycoaugmentation, Phytoremediation, and Microbial-Assisted Phytoremediation Strategies for the Bioremediation of an Aged PAH-Polluted Soil. Ecotoxicol. Environ. Saf. 2018, 147, 165–174. [Google Scholar] [CrossRef]
- Mishra, A.; Mishra, S.; Agarwal, A.; Dwivedi, S. Plant-Microbe Interactions for Bioremediation and Phytoremediation of Environmental Pollutants and Agro-Ecosystem Development. In Bioremediation of Industrial Waste for Environmental Safety; Springer: Berlin/Heidelberg, Germany, 2020; Volume 2, Biological Agents and Methods for Industrial Waste Management; pp. 415–436. ISBN 9789811334252. [Google Scholar]
- Bharagava, R.; Saxena, G. Bioremediation of Industrial Waste for Environmental Safety—Volume II: Biological Agents and Methods for Industrial Waste Management; Springer: Singapore, 2019; ISBN 9789811334269. [Google Scholar]
- He, W.; Megharaj, M.; Wu, C.-Y.; Subashchandrabose, S.R.; Dai, C.-C. Endophyte-Assisted Phytoremediation: Mechanisms and Current Application Strategies for Soil Mixed Pollutants. Crit. Rev. Biotechnol. 2020, 40, 31–45. [Google Scholar] [CrossRef] [PubMed]
- Mesa-Marín, J.; Pérez-Romero, J.A.; Redondo-Gómez, S.; Pajuelo, E.; Rodríguez-Llorente, I.D.; Mateos-Naranjo, E. Impact of Plant Growth Promoting Bacteria on Salicornia Ramosissima Ecophysiology and Heavy Metal Phytoremediation Capacity in Estuarine Soils. Front. Microbiol. 2020, 11, 553018. [Google Scholar] [CrossRef] [PubMed]
- Ayyappan, D.; Sathiyaraj, G.; Ravindran, K. Phytoextraction of Heavy Metals by Sesuvium Portulacastrum, L. a Salt Marsh Halophyte from Tannery Effluent. Int. J. Phytoremediation 2015, 18, 453–459. [Google Scholar] [CrossRef]
- Caparrós, P.G.; Ozturk, M.; Gul, A.; Batool, T.S.; Pirasteh-Anosheh, H.; Unal, B.T.; Altay, V.; Toderich, K.N. Halophytes Have Potential as Heavy Metal Phytoremediators: A Comprehensive Review. Environ. Exp. Bot. 2022, 193, 104666. [Google Scholar] [CrossRef]
- Liang, L.; Liu, W.; Sun, Y.; Huo, X.; Li, S.; Zhou, Q. Phytoremediation of Heavy Metal Contaminated Saline Soils Using Halophytes: Current Progress and Future Perspectives. Environ. Rev. 2016, 25, 269–281. [Google Scholar] [CrossRef] [Green Version]
- Manousaki, E.; Kalogerakis, N. Halophytes--an Emerging Trend in Phytoremediation. Int. J. Phytoremediation 2011, 13, 959–969. [Google Scholar] [CrossRef]
- Mousavi Kouhi, S.M.; Moudi, M. Assessment of Phytoremediation Potential of Native Plant Species Naturally Growing in a Heavy Metal-Polluted Saline–Sodic Soil. Environ. Sci. Pollut. Res. 2020, 27, 10027–10038. [Google Scholar] [CrossRef]
- Naikoo, M.I.; Kafeel, U.; Naushin, F.; Khan, F.A. Halophytes in India and Their Role in Phytoremediation. In Handbook of Halophytes: From Molecules to Ecosystems towards Biosaline Agriculture; Grigore, M.-N., Ed.; Springer International Publishing: Cham, Switzerland, 2020; pp. 1–21. ISBN 978-3-030-17854-3. [Google Scholar]
- Sruthi, P.; Shackira, A.M.; Puthur, J.T. Heavy Metal Detoxification Mechanisms in Halophytes: An Overview. Wetl. Ecol. Manag. 2017, 25, 129–148. [Google Scholar] [CrossRef]
- Flowers, T.J.; Colmer, T.D. Plant Salt Tolerance: Adaptations in Halophytes. Ann. Bot. 2015, 115, 327–331. [Google Scholar] [CrossRef] [Green Version]
- Hasanuzzaman, M.; Nahar, K.; Alam, M.M.; Bhowmik, P.C.; Hossain, M.A.; Rahman, M.M.; Prasad, M.N.V.; Ozturk, M.; Fujita, M. Potential Use of Halophytes to Remediate Saline Soils. Biomed. Res. Int. 2014, 2014, 589341. [Google Scholar] [CrossRef] [PubMed]
- Hasanuzzaman, M.; Shabala, S.; Fujita, M. Halophytes and Climate Change: Adaptive Mechanisms and Potential Uses; CABI: Boston, MA, USA, 2019; ISBN 978-1-78639-433-0. [Google Scholar]
- Etesami, H.; Beattie, G.A. Mining Halophytes for Plant Growth-Promoting Halotolerant Bacteria to Enhance the Salinity Tolerance of Non-Halophytic Crops. Front. Microbiol. 2018, 9, 148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Etesami, H.; Glick, B.R. Halotolerant Plant Growth–Promoting Bacteria: Prospects for Alleviating Salinity Stress in Plants. Environ. Exp. Bot. 2020, 178, 104124. [Google Scholar] [CrossRef]
- Kumar, A.; Singh, S.; Gaurav, A.K.; Srivastava, S.; Verma, J.P. Plant Growth-Promoting Bacteria: Biological Tools for the Mitigation of Salinity Stress in Plants. Front. Microbiol. 2020, 11, 1216. [Google Scholar] [CrossRef] [PubMed]
- Kumar Arora, N.; Fatima, T.; Mishra, J.; Mishra, I.; Verma, S.; Verma, R.; Verma, M.; Bhattacharya, A.; Verma, P.; Mishra, P.; et al. Halo-Tolerant Plant Growth Promoting Rhizobacteria for Improving Productivity and Remediation of Saline Soils. J. Adv. Res. 2020, 26, 69–82. [Google Scholar] [CrossRef]
- Navarro-Torre, S.; Mateos-Naranjo, E.; Caviedes, M.A.; Pajuelo, E.; Rodríguez-Llorente, I.D. Isolation of Plant-Growth-Promoting and Metal-Resistant Cultivable Bacteria from Arthrocnemum macrostachyum in the Odiel Marshes with Potential Use in Phytoremediation. Mar. Pollut. Bull. 2016, 110, 133–142. [Google Scholar] [CrossRef] [PubMed]
- Paredes-Páliz, K.I.; Caviedes, M.A.; Doukkali, B.; Mateos-Naranjo, E.; Rodríguez-Llorente, I.D.; Pajuelo, E. Screening Beneficial Rhizobacteria from Spartina maritima for Phytoremediation of Metal Polluted Salt Marshes: Comparison of Gram-Positive and Gram-Negative Strains. Environ. Sci. Pollut. Res. Int. 2016, 23, 19825–19837. [Google Scholar] [CrossRef]
- Desale, P.; Patel, B.; Singh, S.; Malhotra, A.; Nawani, N. Plant Growth Promoting Properties of Halobacillus sp. and Halomonas sp. in Presence of Salinity and Heavy Metals. J. Basic Microbiol. 2014, 54, 781–791. [Google Scholar] [CrossRef] [PubMed]
- Bano, A.; Hussain, J.; Akbar, A.; Mehmood, K.; Anwar, M.; Hasni, M.S.; Ullah, S.; Sajid, S.; Ali, I. Biosorption of Heavy Metals by Obligate Halophilic Fungi. Chemosphere 2018, 199, 218–222. [Google Scholar] [CrossRef]
- Margesin, R.; Schinner, F. Potential of Halotolerant and Halophilic Microorganisms for Biotechnology. Extremophiles 2001, 5, 73–83. [Google Scholar] [CrossRef]
- Zhuang, X.; Han, Z.; Bai, Z.; Zhuang, G.; Shim, H. Progress in Decontamination by Halophilic Microorganisms in Saline Wastewater and Soil. Environ. Pollut. 2010, 158, 1119–1126. [Google Scholar] [CrossRef] [PubMed]
- Voica, D.M.; Bartha, L.; Banciu, H.L.; Oren, A. Heavy Metal Resistance in Halophilic Bacteria and Archaea. FEMS Microbiol. Lett. 2016, 363, fnw146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sessitsch, A.; Kuffner, M.; Kidd, P.; Vangronsveld, J.; Wenzel, W.W.; Fallmann, K.; Puschenreiter, M. The Role of Plant-Associated Bacteria in the Mobilization and Phytoextraction of Trace Elements in Contaminated Soils. Soil Biol. Biochem. 2013, 60, 182–194. [Google Scholar] [CrossRef] [Green Version]
