
Effective Connectivity Changes among Brain Hierarchical Architecture of Pre-Supplementary Motor Area in Taxi Drivers

Abstract
:Featured Application
Abstract
1. Introduction
- We propose a general framework for investigating functional integration of the human brain. This framework takes functional connectivity results as a prior, enabling subsequent DCM to assess causal organization of brain network at a whole-brain scale.
- We check the quality of DCM model fitting. The results have indicated that our framework can effectively discover the specific brain network integration that best explains the observed BOLD fluctuations.
- We apply this framework to investigate the functional plasticity of the resting human brain underlying driving experience, and, for the first time, we discover a hierarchical architecture with pre-SMA located at a higher level, and reveal that several directed connections in the hierarchy exhibit significant classification ability to distinguish drivers from nondrivers.
2. Materials and Methods
2.1. Subjects and Data Acquisition
2.2. Data Preprocessing
2.3. Functional Connectivity Analysis
2.4. Effective Connectivity Analysis
2.4.1. Spectral Dynamic Causal Modeling
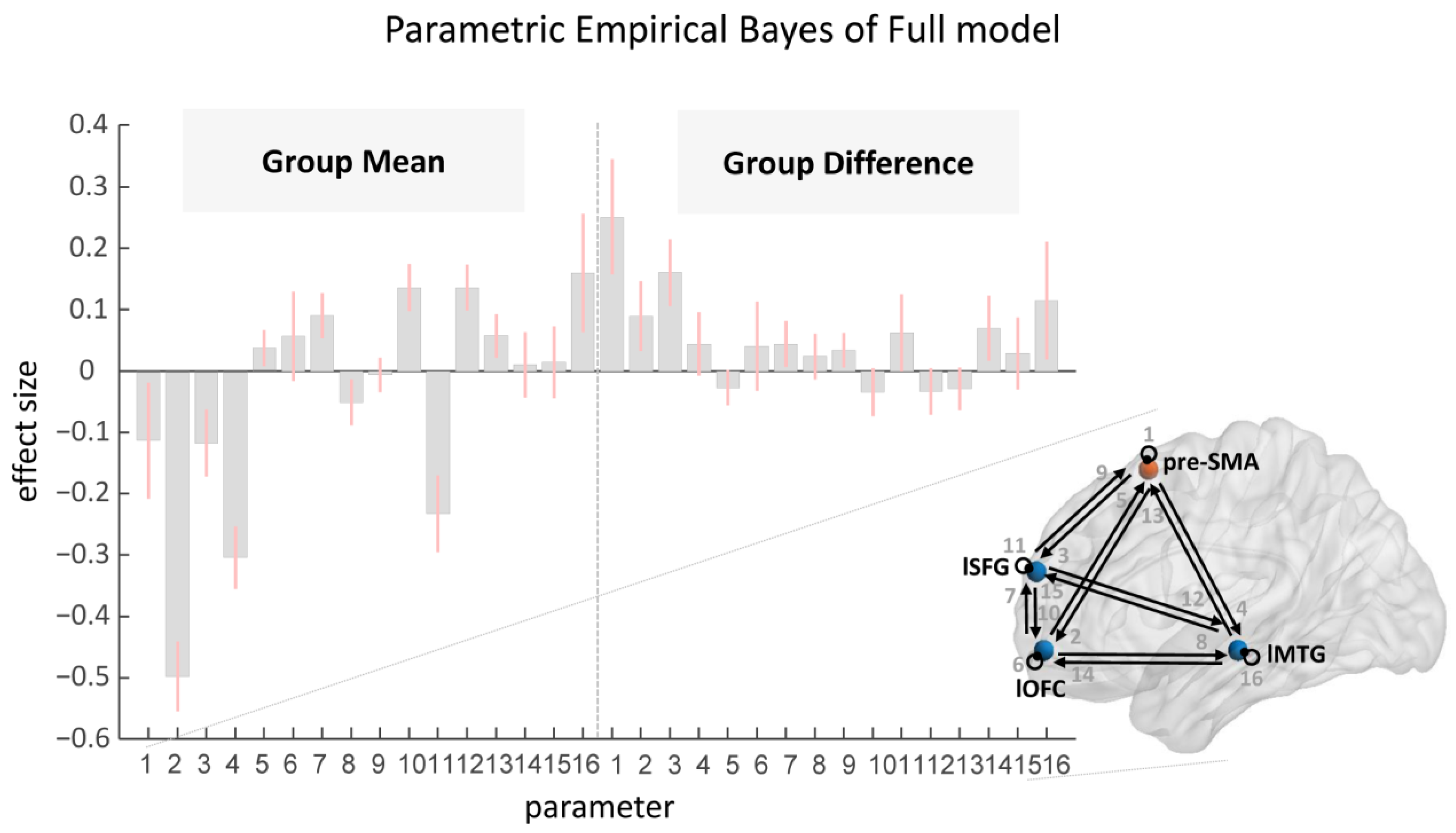
2.4.2. Parametric Empirical Bayes
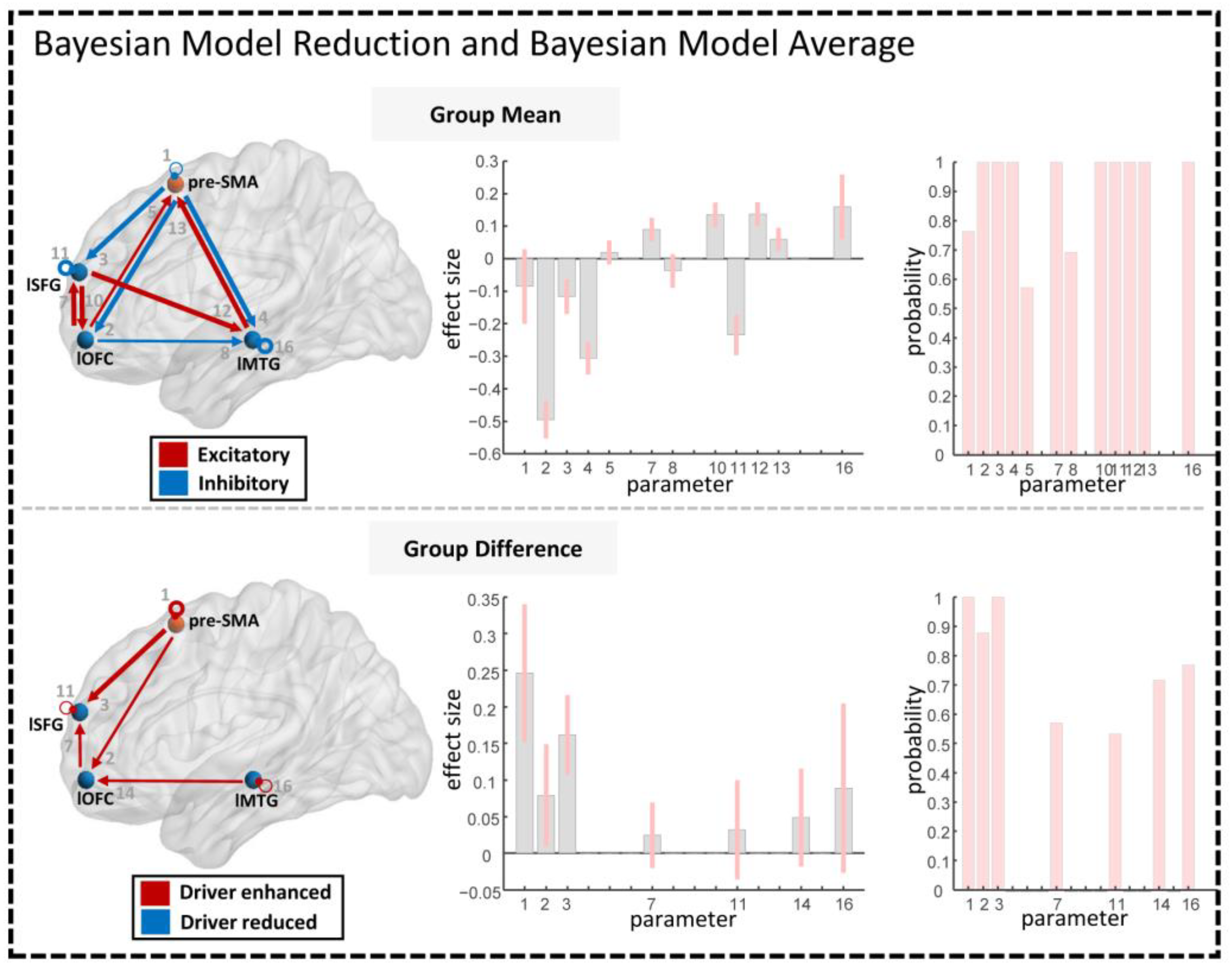
2.4.3. Bayesian Model Reduction and Bayesian Model Averaging
3. Results
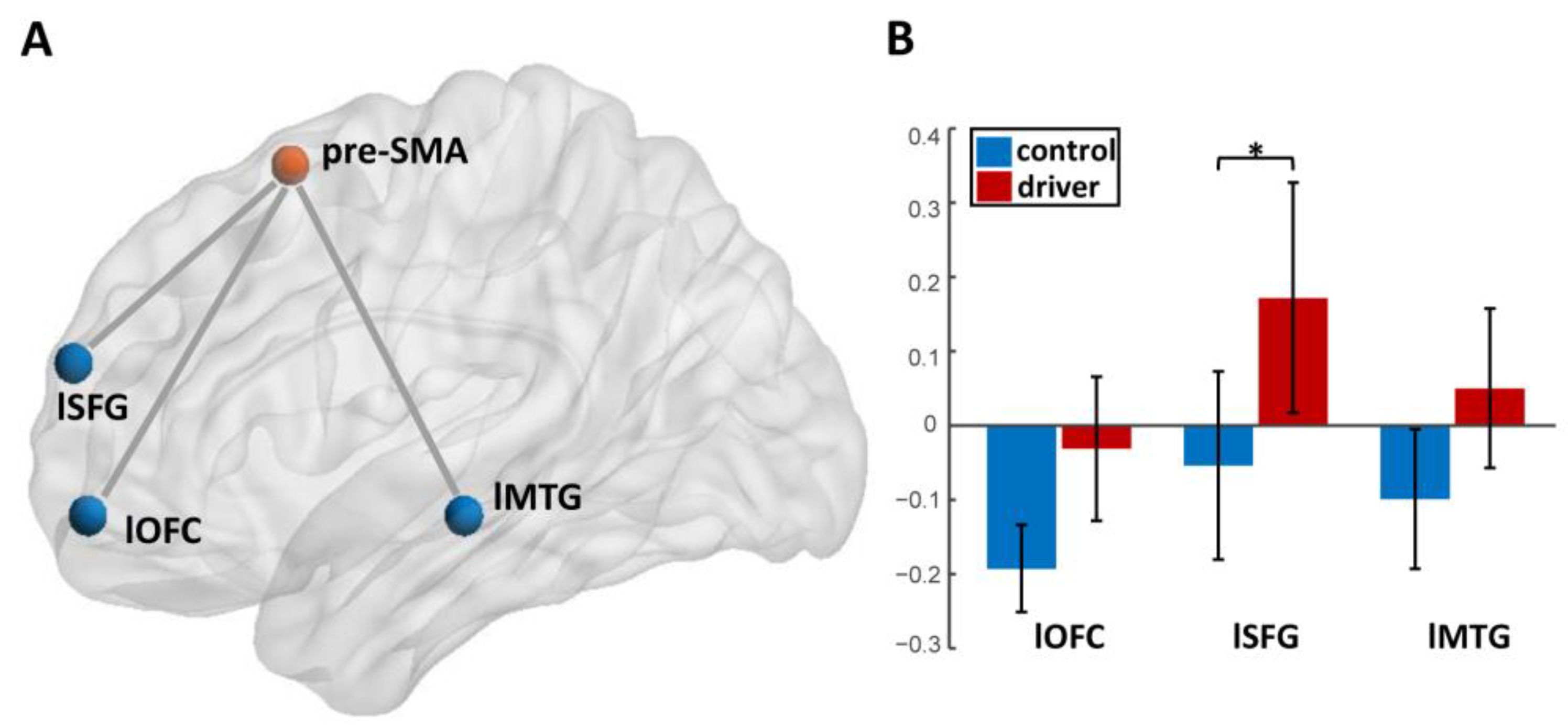
