
Potential of Laurel (Laurus nobilis L.) Leaf Polyphenols for Modulation of Body Composition


Abstract
:Featured Application
Abstract
1. Introduction
2. Review Method
3. Overview of Main Laurel Leaf Polyphenols’ Components
3.1. Kaempferol
3.2. Hydroxycinnamic Acids
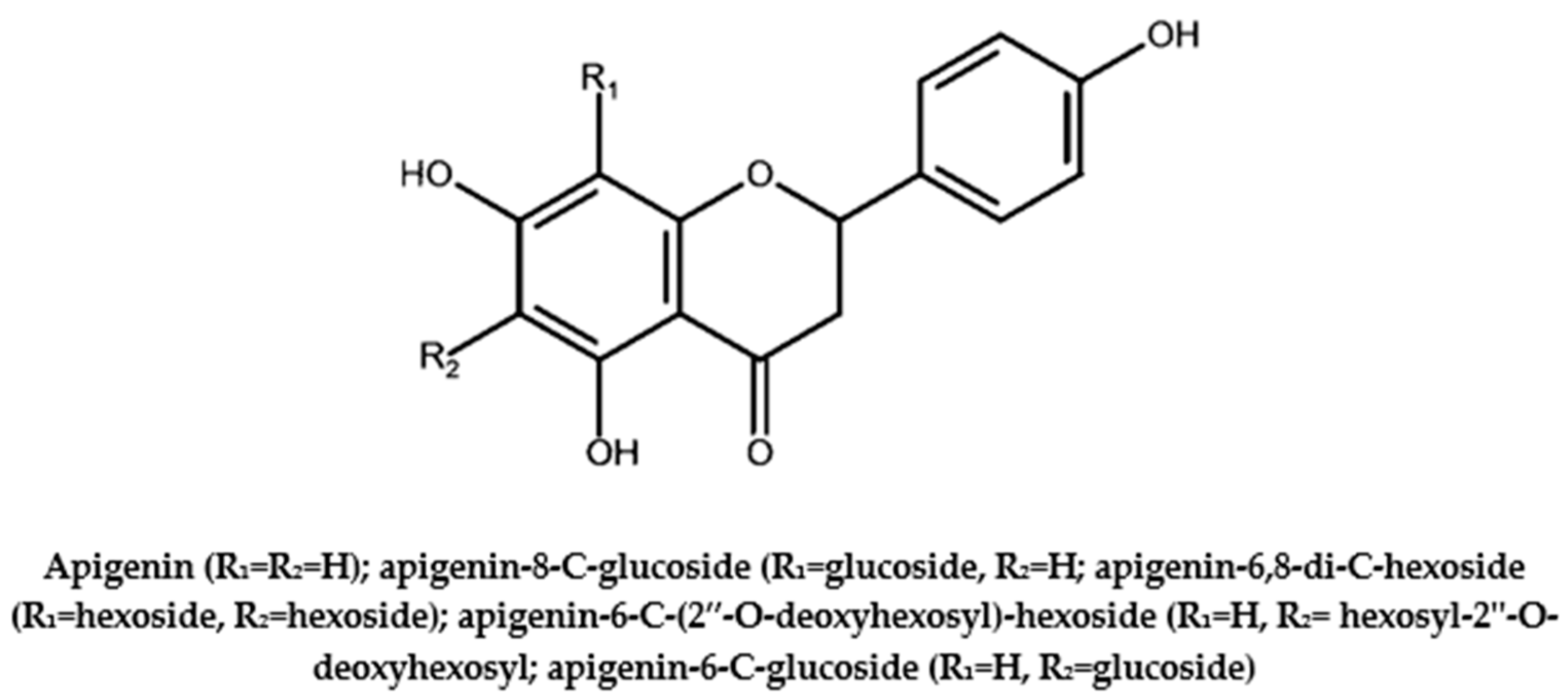
3.3. Apigenin
4. Influence of Laurel Leaf Polyphenols on Body Composition
4.1. Influence of Laurel Leaf Polyphenols on Bones
4.1.1. Kaempferol
4.1.2. Hydroxycinnamic Acids
4.1.3. Apigenin
4.2. Influence of Laurel Leaf Polyphenols on Skeletal Muscle
4.2.1. Kaempferol
4.2.2. Hydroxycinnamic Acids
4.2.3. Apigenin
4.3. Influence of Laurel Leaf Polyphenols on Adipose Tissue
4.3.1. Kaempferol
4.3.2. Hydroxycinnamic Acids
4.3.3. Apigenin
5. Conclusions and Future Perspectives
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- United Nations, Department of Economic and Social Affairs, Population Division. World Population Prospects 2019: Highlights (ST/ESA/SER.A/423); United Nations, Department of Economic and Social Affairs, Population Division: New York, NY, USA, 2019. [Google Scholar]
- Jafari Nasabian, P.; Inglis, J.E.; Reilly, W.; Kelly, O.J.; Ilich, J.Z. Aging human body: Changes in bone, muscle and body fat with consequent changes in nutrient intake. J. Endocrinol. 2017, 234, R37–R51. [Google Scholar] [CrossRef]
- Ilich, J.Z.; Gilman, J.C.; Cvijetic, S.; Boschiero, D. Chronic Stress Contributes to Osteosarcopenic Adiposity via Inflammation and Immune Modulation: The Case for More Precise Nutritional Investigation. Nutrients 2020, 12, 989. [Google Scholar] [CrossRef] [PubMed]
- Ilich, J.Z.; Kelly, O.J.; Inglis, J.E.; Panton, L.B.; Duque, G.; Ormsbee, M.J. Interrelationship among muscle, fat, and bone: Connecting the dots on cellular, hormonal, and whole body levels. Ageing Res. Rev. 2014, 15, 51–60. [Google Scholar] [CrossRef] [PubMed]
- Kelly, O.; Gilman, J.; Ilich, J. Utilizing Dietary Nutrient Ratios in Nutritional Research: Expanding the Concept of Nutrient Ratios to Macronutrients. Nutrients 2019, 11, 282. [Google Scholar] [CrossRef] [PubMed]
- Eckstrom, E.; Neukam, S.; Kalin, L.; Wright, J. Physical Activity and Healthy Aging. Clin. Geriatr. Med. 2020, 36, 671–683. [Google Scholar] [CrossRef]
- Mafra, D.; Ugochukwu, S.A.; Borges, N.A.; Cardozo, L.F.M.F.; Stenvinkel, P.; Shiels, P.G. Food for healthier aging: Power on your plate. Crit. Rev. Food Sci. Nutr. 2022, 62, 1–14. [Google Scholar] [CrossRef]
- Llaha, F.; Zamora-Ros, R. The Effects of Polyphenol Supplementation in Addition to Calorie Restricted Diets and/or Physical Activity on Body Composition Parameters: A Systematic Review of Randomized Trials. Front. Nutr. 2020, 7, 84. [Google Scholar] [CrossRef]
- Gaamouri, N.; Zouhal, H.; Hammami, M.; Hackney, A.C.; Abderrahman, A.B.; Saeidi, A.; El Hage, R.; Ounis, O. Ben Effects of polyphenol (carob) supplementation on body composition and aerobic capacity in taekwondo athletes. Physiol. Behav. 2019, 205, 22–28. [Google Scholar] [CrossRef]
- Shen, C.-L.; Cao, J.J.; Dagda, R.Y.; Chanjaplammootil, S.; Lu, C.; Chyu, M.-C.; Gao, W.; Wang, J.-S.; Yeh, J.K. Green tea polyphenols benefits body composition and improves bone quality in long-term high-fat diet–induced obese rats. Nutr. Res. 2012, 32, 448–457. [Google Scholar] [CrossRef]
- Farhat, G.; Drummond, S.; Al-Dujaili, E.A.S. Polyphenols and Their Role in Obesity Management: A Systematic Review of Randomized Clinical Trials. Phyther. Res. 2017, 31, 1005–1018. [Google Scholar] [CrossRef]
- Sharma, A.; Singh, J.; Kumar, S. Bay leaves. In Handbook of Herbs and Spices; Peter, K.V., Ed.; Woodhead Publishing Ltd.: Oxford, UK, 2012; pp. 73–85. [Google Scholar]
- Alejo-Armijo, A.; Altarejos, J.; Salido, S. Phytochemicals and biological activities of laurel tree (Laurus nobilis). Nat. Prod. Commun. 2017, 12, 743–757. [Google Scholar] [CrossRef]
- Dobroslavić, E.; Repajić, M.; Dragović-Uzelac, V.; Elez Garofulić, I. Isolation of Laurus nobilis Leaf Polyphenols: A Review on Current Techniques and Future Perspectives. Foods 2022, 11, 235. [Google Scholar] [CrossRef] [PubMed]