- Seneviratne, M.; Seneviratne, G.; Madawala, H.; Vithanage, M. Role of Rhizospheric Microbes in Heavy Metal Uptake by Plants. In Agro-Environmental Sustainability: Volume 2: Managing Environmental Pollution; Singh, J.S., Seneviratne, G., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 147–163. ISBN 978-3-319-49727-3. [Google Scholar]
- Sarwar, N.; Imran, M.; Shaheen, M.R.; Ishaque, W.; Kamran, M.A.; Matloob, A.; Rehim, A.; Hussain, S. Phytoremediation Strategies for Soils Contaminated with Heavy Metals: Modifications and Future Perspectives. Chemosphere 2017, 171, 710–721. [Google Scholar] [CrossRef]
- Mishra, J.; Singh, R.; Arora, N.K. Alleviation of Heavy Metal Stress in Plants and Remediation of Soil by Rhizosphere Microorganisms. Front. Microbiol. 2017, 8, 1706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, Y.; Rajkumar, M.; Zhang, C.; Freitas, H. Beneficial Role of Bacterial Endophytes in Heavy Metal Phytoremediation. J. Environ. Manag. 2016, 174, 14–25. [Google Scholar] [CrossRef]
- Lal, S.; Ratna, S.; Said, O.B.; Kumar, R. Biosurfactant and Exopolysaccharide-Assisted Rhizobacterial Technique for the Remediation of Heavy Metal Contaminated Soil: An Advancement in Metal Phytoremediation Technology. Environ. Technol. Innov. 2018, 10, 243–263. [Google Scholar] [CrossRef]
- Guerrero-Zúñiga, A.; López-López, E.; Rodríguez-Tovar, A.; Rodriguez Dorantes, A. Functional Diversity of Plant Endophytes and Their Role in Assisted Phytoremediation. In Bioremediation of Industrial Waste for Environmental Safety; Springer: Singapore, 2019; ISBN 9789811334252. [Google Scholar]
- Qin, Y.; Druzhinina, I.S.; Pan, X.; Yuan, Z. Microbially Mediated Plant Salt Tolerance and Microbiome-Based Solutions for Saline Agriculture. Biotechnol. Adv. 2016, 34, 1245–1259. [Google Scholar] [CrossRef]
- Orhan, F.; Demirci, A. Salt Stress Mitigating Potential of Halotolerant/Halophilic Plant Growth Promoting. Geomicrobiol. J. 2020, 37, 663–669. [Google Scholar] [CrossRef]
- Furtado, B.U.; Gołębiewski, M.; Skorupa, M.; Hulisz, P.; Hrynkiewicz, K. Bacterial and Fungal Endophytic Microbiomes of Salicornia europaea. Appl. Environ. Microbiol. 2019, 85, e00305-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishtiyaq, S.; Kumar, H.; Varun, M.; Ogunkunle, C.O.; Paul, M.S. Chapter 20—Role of Secondary Metabolites in Salt and Heavy Metal Stress Mitigation by Halophytic Plants: An Overview. In Handbook of Bioremediation; Hasanuzzaman, M., Prasad, M.N.V., Eds.; Academic Press: Cambridge, MA, USA, 2021; pp. 307–327. ISBN 978-0-12-819382-2. [Google Scholar]
- Malik, Z.H.; Ravindran, K.C. Bioaccumulation of Salts and Heavy Metals By Suaeda monoica A Salt Marsh Halophyte from Paper Mill Effluent Contaminated Soil. Int. J. Sci. Technol. Res. 2020, 9, 7248–7254. [Google Scholar]
- Lutts, S.; Lefèvre, I. How Can We Take Advantage of Halophyte Properties to Cope with Heavy Metal Toxicity in Salt-Affected Areas? Ann. Bot. 2015, 115, 509–528. [Google Scholar] [CrossRef]
- Lim, S.D.; Mayer, J.A.; Yim, W.C.; Cushman, J.C. Plant Tissue Succulence Engineering Improves Water-Use Efficiency, Water-Deficit Stress Attenuation and Salinity Tolerance in Arabidopsis. Plant J. 2020, 103, 1049–1072. [Google Scholar] [CrossRef]
- Hanana, M.; Hamrouni, L.; Cagnac, O.; Blumwald, E. Mécanismes et Stratégies Cellulaires de Tolérance à La Salinité (NaCl) Chez Les Plantes. Environ. Rev. 2011, 19, 121–141. [Google Scholar] [CrossRef]
- Alam, M.R.; Islam, R.; Anh Tran, T.K.; Van, D.L.; Rahman, M.M.; Griffin, A.S.; Yu, R.M.K.; MacFarlane, G.R. Global Patterns of Accumulation and Partitioning of Metals in Halophytic Saltmarsh Taxa: A Phylogenetic Comparative Approach. J. Hazard. Mater. 2021, 414, 125515. [Google Scholar] [CrossRef] [PubMed]
- Aziz, I.; Mujeeb, A. Halophytes for Phytoremediation of Hazardous Metal(Loid)s: A Terse Review on Metal Tolerance, Bio-Indication and Hyperaccumulation. J. Hazard. Mater. 2022, 424, 127309. [Google Scholar] [CrossRef]
- Mujeeb, A.; Aziz, I.; Ahmed, M.Z.; Alvi, S.K.; Shafiq, S. Comparative Assessment of Heavy Metal Accumulation and Bio-Indication in Coastal Dune Halophytes. Ecotoxicol. Environ. Saf. 2020, 195, 110486. [Google Scholar] [CrossRef] [PubMed]
- Zurayk, R.A.; Khoury, N.F.; Talhouk, S.N.; Baalbaki, R.Z. Salinity-Heavy Metal Interactions in Four Salt-Tolerant Plant Species. J. Plant Nutr. 2001, 24, 1773–1786. [Google Scholar] [CrossRef]
- Wali, M.; Fourati, E.; Hmaeid, N.; Ghabriche, R.; Poschenrieder, C.; Abdelly, C.; Ghnaya, T. NaCl Alleviates Cd Toxicity by Changing Its Chemical Forms of Accumulation in the Halophyte Sesuvium portulacastrum. Environ. Sci. Pollut. Res. 2015, 22, 10769–10777. [Google Scholar] [CrossRef]
- Fourati, E.; Vogel-Mikuš, K.; Wali, M.; Kavčič, A.; Gomilšek, J.P.; Kodre, A.; Kelemen, M.; Vavpetič, P.; Pelicon, P.; Abdelly, C.; et al. Nickel Tolerance and Toxicity Mechanisms in the Halophyte Sesuvium portulacastrum, L. as Revealed by Ni Localization and Ligand Environment Studies. Environ. Sci. Pollut. Res. 2020, 27, 23402–23410. [Google Scholar] [CrossRef]
- Nouairi, I.; Ghnaya, T.; Ben Youssef, N.; Zarrouk, M.; Habib Ghorbel, M. Changes in Content and Fatty Acid Profiles of Total Lipids of Two Halophytes: Sesuvium portulacastrum and Mesembryanthemum crystallinum under Cadmium Stress. J. Plant Physiol. 2006, 163, 1198–1202. [Google Scholar] [CrossRef] [PubMed]
- Rabier, J.; Laffont-Schwob, I.; Anca, P.; Ellili, A.; D’Enjoy, G.; Salducci, M.-D.; Prudent, P.; Lotmani, B.; Tonetto, A.; Masotti, V. Heavy Metal and Arsenic Resistance of the Halophyte Atriplex halimus, L. Along a Gradient of Contamination in a French Mediterranean Spray Zone. Water Air Soil Pollut. 2014, 225, 1993. [Google Scholar] [CrossRef] [Green Version]
- Kachout, S.S.; Mansoura, A.B.; Mechergui, R.; Leclerc, J.C.; Rejeb, M.N.; Ouerghi, Z. Accumulation of Cu, Pb, Ni and Zn in the Halophyte Plant Atriplex Grown on Polluted Soil. J. Sci. Food Agric. 2012, 92, 336–342. [Google Scholar] [CrossRef] [PubMed]
- Kachout, S.; Mansoura, B.; Bouraoui, K. Effects of Metal Toxicity on Growth and Pigment Contents of Annual Halophyte (A. hortensis and A. rosea). Int. J. Environ. Res. 2015, 9, 613–620. [Google Scholar]
- Samiei, L.; Davoudi Pahnehkolayi, M.; Karimian, Z.; Nabati, J. Morpho-Physiological Responses of Halophyte Climacoptera crassa to Salinity and Heavy Metal Stresses in In Vitro Condition. S. Afr. J. Bot. 2020, 131, 468–474. [Google Scholar] [CrossRef]