3.1. Group Differences of Functional Connectivity
3.2. Group Differences of Effective Connectivity
3.3. Significance Evaluation of Effective Connectivity
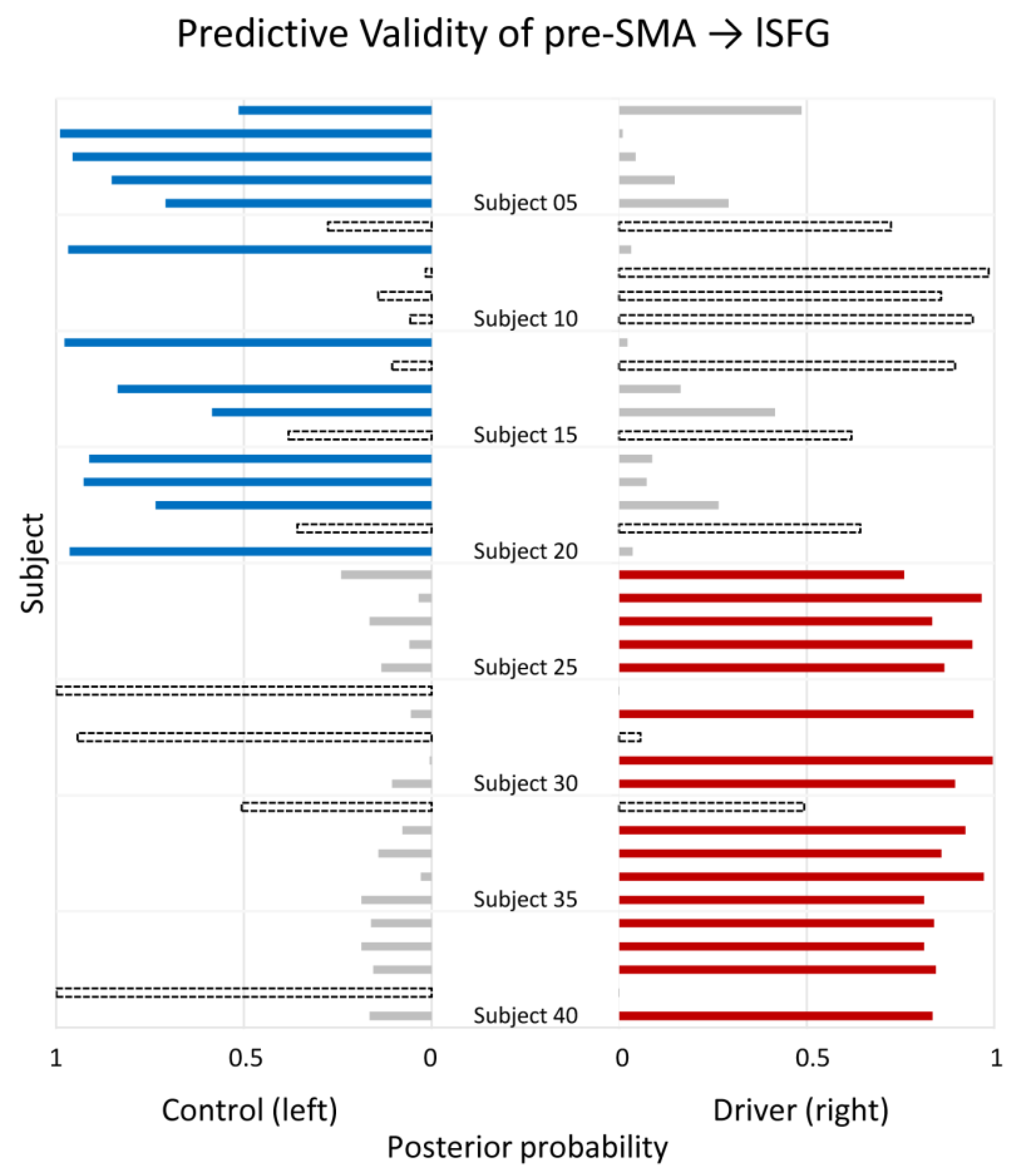
3.4. Predictive Validity Assessment of Effective Connectivity
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Calhoun, V.D.; Pekar, J.J.; McGinty, V.B.; Adali, T.; Watson, T.D.; Pearlson, G.D. Different activation dynamics in multiple neural systems during simulated driving. Hum. Brain Mapp. 2002, 16, 158–167. [Google Scholar] [CrossRef] [PubMed]
- Callan, A.M.; Osu, R.; Yamagishi, Y.; Callan, D.E.; Inoue, N. Neural correlates of resolving uncertainty in driver’s decision making. Hum. Brain Mapp. 2009, 30, 2804–2812. [Google Scholar] [CrossRef] [PubMed]
- Horikawa, E.; Okamura, N.; Tashiro, M.; Sakurada, Y.; Maruyama, M.; Arai, H.; Yamaguchi, K.; Sasaki, H.; Yanai, K.; Itoh, M. The neural correlates of driving performance identified using positron emission tomography. Brain Cogn. 2005, 58, 166–171. [Google Scholar] [CrossRef]
- Spiers, H.J.; Maguire, E.A. Neural substrates of driving behaviour. NeuroImage 2007, 36, 245–255. [Google Scholar] [CrossRef]
- Uchiyama, Y.; Ebe, K.; Kozato, A.; Okada, T.; Sadato, N. The neural substrates of driving at a safe distance: A functional MRI study. Neurosci. Lett. 2003, 352, 199–202. [Google Scholar] [CrossRef]
- Walter, H.; Vetter, S.C.; Grothe, J.; Wunderlich, A.P.; Hahn, S.; Spitzer, M. The neural correlates of driving. Neuroreport 2001, 12, 1763–1767. [Google Scholar] [CrossRef] [PubMed]