- Brahmi, N.; Scognamiglio, M.; Pacifico, S.; Mekhoukhe, A.; Madani, K.; Fiorentino, A.; Monaco, P. 1H NMR based metabolic profiling of eleven Algerian aromatic plants and evaluation of their antioxidant and cytotoxic properties. Food Res. Int. 2015, 76, 334–341. [Google Scholar] [CrossRef] [PubMed]
- Dias, M.I.; Barros, L.; Dueñas, M.; Alves, R.C.; Oliveira, M.B.P.P.; Santos-Buelga, C.; Ferreira, I.C.F.R. Nutritional and antioxidant contributions of Laurus nobilis L. leaves: Would be more suitable a wild or a cultivated sample? Food Chem. 2014, 156, 339–346. [Google Scholar] [CrossRef]
- Muñiz-Márquez, D.B.; Rodríguez, R.; Balagurusamy, N.; Carrillo, M.L.; Belmares, R.; Contreras, J.C.; Nevárez, G.V.; Aguilar, C.N. Phenolic content and antioxidant capacity of extracts of Laurus nobilis L., Coriandrum sativum L. and Amaranthus hybridus L. CYTA J. Food 2014, 12, 271–276. [Google Scholar] [CrossRef]
- Mazzio, E.A.; Li, N.; Bauer, D.; Mendonca, P.; Taka, E.; Darb, M.; Thomas, L.; Williams, H.; Soliman, K.F.A. Natural product HTP screening for antibacterial (E. coli 0157:H7) and anti-inflammatory agents in (LPS from E. coli O111:B4) activated macrophages and microglial cells; focus on sepsis. BMC Complement. Altern. Med. 2016, 16, 467. [Google Scholar] [CrossRef]
- Matsuda, H.; Kagerura, T.; Toguchida, I.; Ueda, H.; Morikawa, T.; Yoshikawa, M. Inhibitory effects of sesquiterpenes from bay leaf on nitric oxide production in lipopolysaccharide-activated macrophages: Structure requirement and role of heat shock protein induction. Life Sci. 2000, 66, 2151–2157. [Google Scholar] [CrossRef]
- Domazetovic, V.; Marcucci, G.; Iantomasi, T.; Brandi, M.L.; Vincenzini, M.T. Oxidative stress in bone remodeling: Role of antioxidants. Clin. Cases Miner. Bone Metab. 2017, 14, 209. [Google Scholar] [CrossRef]
- Dalle, S.; Rossmeislova, L.; Koppo, K. The Role of Inflammation in Age-Related Sarcopenia. Front. Physiol. 2017, 8, 1045. [Google Scholar] [CrossRef]
- Epsley, S.; Tadros, S.; Farid, A.; Kargilis, D.; Mehta, S.; Rajapakse, C.S. The Effect of Inflammation on Bone. Front. Physiol. 2021, 11, 511799. [Google Scholar] [CrossRef]
- Kawai, T.; Autieri, M.V.; Scalia, R. Adipose tissue inflammation and metabolic dysfunction in obesity. Am. J. Physiol. Physiol. 2021, 320, C375–C391. [Google Scholar] [CrossRef] [PubMed]
- Scicchitano, B.M.; Pelosi, L.; Sica, G.; Musarò, A. The physiopathologic role of oxidative stress in skeletal muscle. Mech. Ageing Dev. 2018, 170, 37–44. [Google Scholar] [CrossRef] [PubMed]
- Masschelin, P.M.; Cox, A.R.; Chernis, N.; Hartig, S.M. The Impact of Oxidative Stress on Adipose Tissue Energy Balance. Front. Physiol. 2020, 10, 1638. [Google Scholar] [CrossRef] [PubMed]
- Calderón-Montaño, J.M.; Burgos-Morón, E.; Pérez-Guerrero, C.; López-Lázaro, M. A Review on the Dietary Flavonoid Kaempferol BenthamScience. Mini Rev. Med. Chem. 2011, 11, 298–344. [Google Scholar] [CrossRef] [PubMed]
- Stefanova, G.; Girova, T.; Gochev, V.; Stoyanova, M.; Petkova, Z.; Stoyanova, A.; Zheljazkov, V.D. Comparative study on the chemical composition of laurel (Laurus nobilis L.) leaves from Greece and Georgia and the antibacterial activity of their essential oil. Heliyon 2020, 6, e05491. [Google Scholar] [CrossRef]
- Sova, M.; Saso, L. Natural sources, pharmacokinetics, biological activities and health benefits of hydroxycinnamic acids and their metabolites. Nutrients 2020, 12, 2190. [Google Scholar] [CrossRef]
- Dobroslavić, E.; Elez Garofulić, I.; Šeparović, J.; Zorić, Z.; Pedisić, S.; Dragović-Uzelac, V. Pressurized Liquid Extraction as a Novel Technique for the Isolation of Laurus nobilis L. Leaf Polyphenols. Molecules 2022, 27, 5099. [Google Scholar] [CrossRef]
- Sirotkin, A.V.; Kolesarova, A. Plant molecules and their influence on health and female reproduction. In Environmental Contaminants and Medicinal Plants Action on Female Reproduction; Elsevier: Amsterdam, The Netherlands, 2022; pp. 245–399. [Google Scholar]
- Aguiar, S.S.; Sousa, C.V.; Deus, L.A.; Rosa, T.S.; Sales, M.M.; Neves, R.V.P.; Barbosa, L.P.; Santos, P.A.; Campbell, C.S.; Simões, H.G.; et al. Oxidative stress, inflammatory cytokines and body composition of master athletes: The interplay. Exp. Gerontol. 2020, 130, 110806. [Google Scholar] [CrossRef]