- Mujeeb, A.; Aziz, I.; Ahmed, M.Z.; Shafiq, S.; Fatima, S.; Alvi, S.K. Spatial and Seasonal Metal Variation, Bioaccumulation and Biomonitoring Potential of Halophytes from Littoral Zones of the Karachi Coast. Sci. Total Environ. 2021, 781, 146715. [Google Scholar] [CrossRef]
- De la Rosa, G.; Peralta-Videa, J.R.; Montes, M.; Parsons, J.G.; Cano-Aguilera, I.; Gardea-Torresdey, J.L. Cadmium Uptake and Translocation in Tumbleweed (Salsola Kali), a Potential Cd-Hyperaccumulator Desert Plant Species: ICP/OES and XAS Studies. Chemosphere 2004, 55, 1159–1168. [Google Scholar] [CrossRef]
- Bareen, F.; Tahira, S.A. Metal Accumulation Potential of Wild Plants in Tannery Effluent Contaminated Soil of Kasur, Pakistan: Field Trials for Toxic Metal Cleanup Using Suaeda fruticosa. J. Hazard. Mater. 2011, 186, 443–450. [Google Scholar] [CrossRef]
- Brenner, K.; You, L.; Arnold, F.H. Engineering Microbial Consortia: A New Frontier in Synthetic Biology. Trends. Biotechnol. 2008, 26, 483–489. [Google Scholar] [CrossRef]
- Bromham, L.; Hua, X.; Cardillo, M. Macroevolutionary and Macroecological Approaches to Understanding the Evolution of Stress Tolerance in Plants. Plant Cell Environ. 2020, 43, 2832–2846. [Google Scholar] [CrossRef]
- Mohapatra, R.K.; Parhi, P.K.; Patra, J.K.; Panda, C.R.; Thatoi, H.N. Biodetoxification of Toxic Heavy Metals by Marine Metal Resistant Bacteria- A Novel Approach for Bioremediation of the Polluted Saline Environment. In Microbial Biotechnology: Volume 1. Applications in Agriculture and Environment; Patra, J.K., Vishnuprasad, C.N., Das, G., Eds.; Springer: Singapore, 2017; pp. 343–376. ISBN 978-981-10-6847-8. [Google Scholar]
- Yuan, Z.; Druzhinina, I.S.; Labbé, J.; Redman, R.; Qin, Y.; Rodriguez, R.; Zhang, C.; Tuskan, G.A.; Lin, F. Specialized Microbiome of a Halophyte and Its Role in Helping Non-Host Plants to Withstand Salinity. Sci. Rep. 2016, 6, 32467. [Google Scholar] [CrossRef]
- Paul, S.; Bag, S.K.; Das, S.; Harvill, E.T.; Dutta, C. Molecular Signature of Hypersaline Adaptation: Insights from Genome and Proteome Composition of Halophilic Prokaryotes. Genome Biol. 2008, 9, R70. [Google Scholar] [CrossRef] [Green Version]
- Russell, N.J. Adaptive Modifications in Membranes of Halotolerant and Halophilic Microorganisms. J. Bioenerg. Biomembr. 1989, 21, 93–113. [Google Scholar] [CrossRef] [PubMed]
- Vauclare, P.; Madern, D.; Girard, E.; Gabel, F.; Zaccai, G.; Franzetti, B. New Insights into Microbial Adaptation to Extreme Saline Environments. BIO Web Conf. 2014, 2, 02001. [Google Scholar] [CrossRef] [Green Version]
- Banik, A.; Pandya, P.; Patel, B.; Rathod, C.; Dangar, M. Characterization of Halotolerant, Pigmented, Plant Growth Promoting Bacteria of Groundnut Rhizosphere and Its in-Vitro Evaluation of Plant-Microbe Protocooperation to Withstand Salinity and Metal Stress. Sci. Total Environ. 2018, 630, 231–242. [Google Scholar] [CrossRef]
- Sowmya, M.; Rejula, M.P.; Rejith, P.G.; Mohan, M.; Karuppiah, M.; Hatha, A.A.M. Heavy Metal Tolerant Halophilic Bacteria from Vembanad Lake as Possible Source for Bioremediation of Lead and Cadmium. J. Environ. Biol. 2014, 35, 655–660. [Google Scholar] [PubMed]
- Ibrahim, I.M.; Konnova, S.A.; Sigida, E.N.; Lyubun, E.V.; Muratova, A.Y.; Fedonenko, Y.P.; Elbanna, K. Bioremediation Potential of a Halophilic Halobacillus sp. Strain, EG1HP4QL: Exopolysaccharide Production, Crude Oil Degradation, and Heavy Metal Tolerance. Extremophiles 2020, 24, 157–166. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Razik, M.A.; Azmy, A.F.; Khairalla, A.S.; AbdelGhani, S. Metal Bioremediation Potential of the Halophilic Bacterium, Halomonas sp. Strain WQL9 Isolated from Lake Qarun, Egypt. Egypt. J. Aquat. Res. 2020, 46, 19–25. [Google Scholar] [CrossRef]
- Mallick, I.; Bhattacharyya, C.; Mukherji, S.; Dey, D.; Sarkar, S.C.; Mukhopadhyay, U.K.; Ghosh, A. Effective Rhizoinoculation and Biofilm Formation by Arsenic Immobilizing Halophilic Plant Growth Promoting Bacteria (PGPB) Isolated from Mangrove Rhizosphere: A Step towards Arsenic Rhizoremediation. Sci. Total Environ. 2018, 610–611, 1239–1250. [Google Scholar] [CrossRef] [PubMed]
- Amoozegar, M.A.; Hamedi, J.; Dadashipour, M.; Shariatpanahi, S. Effect of Salinity on the Tolerance to Toxic Metals and Oxyanions in Native Moderately Halophilic Spore-Forming Bacilli. World J. Microbiol. Biotechnol. 2005, 21, 1237–1243. [Google Scholar] [CrossRef]
- Marasco, R.; Mapelli, F.; Rolli, E.; Mosqueira, M.J.; Fusi, M.; Bariselli, P.; Reddy, M.; Cherif, A.; Tsiamis, G.; Borin, S.; et al. Salicornia strobilacea (Synonym of Halocnemum Strobilaceum) Grown under Different Tidal Regimes Selects Rhizosphere Bacteria Capable of Promoting Plant Growth. Front. Microbiol. 2016, 7, 1286. [Google Scholar] [CrossRef] [Green Version]
- Razzaghi Komaresofla, B.; Alikhani, H.A.; Etesami, H.; Khoshkholgh-Sima, N.A. Improved Growth and Salinity Tolerance of the Halophyte Salicornia Sp. by Co–Inoculation with Endophytic and Rhizosphere Bacteria. Appl. Soil Ecol. 2019, 138, 160–170. [Google Scholar] [CrossRef]
- Szymańska, S.; Borruso, L.; Brusetti, L.; Hulisz, P.; Furtado, B.; Hrynkiewicz, K. Bacterial Microbiome of Root-Associated Endophytes of Salicornia Europaea in Correspondence to Different Levels of Salinity. Environ. Sci. Pollut. Res. Int. 2018, 25, 25420–25431. [Google Scholar] [CrossRef] [Green Version]
- Qin, S.; Feng, W.-W.; Zhang, Y.-J.; Wang, T.-T.; Xiong, Y.-W.; Xing, K. Diversity of Bacterial Microbiota of Coastal Halophyte Limonium Sinense and Amelioration of Salinity Stress Damage by Symbiotic Plant Growth-Promoting Actinobacterium Glutamicibacter Halophytocola KLBMP 5180. Appl. Environ. Microbiol. 2018, 84, e01533-18. [Google Scholar] [CrossRef] [Green Version]
- Mateos-Naranjo, E.; Mesa, J.; Pajuelo, E.; Perez-Martin, A.; Caviedes, M.A.; Rodríguez-Llorente, I.D. Deciphering the Role of Plant Growth-Promoting Rhizobacteria in the Tolerance of the Invasive Cordgrass Spartina densiflora to Physicochemical Properties of Salt-Marsh Soils. Plant Soil 2015, 394, 45–55. [Google Scholar] [CrossRef]
- Hmaeid, N.; Wali, M.; Metoui-Ben Mahmoud, O.; Pueyo, J.J.; Ghnaya, T.; Abdelly, C. Efficient Rhizobacteria Promote Growth and Alleviate NaCl-Induced Stress in the Plant Species Sulla Carnosa. Appl. Soil Ecol. 2019, 133, 104–113. [Google Scholar] [CrossRef]