- Damoiseaux, J.S.; Beckmann, C.F.; Arigita, E.J.; Barkhof, F.; Scheltens, P.; Stam, C.J.; Smith, S.M.; Rombouts, S.A. Reduced resting-state brain activity in the “default network” in normal aging. Cereb. Cortex 2008, 18, 1856–1864. [Google Scholar] [CrossRef]
- Hampson, M.; Driesen, N.R.; Skudlarski, P.; Gore, J.C.; Constable, R.T. Brain connectivity related to working memory performance. J. Neurosci. Off. J. Soc. Neurosci. 2006, 26, 13338–13343. [Google Scholar] [CrossRef]
- Seeley, W.W.; Menon, V.; Schatzberg, A.F.; Keller, J.; Glover, G.H.; Kenna, H.; Reiss, A.L.; Greicius, M.D. Dissociable intrinsic connectivity networks for salience processing and executive control. J. Neurosci. Off. J. Soc. Neurosci. 2007, 27, 2349–2356. [Google Scholar] [CrossRef]
- Albert, N.B.; Robertson, E.M.; Miall, R.C. The resting human brain and motor learning. Curr. Biol. CB 2009, 19, 1023–1027. [Google Scholar] [CrossRef]
- Duan, X.; He, S.; Liao, W.; Liang, D.; Qiu, L.; Wei, L.; Li, Y.; Liu, C.; Gong, Q.; Chen, H. Reduced caudate volume and enhanced striatal-DMN integration in chess experts. NeuroImage 2012, 60, 1280–1286. [Google Scholar] [CrossRef]
- Lewis, C.M.; Baldassarre, A.; Committeri, G.; Romani, G.L.; Corbetta, M. Learning sculpts the spontaneous activity of the resting human brain. Proc. Natl. Acad. Sci. USA 2009, 106, 17558–17563. [Google Scholar] [CrossRef] [PubMed]