- El-Missiry, M.A.; Fekri, A.; Kesar, L.A.; Othman, A.I. Polyphenols are potential nutritional adjuvants for targeting COVID-19. Phyther. Res. 2021, 35, 2879–2889. [Google Scholar] [CrossRef]
- Yahfoufi, N.; Alsadi, N.; Jambi, M.; Matar, C. The immunomodulatory and anti-inflammatory role of polyphenols. Nutrients 2018, 10, 1618. [Google Scholar] [CrossRef] [Green Version]
- Findeisen, H.M.; Pearson, K.J.; Gizard, F.; Zhao, Y.; Qing, H.; Jones, K.L.; Cohn, D.; Heywood, E.B.; de Cabo, R.; Bruemmer, D.; et al. Oxidative Stress Accumulates in Adipose Tissue during Aging and Inhibits Adipogenesis. PLoS ONE 2011, 6, e18532. [Google Scholar] [CrossRef]
- Luo, J.; Si, H.; Jia, Z.; Liu, D. Dietary anti-aging polyphenols and potential mechanisms. Antioxidants 2021, 10, 283. [Google Scholar] [CrossRef]
- Kobayashi, M.; Yamamoto, M. Molecular Mechanisms Activating the Nrf2-Keap1 Pathway of Antioxidant Gene Regulation. Antioxid. Redox Signal. 2005, 7, 385–394. [Google Scholar] [CrossRef] [PubMed]
- Pallauf, K.; Duckstein, N.; Hasler, M.; Klotz, L.-O.; Rimbach, G. Flavonoids as Putative Inducers of the Transcription Factors Nrf2, FoxO, and PPAR γ. Oxid. Med. Cell. Longev. 2017, 2017, 1–11. [Google Scholar] [CrossRef]
- Santangelo, C.; Varì, R.; Scazzocchio, B.; di Benedetto, R.; Filesi, C.; Masella, R. Polyphenols, intracellular signalling and inflammation. Ann. Ist. Super. Sanita 2007, 43, 394–405. [Google Scholar] [PubMed]
- Boskey, A.L.; Coleman, R. Aging and bone. J. Dent. Res. 2010, 89, 1333–1348. [Google Scholar] [CrossRef] [PubMed]
- Austermann, K.; Baecker, N.; Stehle, P.; Heer, M. Putative Effects of Nutritive Polyphenols on Bone Metabolism In Vivo—Evidence from Human Studies. Nutrients 2019, 11, 871. [Google Scholar] [CrossRef]
- Banfi, G.; Iorio, E.L.; Corsi, M.M. Oxidative stress, free radicals and bone remodeling. Clin. Chem. Lab. Med. 2008, 46, 1550–1555. [Google Scholar] [CrossRef]
- Grzesik, W.J.; Frazier, C.R.; Shapiro, J.R.; Sponseller, P.D.; Robey, P.G.; Fedarko, N.S. Age-related Changes in Human Bone Proteoglycan Structure. J. Biol. Chem. 2002, 277, 43638–43647. [Google Scholar] [CrossRef]
- Sharma, A.R.; Nam, J.-S. Kaempferol stimulates WNT/β-catenin signaling pathway to induce differentiation of osteoblasts. J. Nutr. Biochem. 2019, 74, 108228. [Google Scholar] [CrossRef]
- Xu, X.; Fan, X.; Wu, X.; Shi, L.; Wang, P.; Gao, F.; Sun, W.; Li, Z. Protective effect of Kaempferol on endothelial cell injury in glucocorticoid induced osteonecrosis of the femoral head. Chin. J. Reparative Reconstr. Surg. 2022, 36, 1277–1287. [Google Scholar] [CrossRef]
- Adhikary, S.; Choudhary, D.; Ahmad, N.; Karvande, A.; Kumar, A.; Banala, V.T.; Mishra, P.R.; Trivedi, R. Dietary flavonoid kaempferol inhibits glucocorticoid-induced bone loss by promoting osteoblast survival. Nutrition 2018, 53, 64–76. [Google Scholar] [CrossRef] [PubMed]
- Tsuchiya, S.; Sugimoto, K.; Kamio, H.; Okabe, K.; Kuroda, K.; Okido, M.; Hibi, H. Kaempferol-immobilized titanium dioxide promotes formation of new bone: Effects of loading methods on bone marrow stromal cell differentiation in vivo and in vitro. Int. J. Nanomed. 2018, 13, 1665–1676. [Google Scholar] [CrossRef]
- Doss, H.M.; Samarpita, S.; Ganesan, R.; Rasool, M. Ferulic acid, a dietary polyphenol suppresses osteoclast differentiation and bone erosion via the inhibition of RANKL dependent NF-κB signalling pathway. Life Sci. 2018, 207, 284–295. [Google Scholar] [CrossRef]
- Hou, T.; Zhang, L.; Yang, X. Ferulic acid, a natural polyphenol, protects against osteoporosis by activating SIRT1 and NF-κB in neonatal rats with glucocorticoid-induced osteoporosis. Biomed. Pharmacother. 2019, 120, 109205. [Google Scholar] [CrossRef]
- Balagangadharan, K.; Trivedi, R.; Vairamani, M.; Selvamurugan, N. Sinapic acid-loaded chitosan nanoparticles in polycaprolactone electrospun fibers for bone regeneration in vitro and in vivo. Carbohydr. Polym. 2019, 216, 1–16. [Google Scholar] [CrossRef]