- Kong, Z.; Glick, B.R. Chapter Two—The Role of Plant Growth-Promoting Bacteria in Metal Phytoremediation. In Advances in Microbial Physiology; Poole, R.K., Ed.; Academic Press: Cambridge, MA, USA, 2017; Volume 71, pp. 97–132. ISBN 978-0-12-812385-0. [Google Scholar]
- Abts, W.; Vandenbussche, B.; De Proft, M.P.; Van de Poel, B. The Role of Auxin-Ethylene Crosstalk in Orchestrating Primary Root Elongation in Sugar Beet. Front. Plant Sci. 2017, 8, 444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muday, G.K.; Rahman, A.; Binder, B.M. Auxin and Ethylene: Collaborators or Competitors? Trends Plant Sci. 2012, 17, 181–195. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, M.; Zahir, Z.A.; Asghar, H.N.; Arshad, M. The Combined Application of Rhizobial Strains and Plant Growth Promoting Rhizobacteria Improves Growth and Productivity of Mung Bean (Vigna radiata, L.) under Salt-Stressed Conditions. Ann. Microbiol. 2012, 62, 1321–1330. [Google Scholar] [CrossRef]
- Berthelot, C.; Leyval, C.; Foulon, J.; Chalot, M.; Blaudez, D. Plant Growth Promotion, Metabolite Production and Metal Tolerance of Dark Septate Endophytes Isolated from Metal-Polluted Poplar Phytomanagement Sites. FEMS Microbiol. Ecol. 2016, 92, fiw144. [Google Scholar] [CrossRef] [Green Version]
- Khalid, A.; Tahir, S.; Arshad, M.; Zahir, Z.A.; Khalid, A.; Tahir, S.; Arshad, M.; Zahir, Z.A. Relative Efficiency of Rhizobacteria for Auxin Biosynthesis in Rhizosphere and Non-Rhizosphere Soils. Soil Res. 2004, 42, 921–926. [Google Scholar] [CrossRef]
- Spaepen, S.; Vanderleyden, J.; Remans, R. Indole-3-Acetic Acid in Microbial and Microorganism-Plant Signaling. FEMS Microbiol. Rev. 2007, 31, 425–448. [Google Scholar] [CrossRef] [Green Version]
- Gravel, V.; Antoun, H.; Tweddell, R.J. Growth Stimulation and Fruit Yield Improvement of Greenhouse Tomato Plants by Inoculation with Pseudomonas putida or Trichoderma atroviride: Possible Role of Indole Acetic Acid (IAA). Soil Biol. Biochem. 2007, 39, 1968–1977. [Google Scholar] [CrossRef]
- Haroon, U.; Liaquat, F.; Khizar, M.; Akbar, M.; Saleem, H.; Arif, S.; Ali, W.; Chaudhary, H.J.; Munis, M.F.H. Isolation of Halotolerant Bacteria from Rhizosphere of Khewra Salt Mine Halophytes and Their Application to Induce Salt Tolerance in Wheat. Geomicrobiol. J. 2021, 38, 768–775. [Google Scholar] [CrossRef]
- Radhakrishnan, R.; Shim, K.-B.; Lee, B.-W.; Hwang, C.-D.; Pae, S.-B.; Park, C.-H.; Kim, S.-U.; Lee, C.-K.; Baek, I.-Y. IAA-Producing Penicillium Sp. NICS01 Triggers Plant Growth and Suppresses Fusarium Sp.-Induced Oxidative Stress in Sesame (Sesamum indicum, L.). J. Microbiol. Biotechnol. 2013, 23, 856–863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, M.-N.; Shim, J.; You, Y.; Myung, H.; Bang, K.-S.; Cho, M.; Kamala-Kannan, S.; Oh, B.-T. Characterization of Lead Resistant Endophytic Bacillus sp. MN3-4 and Its Potential for Promoting Lead Accumulation in Metal Hyperaccumulator Alnus Firma. J. Hazard. Mater. 2012, 199–200, 314–320. [Google Scholar] [CrossRef]
- Bianco, C.; Defez, R. Medicago truncatula Improves Salt Tolerance When Nodulated by an Indole-3-Acetic Acid-Overproducing Sinorhizobium meliloti Strain. J. Exp. Bot. 2009, 60, 3097–3107. [Google Scholar] [CrossRef]
- Goswami, D.; Dhandhukia, P.; Patel, P.; Thakker, J.N. Screening of PGPR from Saline Desert of Kutch: Growth Promotion in Arachis hypogea by Bacillus licheniformis A2. Microbiol. Res. 2014, 169, 66–75. [Google Scholar] [CrossRef]
- Ikram, M.; Ali, N.; Jan, G.; Jan, F.G.; Rahman, I.U.; Iqbal, A.; Hamayun, M. IAA Producing Fungal Endophyte Penicillium roqueforti Thom., Enhances Stress Tolerance and Nutrients Uptake in Wheat Plants Grown on Heavy Metal Contaminated Soils. PLoS ONE 2018, 13, e0208150. [Google Scholar] [CrossRef] [Green Version]
- Carlos, M.-H.J.; Stefani, P.-V.Y.; Janette, A.-M.; Melani, M.-S.S.; Gabriela, P.-O. Assessing the Effects of Heavy Metals in ACC Deaminase and IAA Production on Plant Growth-Promoting Bacteria. Microbiol. Res. 2016, 188–189, 53–61. [Google Scholar] [CrossRef]
- Gupta, S.; Pandey, S. ACC Deaminase Producing Bacteria with Multifarious Plant Growth Promoting Traits Alleviates Salinity Stress in French Bean (Phaseolus Vulgaris) Plants. Front. Microbiol. 2019, 10, 1506. [Google Scholar] [CrossRef]
- Qin, S.; Zhang, Y.-J.; Yuan, B.; Xu, P.-Y.; Xing, K.; Wang, J.; Jiang, J.-H. Isolation of ACC Deaminase-Producing Habitat-Adapted Symbiotic Bacteria Associated with Halophyte Limonium sinense (Girard) Kuntze and Evaluating Their Plant Growth-Promoting Activity under Salt Stress. Plant Soil 2014, 374, 753–766. [Google Scholar] [CrossRef]
- Sagar, A.; Sayyed, R.Z.; Ramteke, P.W.; Sharma, S.; Marraiki, N.; Elgorban, A.M.; Syed, A. ACC Deaminase and Antioxidant Enzymes Producing Halophilic Enterobacter sp. PR14 Promotes the Growth of Rice and Millets under Salinity Stress. Physiol. Mol. Biol. Plants 2020, 26, 1847–1854. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.P.; Jha, P.N. Alleviation of Salinity-Induced Damage on Wheat Plant by an ACC Deaminase-Producing Halophilic Bacterium Serratia Sp. SL- 12 Isolated from a Salt Lake. Symbiosis 2016, 69, 101–111. [Google Scholar] [CrossRef]
- Viterbo, A.; Landau, U.; Kim, S.; Chernin, L.; Chet, I. Characterization of ACC Deaminase from the Biocontrol and Plant Growth-Promoting Agent Trichoderma Asperellum T203. FEMS Microbiol. Lett. 2010, 305, 42–48. [Google Scholar] [CrossRef] [PubMed]
- Xie, H.; Pasternak, J.J.; Glick, B. Isolation and Characterization of Mutants of the Plant Growth-Promoting Rhizobacterium Pseudomonas Putida GR12-2 That Overproduce Indoleacetic Acid. Curr. Microbiol. 1996, 32, 67–71. [Google Scholar] [CrossRef]
- Siddikee, M.A.; Chauhan, P.S.; Anandham, R.; Han, G.-H.; Sa, T. Isolation, Characterization, and Use for Plant Growth Promotion under Salt Stress, of ACC Deaminase-Producing Halotolerant Bacteria Derived from Coastal Soil. J. Microbiol. Biotechnol. 2010, 20, 1577–1584. [Google Scholar] [CrossRef] [Green Version]
- Perillo, G.M.E.; Wolanski, E.; Cahoon, D.R.; Brinson, M.M. Coastal Wetlands: An Integrated Ecosystem Approach, 1st ed.; Elsevier Science: Amsterdam, The Netherlands, 2009; ISBN 978-0-444-53103-2. [Google Scholar]
- Ozawa, T.; Wu, J.; Fujii, S. Effect of Inoculation with a Strain of Pseudomonas pseudoalcaligenes Isolated from the Endorhizosphere of Salicornia europea on Salt Tolerance of the Glasswort. Soil Sci. Plant Nutr. 2007, 53, 12–16. [Google Scholar] [CrossRef] [Green Version]
- Johnstone, T.C.; Nolan, E.M. Beyond Iron: Non-Classical Biological Functions of Bacterial Siderophores. J. Chem. Soc. Dalton Trans. 2015, 44, 6320–6339. [Google Scholar] [CrossRef] [Green Version]