- Taubert, M.; Lohmann, G.; Margulies, D.S.; Villringer, A.; Ragert, P. Long-term effects of motor training on resting-state networks and underlying brain structure. NeuroImage 2011, 57, 1492–1498. [Google Scholar] [CrossRef] [PubMed]
- Michon, J.A. A critical view of driver behavior models: What do. In Human Behavior and Traffic Safety; Springer: Berlin/Heidelberg, Germany, 1985; pp. 485–524. [Google Scholar]
- Biswal, B.; Yetkin, F.Z.; Haughton, V.M.; Hyde, J.S. Functional connectivity in the motor cortex of resting human brain using echo-planar MRI. Magn. Reson. Med. 1995, 34, 537–541. [Google Scholar] [CrossRef] [PubMed]
- Damoiseaux, J.S.; Rombouts, S.A.; Barkhof, F.; Scheltens, P.; Stam, C.J.; Smith, S.M.; Beckmann, C.F. Consistent resting-state networks across healthy subjects. Proc. Natl. Acad. Sci. USA 2006, 103, 13848–13853. [Google Scholar] [CrossRef] [PubMed]
- Fox, M.D.; Raichle, M.E. Spontaneous fluctuations in brain activity observed with functional magnetic resonance imaging. Nat. Rev. Neurosci. 2007, 8, 700–711. [Google Scholar] [CrossRef]
- Greicius, M.D.; Krasnow, B.; Reiss, A.L.; Menon, V. Functional connectivity in the resting brain: A network analysis of the default mode hypothesis. Proc. Natl. Acad. Sci. USA 2003, 100, 253–258. [Google Scholar] [CrossRef]
- Friston, K.J.; Harrison, L.; Penny, W. Dynamic causal modelling. Neuroimage 2003, 19, 1273–1302. [Google Scholar] [CrossRef]