- Bhattarai, G.; Jeon, Y.-M.; Choi, K.-C.; Wagle, S.; Sim, H.-J.; Kim, J.-I.; Zhao, S.; Kim, J.-G.; Cho, E.-S.; Kook, S.-H.; et al. Functional improvement of collagen-based bioscaffold to enhance periodontal-defect healing via combination with dietary antioxidant and COMP-angiopoietin 1. Biomater. Adv. 2022, 135, 112673. [Google Scholar] [CrossRef]
- Filho, A.C.M.L.; Silva, I.S.; Sousa, F.B.M.; de Souza, L.K.M.; da Gomes, B.S.; Gonçalves, R.L.G.; de Rezende, D.C.; Cunha, F.V.M.; Wong, D.V.T.; Júnior, R.C.P.L.; et al. Inhibition of neutrophil migration and reduction of oxidative stress by ethyl p-coumarate in acute and chronic inflammatory models. Phytomedicine 2019, 57, 9–17. [Google Scholar] [CrossRef]
- Cheng, Y.-H.; Yang, S.-H.; Liu, C.-C.; Gefen, A.; Lin, F.-H. Thermosensitive hydrogel made of ferulic acid-gelatin and chitosan glycerophosphate. Carbohydr. Polym. 2013, 92, 1512–1519. [Google Scholar] [CrossRef]
- Jiao, R.; Chen, H.; Wan, Q.; Zhang, X.; Dai, J.; Li, X.; Yan, L.; Sun, Y. Apigenin inhibits fibroblast proliferation and reduces epidural fibrosis by regulating Wnt3a/β-catenin signaling pathway. J. Orthop. Surg. Res. 2019, 14, 258. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Zhao, Q.; Liu, T.; Zhao, H.; Wang, R.; Li, H.; Zhang, Y.; Shan, L.; He, B.; Wang, X.; et al. Effect of Vicenin-2 on ovariectomy-induced osteoporosis in rats. Biomed. Pharmacother. 2020, 129, 110474. [Google Scholar] [CrossRef]
- Ranjbar, F.E.; Foroutan, F.; Hajian, M.; Ai, J.; Farsinejad, A.; Ebrahimi-Barough, S.; Dehghan, M.M.; Azami, M. Preparation and characterization of 58S bioactive glass based scaffold with Kaempferol-containing Zein coating for bone tissue engineering. J. Biomed. Mater. Res. B Appl. Biomater. 2021, 109, 1259–1270. [Google Scholar] [CrossRef]
- Tieland, M.; Trouwborst, I.; Clark, B.C. Skeletal muscle performance and ageing. J. Cachexia Sarcopenia Muscle 2018, 9, 3–19. [Google Scholar] [CrossRef] [PubMed]
- Stump, C.S.; Henriksen, E.J.; Wei, Y.; Sowers, J.R. The metabolic syndrome: Role of skeletal muscle metabolism. Ann. Med. 2006, 38, 389–402. [Google Scholar] [CrossRef]
- Schnyder, S.; Handschin, C. Skeletal muscle as an endocrine organ: PGC-1α, myokines and exercise. Bone 2015, 80, 115–125. [Google Scholar] [CrossRef] [PubMed]
- Nikawa, T.; Ulla, A.; Sakakibara, I. Polyphenols and Their Effects on Muscle Atrophy and Muscle Health. Molecules 2021, 26, 4887. [Google Scholar] [CrossRef] [PubMed]
- Alkhalidy, H.; Moore, W.; Wang, Y.; Luo, J.; McMillan, R.; Zhen, W.; Zhou, K.; Liu, D. The Flavonoid Kaempferol Ameliorates Streptozotocin-Induced Diabetes by Suppressing Hepatic Glucose Production. Molecules 2018, 23, 2338. [Google Scholar] [CrossRef]
- Da Silva, D.; Casanova, L.M.; Marcondes, M.C.; Espindola-Netto, J.M.; Paixão, L.P.; de Melo, G.O.; Zancan, P.; Sola-Penna, M.; Costa, S.S. Antidiabetic activity of Sedum dendroideum: Metabolic enzymes as putative targets for the bioactive flavonoid kaempferitrin. IUBMB Life 2014, 66, 361–370. [Google Scholar] [CrossRef] [PubMed]
- Kashyap, B.; Saikia, K.; Samanta, S.K.; Thakur, D.; Banerjee, S.K.; Borah, J.C.; Talukdar, N.C. Kaempferol 3-O-rutinoside from Antidesma acidum Retz. Stimulates glucose uptake through SIRT1 induction followed by GLUT4 translocation in skeletal muscle L6 cells. J. Ethnopharmacol. 2022, 301, 115788. [Google Scholar] [CrossRef]
- Kitakaze, T.; Jiang, H.; Nomura, T.; Hironao, K.; Yamashita, Y.; Ashida, H. Kaempferol Promotes Glucose Uptake in Myotubes through a JAK2-Dependent Pathway. J. Agric. Food Chem. 2020, 68, 13720–13729. [Google Scholar] [CrossRef]
- Roy, J.R.; Janaki, C.S.; Jayaraman, S.; Periyasamy, V.; Balaji, T.; Vijayamalathi, M.; Veeraraghavan, V.P. Carica papaya Reduces Muscle Insulin Resistance via IR/GLUT4 Mediated Signaling Mechanisms in High Fat Diet and Streptozotocin-Induced Type-2 Diabetic Rats. Antioxidants 2022, 11, 2081. [Google Scholar] [CrossRef] [PubMed]
- Akiyama, M.; Mizokami, T.; Miyamoto, S.; Ikeda, Y. Kaempferol increases intracellular ATP content in C2C12 myotubes under hypoxic conditions by suppressing the HIF-1α stabilization and/or by enhancing the mitochondrial complex IV activity. J. Nutr. Biochem. 2022, 103, 108949. [Google Scholar] [CrossRef]