- Mesa, J.; Mateos-Naranjo, E.; Caviedes, M.A.; Redondo-Gómez, S.; Pajuelo, E.; Rodríguez-Llorente, I.D. Scouting Contaminated Estuaries: Heavy Metal Resistant and Plant Growth Promoting Rhizobacteria in the Native Metal Rhizoaccumulator Spartina maritima. Mar. Pollut. Bull. 2015, 90, 150–159. [Google Scholar] [CrossRef]
- Mesa, J.; Rodríguez-Llorente, I.D.; Pajuelo, E.; Piedras, J.M.B.; Caviedes, M.A.; Redondo-Gómez, S.; Mateos-Naranjo, E. Moving Closer towards Restoration of Contaminated Estuaries: Bioaugmentation with Autochthonous Rhizobacteria Improves Metal Rhizoaccumulation in Native Spartina maritima. J. Hazard. Mater. 2015, 300, 263–271. [Google Scholar] [CrossRef]
- Andrades-Moreno, L.; del Castillo, I.; Parra, R.; Doukkali, B.; Redondo-Gómez, S.; Pérez-Palacios, P.; Caviedes, M.A.; Pajuelo, E.; Rodríguez-Llorente, I.D. Prospecting Metal-Resistant Plant-Growth Promoting Rhizobacteria for Rhizoremediation of Metal Contaminated Estuaries Using Spartina densiflora. Environ. Sci. Pollut. Res. 2014, 21, 3713–3721. [Google Scholar] [CrossRef] [PubMed]
- Bhaskar, P.V.; Bhosle, N.B. Bacterial Extracellular Polymeric Substance (EPS): A Carrier of Heavy Metals in the Marine Food-Chain. Environ. Int. 2006, 32, 191–198. [Google Scholar] [CrossRef]
- Stahl, A.; Pletzer, D.; Mehmood, A.; Ullrich, M.S. Marinobacter Adhaerens HP15 Harbors Two CzcCBA Efflux Pumps Involved in Zinc Detoxification. Antonie Van Leeuwenhoek 2015, 108, 649–658. [Google Scholar] [CrossRef] [PubMed]
- Srinath, T.; Verma, T.; Ramteke, P.W.; Garg, S.K. Chromium (VI) Biosorption and Bioaccumulation by Chromate Resistant Bacteria. Chemosphere 2002, 48, 427–435. [Google Scholar] [CrossRef]
- Amoozegar, M.A.; Ashengroph, M.; Malekzadeh, F.; Reza Razavi, M.; Naddaf, S.; Kabiri, M. Isolation and Initial Characterization of the Tellurite Reducing Moderately Halophilic Bacterium, Salinicoccus sp. Strain QW6. Microbiol. Res. 2008, 163, 456–465. [Google Scholar] [CrossRef] [PubMed]
- Flemming, H.-C.; Wingender, J. The Biofilm Matrix. Nat. Rev. Microbiol. 2010, 8, 623–633. [Google Scholar] [CrossRef]
- Gupta, P.; Diwan, B. Bacterial Exopolysaccharide Mediated Heavy Metal Removal: A Review on Biosynthesis, Mechanism and Remediation Strategies. Biotechnol. Rep. 2017, 13, 58–71. [Google Scholar] [CrossRef]
- Caruso, C.; Rizzo, C.; Mangano, S.; Poli, A.; Donato, P.D.; Nicolaus, B.; Finore, I.; Marco, G.D.; Michaud, L.; Giudice, A.L. Isolation, Characterization and Optimization of EPSs Produced by a Cold-Adapted Marinobacter Isolate from Antarctic Seawater. Antarct. Sci. 2019, 31, 69–79. [Google Scholar] [CrossRef]
- Mathivanan, K.; Chandirika, J.U.; Mathimani, T.; Rajaram, R.; Annadurai, G.; Yin, H. Production and Functionality of Exopolysaccharides in Bacteria Exposed to a Toxic Metal Environment. Ecotoxicol. Environ. Saf. 2021, 208, 111567. [Google Scholar] [CrossRef]
- Mukherjee, P.; Mitra, A.; Roy, M. Halomonas Rhizobacteria of Avicennia marina of Indian Sundarbans Promote Rice Growth Under Saline and Heavy Metal Stresses Through Exopolysaccharide Production. Front. Microbiol. 2019, 10, 1207. [Google Scholar] [CrossRef] [Green Version]
- Juniper, S.; Abbott, L. Vesicular-Arbuscular Mycorrhizas and Soil Salinity. Mycorrhiza 1993, 4, 45–57. [Google Scholar] [CrossRef]
- Alsadat Hashemi Nejad, R.; Kafi, M.; Jari, S.K.; Mozafari, H.; Motesharezadeh, B. Arbuscular Mycorrhizal Fungi Improve Growth, Physiological Status and Nutrients Accumulation of Ailanthus Altissima Seedlings under Cadmium Pollution and Salinity. Russ. J. Plant Physiol. 2021, 68, 266–273. [Google Scholar] [CrossRef]
- Carvalho, L.M.; Caçador, I.; Martins-Loução, M.A. Arbuscular Mycorrhizal Fungi Enhance Root Cadmium and Copper Accumulation in the Roots of the Salt Marsh Plant Aster tripolium, L. Plant Soil 2006, 285, 161–169. [Google Scholar] [CrossRef]
- Suntornvongsagul, K.; Burke, D.J.; Hamerlynck, E.P.; Hahn, D. Fate and Effects of Heavy Metals in Salt Marsh Sediments. Environ. Pollut. 2007, 149, 79–91. [Google Scholar] [CrossRef]
- Berthelot, C.; Chalot, M.; Leyval, C.; Blaudez, D. From Darkness to Light: Emergence of the Mysterious Dark Septate Endophytes in Plant Growth Promotion and Stress Alleviation. In Endophytes for a Growing World; Cambridge University Press: Cambridge, UK, 2019; pp. 143–164. ISBN 978-1-108-47176-3. [Google Scholar]
- Jumpponen, A.; Trappe, J.M. Dark Septate Endophytes: A Review of Facultative Biotrophic Root-Colonizing Fungi. New Phytol. 1998, 140, 295–310. [Google Scholar] [CrossRef]
- Ban, Y.; Tang, M.; Chen, H.; Xu, Z.; Zhang, H.; Yang, Y. The Response of Dark Septate Endophytes (DSE) to Heavy Metals in Pure Culture. PLoS ONE 2012, 7, e47968. [Google Scholar] [CrossRef]
- Zhan, F.; He, Y.; Yang, Y.; Li, Y.; Li, T.; Zhao, Z. Effects of Tricyclazole on Cadmium Tolerance and Accumulation Characteristics of a Dark Septate Endophyte (DSE), Exophiala pisciphila. Bull. Environ. Contam. Toxicol. 2016, 96, 235–241. [Google Scholar] [CrossRef]
- Priyadarshini, E.; Priyadarshini, S.S.; Cousins, B.G.; Pradhan, N. Metal-Fungus Interaction: Review on Cellular Processes Underlying Heavy Metal Detoxification and Synthesis of Metal Nanoparticles. Chemosphere 2021, 274, 129976. [Google Scholar] [CrossRef] [PubMed]
- Nosanchuk, J.D.; Stark, R.E.; Casadevall, A. Fungal Melanin: What Do We Know About Structure? Front. Microbiol. 2015, 6, 1463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, S.; Nimse, S.B.; Mathew, D.E.; Dhimmar, A.; Sahastrabudhe, H.; Gajjar, A.; Ghadge, V.A.; Kumar, P.; Shinde, P.B. Microbial Melanin: Recent Advances in Biosynthesis, Extraction, Characterization, and Applications. Biotechnol. Adv. 2021, 53, 107773. [Google Scholar] [CrossRef]
- Oh, J.-J.; Kim, J.Y.; Kim, Y.J.; Kim, S.; Kim, G.-H. Utilization of Extracellular Fungal Melanin as an Eco-Friendly Biosorbent for Treatment of Metal-Contaminated Effluents. Chemosphere 2021, 272, 129884. [Google Scholar] [CrossRef]
- Gadd, G.M.; de Rome, L. Biosorption of Copper by Fungal Melanin. Appl. Microbiol. Biotechnol. 1988, 29, 610–617. [Google Scholar] [CrossRef]
- Hidayat, I. Dark Septate Endophytes and Their Role in Enhancing Plant Resistance to Abiotic and Biotic Stresses. In Plant Growth Promoting Rhizobacteria for Sustainable Stress Management: Volume 1: Rhizobacteria in Abiotic Stress Management; Sayyed, R.Z., Arora, N.K., Reddy, M.S., Eds.; Microorganisms for Sustainability; Springer: Singapore, 2019; pp. 35–63. ISBN 9789811365362. [Google Scholar]