- Razi, A.; Seghier, M.L.; Zhou, Y.; McColgan, P.; Zeidman, P.; Park, H.J.; Sporns, O.; Rees, G.; Friston, K.J. Large-scale DCMs for resting-state fMRI. Netw. Neurosci. 2017, 1, 222–241. [Google Scholar] [CrossRef]
- Seghier, M.L.; Friston, K.J. Network discovery with large DCMs. NeuroImage 2013, 68, 181–191. [Google Scholar] [CrossRef]
- Cohen, A.L.; Fair, D.A.; Dosenbach, N.U.; Miezin, F.M.; Dierker, D.; Van Essen, D.C.; Schlaggar, B.L.; Petersen, S.E. Defining functional areas in individual human brains using resting functional connectivity MRI. NeuroImage 2008, 41, 45–57. [Google Scholar] [CrossRef] [PubMed]
- Craddock, R.C.; James, G.A.; Holtzheimer, P.E., 3rd; Hu, X.P.; Mayberg, H.S. A whole brain fMRI atlas generated via spatially constrained spectral clustering. Hum. Brain Mapp. 2012, 33, 1914–1928. [Google Scholar] [CrossRef] [PubMed]
- Nelson, S.M.; Cohen, A.L.; Power, J.D.; Wig, G.S.; Miezin, F.M.; Wheeler, M.E.; Velanova, K.; Donaldson, D.I.; Phillips, J.S.; Schlaggar, B.L.; et al. A parcellation scheme for human left lateral parietal cortex. Neuron 2010, 67, 156–170. [Google Scholar] [CrossRef]
- Shirer, W.R.; Ryali, S.; Rykhlevskaia, E.; Menon, V.; Greicius, M.D. Decoding subject-driven cognitive states with whole-brain connectivity patterns. Cereb. Cortex 2012, 22, 158–165. [Google Scholar] [CrossRef]
- Friston, K.J.; Kahan, J.; Biswal, B.; Razi, A. A DCM for resting state fMRI. NeuroImage 2014, 94, 396–407. [Google Scholar] [CrossRef] [PubMed]