- Wen, Y.; Ushio, H. Ferulic Acid Promotes Hypertrophic Growth of Fast Skeletal Muscle in Zebrafish Model. Nutrients 2017, 9, 1066. [Google Scholar] [CrossRef] [PubMed]
- Yin, X.; Liu, W.; Chen, H.; Qi, C.; Chen, H.; Niu, H.; Yang, J.; Kwok, K.W.H.; Dong, W. Effects of ferulic acid on muscle development and intestinal microbiota of zebrafish. J. Anim. Physiol. Anim. Nutr. 2022, 106, 429–440. [Google Scholar] [CrossRef]
- Kuppusamy, P.; Soundharrajan, I.; Kim, D.H.; Hwang, I.; Choi, K.C. 4-hydroxy-3-methoxy cinnamic acid accelerate myoblasts differentiation on C2C12 mouse skeletal muscle cells via AKT and ERK 1/2 activation. Phytomedicine 2019, 60, 152873. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Chen, X.; Huang, Z.; Chen, D.; Yu, B.; Chen, H.; Yu, J.; Luo, Y.; Zheng, P.; He, J. Effects of dietary ferulic acid supplementation on growth performance and skeletal muscle fiber type conversion in weaned piglets. J. Sci. Food Agric. 2021, 101, 5116–5123. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Guo, Y.; Jia, G.; Zhao, H.; Liu, G.; Huang, Z. Ferulic acid regulates muscle fiber type formation through the Sirt1/AMPK signaling pathway. Food Funct. 2019, 10, 259–265. [Google Scholar] [CrossRef]
- Ilavenil, S.; Kim, D.; Srigopalram, S.; Arasu, M.; Lee, K.; Lee, J.; Lee, J.; Renganathan, S.; Choi, K. Potential Application of p-Coumaric Acid on Differentiation of C2C12 Skeletal Muscle and 3T3-L1 Preadipocytes—An in Vitro and in Silico Approach. Molecules 2016, 21, 997. [Google Scholar] [CrossRef]
- Gogoi, B.; Chatterjee, P.; Mukherjee, S.; Buragohain, A.K.; Bhattacharya, S.; Dasgupta, S. A polyphenol rescues lipid induced insulin resistance in skeletal muscle cells and adipocytes. Biochem. Biophys. Res. Commun. 2014, 452, 382–388. [Google Scholar] [CrossRef]
- Kang, B.; Chiang, B. Amelioration of insulin resistance using the additive effect of ferulic acid and resveratrol on vesicle trafficking for skeletal muscle glucose metabolism. Phyther. Res. 2020, 34, 808–816. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.S.B.; Patil, K.N. Trans-ferulic acid attenuates hyperglycemia-induced oxidative stress and modulates glucose metabolism by activating AMPK signaling pathway in vitro. J. Food Biochem. 2022, 46, e14038. [Google Scholar] [CrossRef] [PubMed]
- Yoon, S.-A.; Kang, S.-I.; Shin, H.-S.; Kang, S.-W.; Kim, J.-H.; Ko, H.-C.; Kim, S.-J. P-Coumaric acid modulates glucose and lipid metabolism via AMP-activated protein kinase in L6 skeletal muscle cells. Biochem. Biophys. Res. Commun. 2013, 432, 553–557. [Google Scholar] [CrossRef] [PubMed]
- Shiota, C.; Abe, T.; Kawai, N.; Ohno, A.; Teshima-Kondo, S.; Mori, H.; Terao, J.; Tanaka, E.; Nikawa, T. Flavones Inhibit LPS-Induced Atrogin-1/MAFbx Expression in Mouse C2C12 Skeletal Myotubes. J. Nutr. Sci. Vitaminol. 2015, 61, 188–194. [Google Scholar] [CrossRef] [PubMed]
- Choi, W.H.; Jang, Y.J.; Son, H.J.; Ahn, J.; Jung, C.H.; Ha, T.Y. Apigenin inhibits sciatic nerve denervation–induced muscle atrophy. Muscle Nerve 2018, 58, 314–318. [Google Scholar] [CrossRef]
- Choi, W.H.; Son, H.J.; Jang, Y.J.; Ahn, J.; Jung, C.H.; Ha, T.Y. Apigenin Ameliorates the Obesity-Induced Skeletal Muscle Atrophy by Attenuating Mitochondrial Dysfunction in the Muscle of Obese Mice. Mol. Nutr. Food Res. 2017, 61, 1700218. [Google Scholar] [CrossRef]
- Jang, Y.J.; Son, H.J.; Choi, Y.M.; Ahn, J.; Jung, C.H.; Ha, T.Y. Apigenin enhances skeletal muscle hypertrophy and myoblast differentiation by regulating Prmt7. Oncotarget 2017, 8, 78300–78311. [Google Scholar] [CrossRef]
- Wang, D.; Yang, Y.; Zou, X.; Zhang, J.; Zheng, Z.; Wang, Z. Antioxidant Apigenin Relieves Age-Related Muscle Atrophy by Inhibiting Oxidative Stress and Hyperactive Mitophagy and Apoptosis in Skeletal Muscle of Mice. J. Gerontol. Ser. A 2020, 75, 2081–2088. [Google Scholar] [CrossRef]
- Ou, M.Y.; Zhang, H.; Tan, P.C.; Zhou, S.B.; Li, Q.F. Adipose tissue aging: Mechanisms and therapeutic implications. Cell Death Dis. 2022, 13, 300. [Google Scholar] [CrossRef]
- Cinti, S. Anatomy and physiology of the nutritional system. Mol. Aspects Med. 2019, 68, 101–107. [Google Scholar] [CrossRef]
- Cannon, B.; Nedergaard, J. Brown Adipose Tissue: Function and Physiological Significance. Physiol. Rev. 2004, 84, 277–359. [Google Scholar] [CrossRef]