- Warda, S.; Fortas, Z. The Beneficial Role of Indigenous Arbuscular Mycorrhizal Fungi in Phytoremediation of Wetland Plants and Tolerance to Metal Stress. Arch. Environ. Prot. 2019, 45, 103–114. [Google Scholar] [CrossRef]
- Kandalepas, D.; Stevens, K.J.; Shaffer, G.P.; Platt, W.J. How Abundant Are Root-Colonizing Fungi in Southeastern Louisiana’s Degraded Marshes? Wetlands 2010, 30, 189–199. [Google Scholar] [CrossRef]
- Liu, X.-D.; Xie, L.; Wei, Y.; Zhou, X.; Jia, B.; Liu, J.; Zhang, S. Abiotic Stress Resistance, a Novel Moonlighting Function of Ribosomal Protein RPL44 in the Halophilic Fungus Aspergillus glaucus. Appl. Environ. Microbiol. 2014, 80, 4294–4300. [Google Scholar] [CrossRef] [Green Version]
- Kaewdoung, B.; Sutjaritvorakul, T.; Gadd, G.M.; Whalley, A.J.S.; Sihanonth, P. Heavy Metal Tolerance and Biotransformation of Toxic Metal Compounds by New Isolates of Wood-Rotting Fungi from Thailand. Geomicrobiol. J. 2016, 33, 283–288. [Google Scholar] [CrossRef]
- Paredes-Páliz, K.I.; Pajuelo, E.; Doukkali, B.; Caviedes, M.Á.; Rodríguez-Llorente, I.D.; Mateos-Naranjo, E. Bacterial Inoculants for Enhanced Seed Germination of Spartina densiflora: Implications for Restoration of Metal Polluted Areas. Mar. Pollut. Bull. 2016, 110, 396–400. [Google Scholar] [CrossRef] [PubMed]
- Paredes-Páliz, K.I.; Mateos-Naranjo, E.; Doukkali, B.; Caviedes, M.A.; Redondo-Gómez, S.; Rodríguez-Llorente, I.D.; Pajuelo, E. Modulation of Spartina densiflora Plant Growth and Metal Accumulation upon Selective Inoculation Treatments: A Comparison of Gram Negative and Gram Positive Rhizobacteria. Mar. Pollut. Bull. 2017, 125, 77–85. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Garrido, M.; Mora Navarro, J.; Murcia Navarro, F.J.; Faz Cano, Á. The Chelating Effect of Citric Acid, Oxalic Acid, Amino Acids and Pseudomonas Fluorescens Bacteria on Phytoremediation of Cu, Zn, and Cr from Soil Using Suaeda vera. Int. J. Phytoremediation 2018, 20, 1033–1042. [Google Scholar] [CrossRef]
- Zhao, G.; Cheng, J.; Sun, N.; Ma, C.; Dai, M. Two Endophytic Bacterial Strains Modulate Mn Oxidation and Accumulation in the Wetland Plant Suaeda salsa Pall. Plant Soil 2019, 438, 223–237. [Google Scholar] [CrossRef] [Green Version]
- De-Bashan, L.E.; Hernandez, J.-P.; Bashan, Y.; Maier, R.M. Bacillus pumilus ES4: Candidate Plant Growth-Promoting Bacterium to Enhance Establishment of Plants in Mine Tailings. Environ. Exp. Bot. 2010, 69, 343–352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, X.; Jia, B.; Diao, F.; Li, X.; Xu, J.; Zhang, Z.; Li, F.Y.; Guo, W. Transcriptomic Analysis Reveals the Molecular Mechanisms of Arbuscular Mycorrhizal Fungi and Nitrilotriacetic Acid on Suaeda salsa Tolerance to Combined Stress of Cadmium and Salt. Process Saf. Environ. Prot. 2022, 160, 210–220. [Google Scholar] [CrossRef]
- Durand, A.; Leglize, P.; Benizri, E. Are Endophytes Essential Partners for Plants and What Are the Prospects for Metal Phytoremediation? Plant Soil 2021, 460, 1–30. [Google Scholar] [CrossRef]
- Kaminsky, L.M.; Trexler, R.V.; Malik, R.J.; Hockett, K.L.; Bell, T.H. The Inherent Conflicts in Developing Soil Microbial Inoculants. Trends Biotechnol. 2019, 37, 140–151. [Google Scholar] [CrossRef]
- Thijs, S.; Sillen, W.; Rineau, F.; Weyens, N.; Vangronsveld, J. Towards an Enhanced Understanding of Plant–Microbiome Interactions to Improve Phytoremediation: Engineering the Metaorganism. Front. Microbiol. 2016, 7, 341. [Google Scholar] [CrossRef] [PubMed]
- Hart, M.M.; Antunes, P.M.; Chaudhary, V.B.; Abbott, L.K. Fungal Inoculants in the Field: Is the Reward Greater than the Risk? Funct. Ecol. 2018, 32, 126–135. [Google Scholar] [CrossRef] [Green Version]
- Koch, A.M.; Antunes, P.M.; Kathryn Barto, E.; Cipollini, D.; Mummey, D.L.; Klironomos, J.N. The Effects of Arbuscular Mycorrhizal (AM) Fungal and Garlic Mustard Introductions on Native AM Fungal Diversity. Biol. Invasions 2011, 13, 1627–1639. [Google Scholar] [CrossRef]
- Symanczik, S.; Courty, P.-E.; Boller, T.; Wiemken, A.; Al-Yahya’ei, M.N. Impact of Water Regimes on an Experimental Community of Four Desert Arbuscular Mycorrhizal Fungal (AMF) Species, as Affected by the Introduction of a Non-Native AMF Species. Mycorrhiza 2015, 25, 639–647. [Google Scholar] [CrossRef]
- Amir, H.; Cavaloc, Y.; Laurent, A.; Pagand, P.; Gunkel, P.; Lemestre, M.; Médevielle, V.; Pain, A.; McCoy, S. Arbuscular Mycorrhizal Fungi and Sewage Sludge Enhance Growth and Adaptation of Metrosideros laurifolia on Ultramafic Soil in New Caledonia: A Field Experiment. Sci. Total Environ. 2019, 651, 334–343. [Google Scholar] [CrossRef]
- Annamalai, S.; Sundaram, M. Electro-Bioremediation: An Advanced Remediation Technology for the Treatment and Management of Contaminated Soil. In Bioremediation of Industrial Waste for Environmental Safety; Springer: Singapore, 2019; pp. 183–214. ISBN 9789811334252. [Google Scholar]
- Wang, X.; Sun, R.; Tian, Y.; Guo, K.; Sun, H.; Liu, X.; Chu, H.; Liu, B. Long-Term Phytoremediation of Coastal Saline Soil Reveals Plant Species-Specific Patterns of Microbial Community Recruitment. mSystems 2020, 5, e00741-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bashan, Y.; de-Bashan, L.E.; Prabhu, S.R.; Hernandez, J.-P. Advances in Plant Growth-Promoting Bacterial Inoculant Technology: Formulations and Practical Perspectives (1998–2013). Plant Soil 2014, 378, 1–33. [Google Scholar] [CrossRef] [Green Version]
- Nazareth, S.; Marbaniang, T. Effect of Heavy Metals on Cultural and Morphological Growth Characteristics of Halotolerant Penicillium Morphotypes. J. Basic Microbiol. 2008, 48, 363–369. [Google Scholar] [CrossRef] [PubMed]
- Giasson, P.; Jaouich, A.; Gagné, S.; Moutoglis, P. Arbuscular Mycorrhizal Fungi Involvement in Zinc and Cadmium Speciation Change and Phytoaccumulation. Remediation J. 2005, 15, 75–81. [Google Scholar] [CrossRef]
- Brandl, H.; Bosshard, R.; Wegmann, M. Computer-Munching Microbes: Metal Leaching from Electronic Scrap by Bacteria and Fungi. Hydrometallurgy 2001, 59, 319–326. [Google Scholar] [CrossRef]
- Unz, R.F.; Shuttleworth, K.L. Microbial Mobilization and Immobilization of Heavy Metals. Curr. Opin. Biotechnol. 1996, 7, 307–310. [Google Scholar] [CrossRef] [PubMed]
- Wick, L.Y.; Remer, R.; Würz, B.; Reichenbach, J.; Braun, S.; Schäfer, F.; Harms, H. Effect of Fungal Hyphae on the Access of Bacteria to Phenanthrene in Soil. Environ. Sci. Technol. 2007, 41, 500–505. [Google Scholar] [CrossRef]