- Kuk, J.L.; Saunders, T.J.; Davidson, L.E.; Ross, R. Age-related changes in total and regional fat distribution. Ageing Res. Rev. 2009, 8, 339–348. [Google Scholar] [CrossRef]
- Ouchi, N.; Parker, J.L.; Lugus, J.J.; Walsh, K. Adipokines in inflammation and metabolic disease. Nat. Rev. Immunol. 2011, 11, 85–97. [Google Scholar] [CrossRef]
- Ilich, J.Z.; Kelly, O.J.; Kim, Y.; Spicer, M.T. Low-grade chronic inflammation perpetuated by modern diet as a promoter of obesity and osteoporosis. Arch. Ind. Hyg. Toxicol. 2014, 65, 139–148. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.; Kwon, M.; Choi, J.S.; Jeong, H.O.; Chung, H.Y.; Kim, H.-R. Kaempferol Isolated from Nelumbo nucifera Inhibits Lipid Accumulation and Increases Fatty Acid Oxidation Signaling in Adipocytes. J. Med. Food 2015, 18, 1363–1370. [Google Scholar] [CrossRef]
- Li, H.; Kang, J.-H.; Han, J.-M.; Cho, M.-H.; Chung, Y.-J.; Park, K.H.; Shin, D.-H.; Park, H.-Y.; Choi, M.-S.; Jeong, T.-S.; et al. Anti-Obesity Effects of Soy Leaf via Regulation of Adipogenic Transcription Factors and Fat Oxidation in Diet-Induced Obese Mice and 3T3-L1 Adipocytes. J. Med. Food 2015, 18, 899–908. [Google Scholar] [CrossRef]
- Mehanna, E.T.; El-sayed, N.M.; Ibrahim, A.K.; Ahmed, S.A.; Abo-Elmatty, D.M. Isolated compounds from Cuscuta pedicellata ameliorate oxidative stress and upregulate expression of some energy regulatory genes in high fat diet induced obesity in rats. Biomed. Pharmacother. 2018, 108, 1253–1258. [Google Scholar] [CrossRef]
- Chaves, W.F.; Pinheiro, I.L.; da Silva, L.O.; Lima-Oliveira, D.P.; de Muniz, G.S.; do Barreto, Á.N.; da Silva, B.J.; Manhães-de-Castro, R.; da Silva Aragão, R. Neonatal administration of kaempferol does not alter satiety but increases somatic growth and reduces adiposity in offspring of high-fat diet dams. Life Sci. 2020, 259, 118224. [Google Scholar] [CrossRef]
- Barge, S.; Deka, B.; Kashyap, B.; Bharadwaj, S.; Kandimalla, R.; Ghosh, A.; Dutta, P.P.; Samanta, S.K.; Manna, P.; Borah, J.C.; et al. Astragalin mediates the pharmacological effects of Lysimachia candida Lindl on adipogenesis via downregulating PPARG and FKBP51 signaling cascade. Phyther. Res. 2021, 35, 6990–7003. [Google Scholar] [CrossRef] [PubMed]
- Bian, Y.; Lei, J.; Zhong, J.; Wang, B.; Wan, Y.; Li, J.; Liao, C.; He, Y.; Liu, Z.; Ito, K.; et al. Kaempferol reduces obesity, prevents intestinal inflammation, and modulates gut microbiota in high-fat diet mice. J. Nutr. Biochem. 2022, 99, 108840. [Google Scholar] [CrossRef] [PubMed]
- Kitamura, H.; Saito, N.; Fujimoto, J.; Nakashima, K.; Fujikura, D. Brazilian propolis ethanol extract and its component kaempferol induce myeloid-derived suppressor cells from macrophages of mice in vivo and in vitro. BMC Complement. Altern. Med. 2018, 18, 138. [Google Scholar] [CrossRef] [Green Version]
- Han, X.; Guo, J.; You, Y.; Zhan, J.; Huang, W. p-Coumaric acid prevents obesity via activating thermogenesis in brown adipose tissue mediated by mTORC1-RPS6. FASEB J. 2020, 34, 7810–7824. [Google Scholar] [CrossRef]
- Yoon, D.S.; Cho, S.Y.; Yoon, H.J.; Kim, S.R.; Jung, U.J. Protective effects of p-coumaric acid against high-fat diet-induced metabolic dysregulation in mice. Biomed. Pharmacother. 2021, 142, 111969. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Pan, Y.; Wang, L.; Zhou, H.; Song, G.; Wang, Y.; Liu, J.; Li, A. Optimal Dietary Ferulic Acid for Suppressing the Obesity-Related Disorders in Leptin-Deficient Obese C57BL/6J -ob/ob Mice. J. Agric. Food Chem. 2019, 67, 4250–4258. [Google Scholar] [CrossRef] [PubMed]
- Seo, S.H.; Jo, S.-M.; Truong, T.T.M.; Zhang, G.; Kim, D.-S.; Lee, M.; Lee, Y.; Kang, I. Peanut sprout rich in p -coumaric acid ameliorates obesity and lipopolysaccharide-induced inflammation and the inhibition of browning in adipocytes via mitochondrial activation. Food Funct. 2021, 12, 5361–5374. [Google Scholar] [CrossRef]
- Kuppusamy, P.; Ilavenil, S.; Hwang, I.H.; Kim, D.; Choi, K.C. Ferulic Acid Stimulates Adipocyte-Specific Secretory Proteins to Regulate Adipose Homeostasis in 3T3-L1 Adipocytes. Molecules 2021, 26, 1984. [Google Scholar] [CrossRef]