- Duponnois, R. Bacteria Helping Mycorrhiza Development. In Microbial Activity in the Rhizoshere; Mukerji, K.G., Manoharachary, C., Singh, J., Eds.; Soil Biology; Springer: Berlin/Heidelberg, Germany, 2006; pp. 297–310. ISBN 978-3-540-29420-7. [Google Scholar]
- Garg, N.; Chandel, S. Role of Arbuscular Mycorrhiza in Arresting Reactive Oxygen Species (ROS) and Strengthening Antioxidant Defense in Cajanus Cajan (L.) Millsp. Nodules under Salinity (NaCl) and Cadmium (Cd) Stress. Plant Growth Regul. 2015, 75, 521–534. [Google Scholar] [CrossRef]
- Teixeira, C.; Almeida, C.M.R.; Nunes da Silva, M.; Bordalo, A.A.; Mucha, A.P. Development of Autochthonous Microbial Consortia for Enhanced Phytoremediation of Salt-Marsh Sediments Contaminated with Cadmium. Sci. Total Environ. 2014, 493, 757–765. [Google Scholar] [CrossRef]
- Berthelot, C.; Leyval, C.; Chalot, M.; Blaudez, D. Interactions between Dark Septate Endophytes, Ectomycorrhizal Fungi and Root Pathogens in Vitro. FEMS Microbiol. Lett. 2019, 366, fnz158. [Google Scholar] [CrossRef] [Green Version]
- Beans, C. Phytoremediation Advances in the Lab but Lags in the Field. Proc. Natl. Acad. Sci. USA 2017, 114, 7475–7477. [Google Scholar] [CrossRef] [Green Version]
- Otero, X.L.; Macías, F. Spatial and Seasonal Variation in Heavy Metals in Interstitial Water of Salt Marsh Soils. Environ. Pollut. 2002, 120, 183–190. [Google Scholar] [CrossRef]
- Wan, X.-M.; Lei, M.; Chen, T. Cost–Benefit Calculation of Phytoremediation Technology for Heavy-Metal-Contaminated Soil. Sci. Total Environ. 2016, 563, 796–802. [Google Scholar] [CrossRef]
- Ghnaya, T.; Nouairi, I.; Slama, I.; Messedi, D.; Grignon, C.; Abdelly, C.; Ghorbel, M.H. Cadmium Effects on Growth and Mineral Nutrition of Two Halophytes: Sesuvium portulacastrum and Mesembryanthemum crystallinum. J. Plant Physiol. 2005, 162, 1133–1140. [Google Scholar] [CrossRef]
- Ghnaya, T.; Slama, I.; Messedi, D.; Grignon, C.; Ghorbel, M.H.; Abdelly, C. Cd-Induced Growth Reduction in the Halophyte Sesuvium portulacastrum Is Significantly Improved by NaCl. J. Plant Res. 2007, 120, 309–316. [Google Scholar] [CrossRef]
- Ghnaya, T.; Slama, I.; Messedi, D.; Grignon, C.; Ghorbel, M.H.; Abdelly, C. Effects of Cd2+ on K+, Ca2+ and N Uptake in Two Halophytes Sesuvium portulacastrum and Mesembryanthemum crystallinum: Consequences on Growth. Chemosphere 2007, 67, 72–79. [Google Scholar] [CrossRef] [PubMed]
- Guarino, C.; Sciarrillo, R. Effectiveness of in Situ Application of an Integrated Phytoremediation System (IPS) by Adding a Selected Blend of Rhizosphere Microbes to Heavily Multi-Contaminated Soils. Ecol. Eng. 2017, 99, 70–82. [Google Scholar] [CrossRef]
- Bissonnette, L.; St-Arnaud, M.; Labrecque, M. Phytoextraction of Heavy Metals by Two Salicaceae Clones in Symbiosis with Arbuscular Mycorrhizal Fungi during the Second Year of a Field Trial. Plant Soil 2010, 332, 55–67. [Google Scholar] [CrossRef]
- Yang, Y.; Liu, Y.; Li, Z.; Wang, Z.; Li, C.; Wei, H. Significance of Soil Microbe in Microbial-Assisted Phytoremediation: An Effective Way to Enhance Phytoremediation of Contaminated Soil. Int. J. Environ. Sci. Technol. 2020, 17, 2477–2484. [Google Scholar] [CrossRef]
- Frick, C.; Farrell, R.; Germida, J. Assessment of Phytoremediation as an In-Situ Technique for Cleaning Oil-Contaminated Sites; Wageningen University: Wageningen, The Netherlands, 2000. [Google Scholar]
- Wolfe, A.K.; Bjornstad, D.J. Why Would Anyone Object? An Exploration of Social Aspects of Phytoremediation Acceptability. Crit. Rev. Plant Sci. 2002, 21, 429–438. [Google Scholar] [CrossRef]
- Weir, E.; Doty, S. Social Acceptability of Phytoremediation: The Role of Risk and Values. Int. J. Phytoremediation 2016, 18, 1029–1036. [Google Scholar] [CrossRef] [PubMed]
- Lachapelle, E.; Montpetit, É. Putting Phytoremediation into Action. The Scientist 2015, 29, 18–19. [Google Scholar]
- Montpetit, É.; Lachapelle, E. New Environmental Technology Uptake and Bias toward the Status Quo: The Case of Phytoremediation. Environ. Technol. Innov. 2017, 7, 102–109. [Google Scholar] [CrossRef]
- European Union: European Commission. Communication from the Commission to the European Parliament, the Council, the European Economic and Social Commitee and the Commitee of the Regions, EU Soil Strategy for 2030 Reaping the Benefits of Healthy Soils for People, Food; EU: Brussel, Belgium, 2021. [Google Scholar]
- Yang, L.; Wang, J.; Yang, Y.; Li, S.; Wang, T.; Oleksak, P.; Chrienova, Z.; Wu, Q.; Nepovimova, E.; Zhang, X.; et al. Phytoremediation of Heavy Metal Pollution: Hotspots and Future Prospects. Ecotoxicol. Environ. Saf. 2022, 234, 113403. [Google Scholar] [CrossRef] [PubMed]
- Summersgill, M. Remediation Technology Costs in the UK & Europe; Drivers and Changes from 2001 to 2005. In 5th ICEG Environmental Geotechnics: Opportunities, Challenges and Responsibilities for Environmental Geotechnics; Thomas Telford Publishing: London, UK, 2006; pp. 310–317. ISBN 978-0-7277-3474-7. [Google Scholar]


{kind=link}
| Microbial Species Tested | Native Halophyte * | Metal(s) ** | Inoculated Plant and Its Metal Phytoremediation Potential in Parentheses | Type of Inoculum | Experimental Conditions | In Vitro Tested PGP and Metal Resistance Capacities | Effects of the Inoculation Compared to the Control | Metals’ Remediation Capacity | References |
|---|---|---|---|---|---|---|---|---|---|
| Aeromonas aquariorum, Pseudomonas composti, Bacillus sp. | Spartina densiflora | Cu | Spartina densiflora (frequently recommended for phytoremediation) | Consortium of 3 bacteria, imbibition of seeds in 1 mL of bacterial culture | Plate assay. Evaluation of seed germination rate and fungal infection | N fixation, P solubilization, siderophores, auxins, high resistance to NaCl and Cu | Enhanced seed germination rate | Not evaluated | [127] |
| Two Halobacillus sp. strains and one Halomonas sp. strain | Mangrove rhizospheric soil | Co, Cd, Ni, Hg, Ag | Sesuvium portulacastrum (frequently recommended for phytoremediation) | Consortium of the 3 bacteria. Inoculation of cuttings with a 10 mL suspension of each of strain | Pot trial with cuttings cultivated in polluted sterilized sandy soil from coastal regions | IAA, P solubilization | Higher roots length and roots dry weight | Reduced metal contents in soil especially for Cd and Ni | [47] |
| As-resistant halophilic Kocuria flava and Bacillus vietnamensis | Rhizosphere of Ceriops decandra (mangrove species) | As | Rice (none) | Single-strain inoculation and co-inoculation of seedling rhizosphere with bacterial suspensions | Pot trial with seeds germinated in Soil Rite Mix | Absorption of As in bacteria, EPS, biofilms, siderophores, IAA | Higher root length, shoot length, dry weight, wet weight, and chlorophyll content | Not evaluated | [92] |
| Pseudomonas composti, Aeromonas aquariorum, Bacillus sp. | Spartina densiflora | Cu, Fe, K, Mg, Mn, Na, Zn | Spartina densiflora (frequently recommended for phytoremediation) | Consortium of 3 bacteria. Inoculation of seedlings with an amendment of 5 mL of each bacterial suspension | Pot trial with 2 soils from non-polluted and polluted marshes each supplemented with perlite (ratio 9:1) | Not evaluated | Higher root length and diameter, higher leaf water content | Higher metal accumulation in roots and leaves | [98] |
| Bacillus methylotrophicus, Bacillus aryabhattai, B. aryabhattai, Bacillus licheniformis | Spartina maritima | As Cu Pb Zn | Medicago sativa (frequently recommended for phytoremediation of non-saline soils) | Single-strain inoculation of germinated seeds with 5 mL of bacterial suspension | Plate assay in solid Fahraeus medium with different Cu concentrations | N fixation, P solubilization, siderophores, IAA, ACCD, biofilms | Higher root length, more lateral roots, increase of root hair formation | Not evaluated | [125] |
| Vibrio neocaledonicus, Thalassospira australica, Pseudarthrobacter oxydans | Salicornia ramosissima | As, Cd, Cu, Co, Ni, Zn, Pb | Salicornia ramosissima (recommended for phytoremediation, but scarce studies on this species) | Consortium of 3 bacteria, seedlings inoculated with 50 mL of bacterial suspension | Pot trial in soils from non-polluted and polluted marshes | N fixation, P solubilization, biofilms, ACCD, IAA, siderophores | Enhanced growth rate, higher number of branches, higher photosynthetic rate and functionality of PSII, and higher electron transport rate | Higher accumulation of As, Cd, Cu, Ni, Pb, and Zn, especially in roots | [30] |
| Several strains of Halomonas sp. | Avicennia marina | As | moderately salt-tolerant rice variety-Jarava (none) | Single and 6-strain consortiums. Germination test: seeds in bacterial suspensions. Pot trial: seedlings inoculated with 5 mL of each bacterial suspension | Effect on germination: plate assay. Pot trial: soil supplemented with NaCl, urea, muriate of potash, single superphosphate and NaAsO2 | EPS, P solubilization, IAA, siderophores, NH3 production, hydrogen cyanide, N fixation | Slightly enhanced germination; higher N2 and PO43− contents in roots and shoots | Reduction of As(III) into a less toxic form [As(V)], reduction of As translocation | [136] |
| Consortia 1: Kushneria marisflavi, Micrococcus aloeverae, Bacillus vietnamensis, Halomonas zincidurans; Consortia 2: Vibrio kanaloae, Pseudoalteromonas distinct, P. prydzensis, Staphylococcus warneri | Arthrocnemum macrostachyum | As, Zn, Cu, Pb | Arthrocnemum macrostachyum (known as a promising candidate for phytostabilization of metals) | A 3-endophytic-strains consortium and a 3-rhizospheric-strains one. Seeds submerged with bacterial suspensions | Plate assay with a mixture of metals | Siderophores, N fixation, P solubilization | Acceleration of germination | Not evaluated | [45] |
| Two Pantoea agglomerans strains, Bacillus aryabhattai | Spartina maritima | As, Cu, Zn, Pb | Spartina maritima (frequently recommended for metal phytoremediation) | Single-strain inoculation and co-inoculation of sterilized seeds with 1 to 3 strains | Pot trial on seedlings, in collected sediments from non-polluted and polluted marshes | N fixation, IAA, P solubilization, siderophores | Higher germination rate, accelerated germination | Higher metal accumulation in roots only in the bacterial treatments with B. aryabhattai and with the 3 bacteria consortia | [46,156,157] |
| Pseudomonas fluorescens | Not known (from a manufacturer) | Cu, Zn, Cr | Suaeda vera (recently studied for its metal phytoextraction capacities) | One single strain. Dissolution of the inoculum in the irrigation water | Field trial on S. vera plantation. Cr−, Cu−, and Zn- contaminated soils | Not available | No effect on fresh weight | Increased metals’ accumulation especially in roots | [158] |
| Pantoea eucrina, Pseudomonas composti | Suaeda salsa | Mn | Suaeda salsa (frequently recommended for metal phytoremediation) | Single strain directly added in the hydroponic solution | Hydroponic trial in Hoagland solution supplemented with 200 μM MnCl2 | Not evaluated | Not evaluated | Enhanced Mn accumulation: Mn oxide precipitates at the root and leaves surface | [159] |
| Two Bacillus pumilus strains, Azospirillum brasilense | Atriplex lentiformis | Pb, Mn, Zn, Cu, As, Cd | Atriplex lentiformis (known for its metal-phytoextraction capacities) | Single-strain inoculation of seeds with microbead alginate inoculants added in the same planting hole | Pot trial with two substrate (acidic high-metal-content tailings and neutral low-metal-content natural tailings) supplemented with compost | N fixing and P solubilizing bacteria | Enhanced germination, root and shoot length, shoots, roots and total dry weight, root/shoot ratio, and number of leaves depending on the substrate | Not evaluated but the study aims to develop bio-assisted phytostabilization | [160] |
| Glomus mosseae | Suaeda salsa | Cd | Suaeda salsa (frequently recommended for metals phytoremediation) | Associated with biodegradable chelators nitrilotriacetic acid | Pot trial with soil from desert | Not evaluated (species largely used in the promotion of plant growth) | Reduced malondialdehyde concentrations in shoots; enhanced antioxidant defense, osmoregulation, and photosynthesis; promotion of specific bacterial communities | Enhanced Cd accumulation | [16,161] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bonaventure, P.; Guentas, L.; Burtet-Sarramegna, V.; Amir, H. Potential of Halophytes-Associated Microbes for the Phytoremediation of Metal-Polluted Saline Soils. Appl. Sci. 2023, 13, 4228. https://doi.org/10.3390/app13074228
Bonaventure P, Guentas L, Burtet-Sarramegna V, Amir H. Potential of Halophytes-Associated Microbes for the Phytoremediation of Metal-Polluted Saline Soils. Applied Sciences. 2023; 13(7):4228. https://doi.org/10.3390/app13074228
Chicago/Turabian StyleBonaventure, Pauline, Linda Guentas, Valérie Burtet-Sarramegna, and Hamid Amir. 2023. "Potential of Halophytes-Associated Microbes for the Phytoremediation of Metal-Polluted Saline Soils" Applied Sciences 13, no. 7: 4228. https://doi.org/10.3390/app13074228