- Hossain, M.; Imran, K.M.; Rahman, M.S.; Yoon, D.; Marimuthu, V.; Kim, Y.-S. Sinapic acid induces the expression of thermogenic signature genes and lipolysis through activation of PKA/CREB signaling in brown adipocytes. BMB Rep. 2020, 53, 142–147. [Google Scholar] [CrossRef]
- Kinyua, A.W.; Ko, C.M.; Doan, K.V.; Yang, D.J.; Huynh, M.K.Q.; Moh, S.H.; Choi, Y.-H.; Kim, K.W. 4-hydroxy-3-methoxycinnamic acid regulates orexigenic peptides and hepatic glucose homeostasis through phosphorylation of FoxO1. Exp. Mol. Med. 2018, 50, e437. [Google Scholar] [CrossRef]
- Naowaboot, J.; Piyabhan, P.; Munkong, N.; Parklak, W.; Pannangpetch, P. Ferulic acid improves lipid and glucose homeostasis in high-fat diet-induced obese mice. Clin. Exp. Pharmacol. Physiol. 2016, 43, 242–250. [Google Scholar] [CrossRef]
- Cho, J.; Park, E. Ferulic acid maintains the self-renewal capacity of embryo stem cells and adipose-derived mesenchymal stem cells in high fat diet-induced obese mice. J. Nutr. Biochem. 2020, 77, 108327. [Google Scholar] [CrossRef] [PubMed]
- De Melo, T.S.; Lima, P.R.; Carvalho, K.M.M.B.; Fontenele, T.M.; Solon, F.R.N.; Tomé, A.R.; de Lemos, T.L.G.; da Cruz Fonseca, S.G.; Santos, F.A.; Rao, V.S.; et al. Ferulic acid lowers body weight and visceral fat accumulation via modulation of enzymatic, hormonal and inflammatory changes in a mouse model of high-fat diet-induced obesity. Braz. J. Med. Biol. Res. 2017, 50, e5630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van den Hoek-Hil, E.F.; van Schothorst, E.M.; van der Stelt, I.; Swarts, H.J.M.; van Vliet, M.; Amolo, T.; Vervoort, J.J.M.; Venema, D.; Hollman, P.C.H.; Rietjens, I.M.C.M.; et al. Direct comparison of metabolic health effects of the flavonoids quercetin, hesperetin, epicatechin, apigenin and anthocyanins in high-fat-diet-fed mice. Genes Nutr. 2015, 10, 23. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Liu, J.; Cai, S.; Wang, O.; Ji, B. Synergistic interactions of apigenin, naringin, quercetin and emodin on inhibition of 3T3-L1 preadipocyte differentiation and pancreas lipase activity. Obes. Res. Clin. Pract. 2016, 10, 327–339. [Google Scholar] [CrossRef] [PubMed]
- Gentile, D.; Fornai, M.; Colucci, R.; Pellegrini, C.; Tirotta, E.; Benvenuti, L.; Segnani, C.; Ippolito, C.; Duranti, E.; Virdis, A.; et al. The flavonoid compound apigenin prevents colonic inflammation and motor dysfunctions associated with high fat diet-induced obesity. PLoS ONE 2018, 13, e0195502. [Google Scholar] [CrossRef]
- Okla, M.; Al Madani, J.O.; Chung, S.; Alfayez, M. Apigenin Reverses Interleukin-1β-Induced Suppression of Adipocyte Browning via COX2/PGE2 Signaling Pathway in Human Adipocytes. Mol. Nutr. Food Res. 2020, 64, 1900925. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.; Sun, Q.; Xu, W.; He, Y.; Jin, W.; Yuan, L.; Gao, R. Vitexin ameliorates high fat diet-induced obesity in male C57BL/6J mice via the AMPKα-mediated pathway. Food Funct. 2019, 10, 1940–1947. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.-S.; Qu, W. Dietary Apigenin promotes lipid catabolism, thermogenesis, and browning in adipose tissues of HFD-Fed mice. Food Chem. Toxicol. 2019, 133, 110780. [Google Scholar] [CrossRef]
- Su, T.; Huang, C.; Yang, C.; Jiang, T.; Su, J.; Chen, M.; Fatima, S.; Gong, R.; Hu, X.; Bian, Z.; et al. Apigenin inhibits STAT3/CD36 signaling axis and reduces visceral obesity. Pharmacol. Res. 2020, 152, 104586. [Google Scholar] [CrossRef]
- Sreekumar, S.; Vijayan, V.; Singh, F.; Sudhakar, M.; Lakra, R.; Korrapati, P.S.; Kiran, M.S. White to brown adipocyte transition mediated by Apigenin via VEGF-PRDM16 signaling. J. Cell. Biochem. 2022, 123, 1793–1807. [Google Scholar] [CrossRef]
- Feng, X.; Weng, D.; Zhou, F.; Owen, Y.D.; Qin, H.; Zhao, J.; Yu, W.; Huang, Y.; Chen, J.; Fu, H.; et al. Activation of PPARγ by a Natural Flavonoid Modulator, Apigenin Ameliorates Obesity-Related Inflammation Via Regulation of Macrophage Polarization. Ebio Med. 2016, 9, 61–76. [Google Scholar] [CrossRef]
- Wong, S.K.; Chin, K.-Y.; Ima-Nirwana, S. Quercetin as an Agent for Protecting the Bone: A Review of the Current Evidence. Int. J. Mol. Sci. 2020, 21, 6448. [Google Scholar] [CrossRef]
- Lin, W.; Zhao, Y.; Liu, C.; Yan, Y.; Ou, Q. Quercetin supplementation and muscular atrophy in animal models: A systematic review and meta-analysis. Int. J. Food Prop. 2022, 25, 2166–2183. [Google Scholar] [CrossRef]
- Sato, S.; Mukai, Y. Modulation of Chronic Inflammation by Quercetin: The Beneficial Effects on Obesity. J. Inflamm. Res. 2020, 13, 421–431. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound(s) | Effects | Reference |
|---|---|---|
| Kaempferol | Induction of SaOS-2 osteoblast differentiation | [43] |
| Promotion of osteoblast survival in glucocorticoid induced osteonecrosis on human bone microvascular endothelial cells and in vivo in female ovariectomized rats | [44,45] | |
| Promotion of bone regeneration in rat bone marrow stromal cells and in vivo in rat femur bones | [46] | |
| Hydroxycinnamic acids | Supression of bone erosion and osteoporosis in murine bone marrow cells and RAW 264.7 macrophage cell line; in vivo in neonatal rats with induced osteoporosis | [47,48] |
| Promotion of bone regeneration in a critical-sized rat calvarial bone defect model system and in a rat model of critically sized mandible defects | [49,50] | |
| Inhibition of oxidative stress and inflammation related to arthritis and disc degradation in nucleus pulposus cells and in acute and chronic models of inflammation | [51,52] | |
| Apigenin | Reduction of epidural fibrosis after spine surgery in human fibroblasts and in vivo in Sprague-Dawley rats | [53] |
| Treatment of osteoporosis in ovariectomy-induced osteoporosis rats | [54] |
| Compound(s) | Effects | Reference |
|---|---|---|
| Kaempferol | Antidiabetic on L6 murine skeletal muscle cells and in vivo in streptozotocin-induced diabetic rats | [60,61,62,63,64] |
| Increase of ATP content in myotubes at hypoxic conditions | [65] | |
| Hydroxycinnamic acids | Muscle growth promotion on C2C12 murine skeletal muscle cells and in vivo in zebrafish models | [66,67,68] |
| Modification of muscle fiber type on C2C12 murine skeletal muscle cells and in vivo in weaned piglets | [69,70] | |
| Influence on skeletal muscle differentiation in C2C12 murine skeletal muscle cells | [68,71] | |
| Antidiabetic on L6 murine skeletal muscle cells | [72,73,74,75] | |
| Apigenin | Suppression of muscle atrophy on C2C12 murine skeletal muscle cells and in vivo in mice | [76,77,78,79,80] |
| Compound(s) | Effects | Reference |
|---|---|---|
| Kaempferol | Antiobesity on murine adipocytes and in vivo in obese mice | [87,88,89,90,91,92] |
| Antidiabetic on murine adipocytes and in vivo in obese C57BL/6 mice | [87,89,93] | |
| Anti-inflammatory on macrophage-like cell line J774.1 and in vivo in obese mice | [92,93] | |
| Hydroxycinnamic acids | Antiobesity on human and murine adipocytes and in vivo in obese mice | [94,95,96,97,98,99,100,101,102,103] |
| Antidiabetic on murine adipocytes and in vivo in obese and diabetic mice | [72,95,100,101] | |
| Anti-inflammatory on murine 3T3-L1 adipocytes and lipopolysaccharides (LPS)-induced macrophages; in vivo in obese mice | [72,95,97] | |
| Management of NAFLD a in vivo in obese mice | [95] | |
| Apigenin | Antiobesity on murine 3T3-L1 and human adipocytes and in vivo in obese mice | [104,105,106,107,108,109,110,111] |
| Anti-inflammatory on human adipocytes and in vivo in obese mice | [106,107,112] | |
| Antidiabetic in vivo in obese mice | [109,112] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dobroslavić, E.; Elez Garofulić, I.; Ilich, J.Z. Potential of Laurel (Laurus nobilis L.) Leaf Polyphenols for Modulation of Body Composition. Appl. Sci. 2023, 13, 2275. https://doi.org/10.3390/app13042275
Dobroslavić E, Elez Garofulić I, Ilich JZ. Potential of Laurel (Laurus nobilis L.) Leaf Polyphenols for Modulation of Body Composition. Applied Sciences. 2023; 13(4):2275. https://doi.org/10.3390/app13042275
Chicago/Turabian StyleDobroslavić, Erika, Ivona Elez Garofulić, and Jasminka Z. Ilich. 2023. "Potential of Laurel (Laurus nobilis L.) Leaf Polyphenols for Modulation of Body Composition" Applied Sciences 13, no. 4: 2275. https://doi.org/10.3390/app13042275