- Razi, A.; Kahan, J.; Rees, G.; Friston, K.J. Construct validation of a DCM for resting state fMRI. NeuroImage 2015, 106, 1–14. [Google Scholar] [CrossRef]
- Friston, K.J.; Litvak, V.; Oswal, A.; Razi, A.; Stephan, K.E.; van Wijk, B.C.M.; Ziegler, G.; Zeidman, P. Bayesian model reduction and empirical Bayes for group (DCM) studies. NeuroImage 2016, 128, 413–431. [Google Scholar] [CrossRef]
- Zeidman, P.; Jafarian, A.; Corbin, N.; Seghier, M.L.; Razi, A.; Price, C.J.; Friston, K.J. A guide to group effective connectivity analysis, part 1: First level analysis with DCM for fMRI. NeuroImage 2019, 200, 174–190. [Google Scholar] [CrossRef]
- Zeidman, P.; Jafarian, A.; Seghier, M.L.; Litvak, V.; Cagnan, H.; Price, C.J.; Friston, K.J. A guide to group effective connectivity analysis, part 2: Second level analysis with PEB. NeuroImage 2019, 200, 12–25. [Google Scholar] [CrossRef]
- Johansen-Berg, H.; Behrens, T.E.; Robson, M.D.; Drobnjak, I.; Rushworth, M.F.; Brady, J.M.; Smith, S.M.; Higham, D.J.; Matthews, P.M. Changes in connectivity profiles define functionally distinct regions in human medial frontal cortex. Proc. Natl. Acad. Sci. USA 2004, 101, 13335–13340. [Google Scholar] [CrossRef]
- Matsuzaka, Y.; Aizawa, H.; Tanji, J. A motor area rostral to the supplementary motor area (presupplementary motor area) in the monkey: Neuronal activity during a learned motor task. J. Neurophysiol. 1992, 68, 653–662. [Google Scholar] [CrossRef] [PubMed]
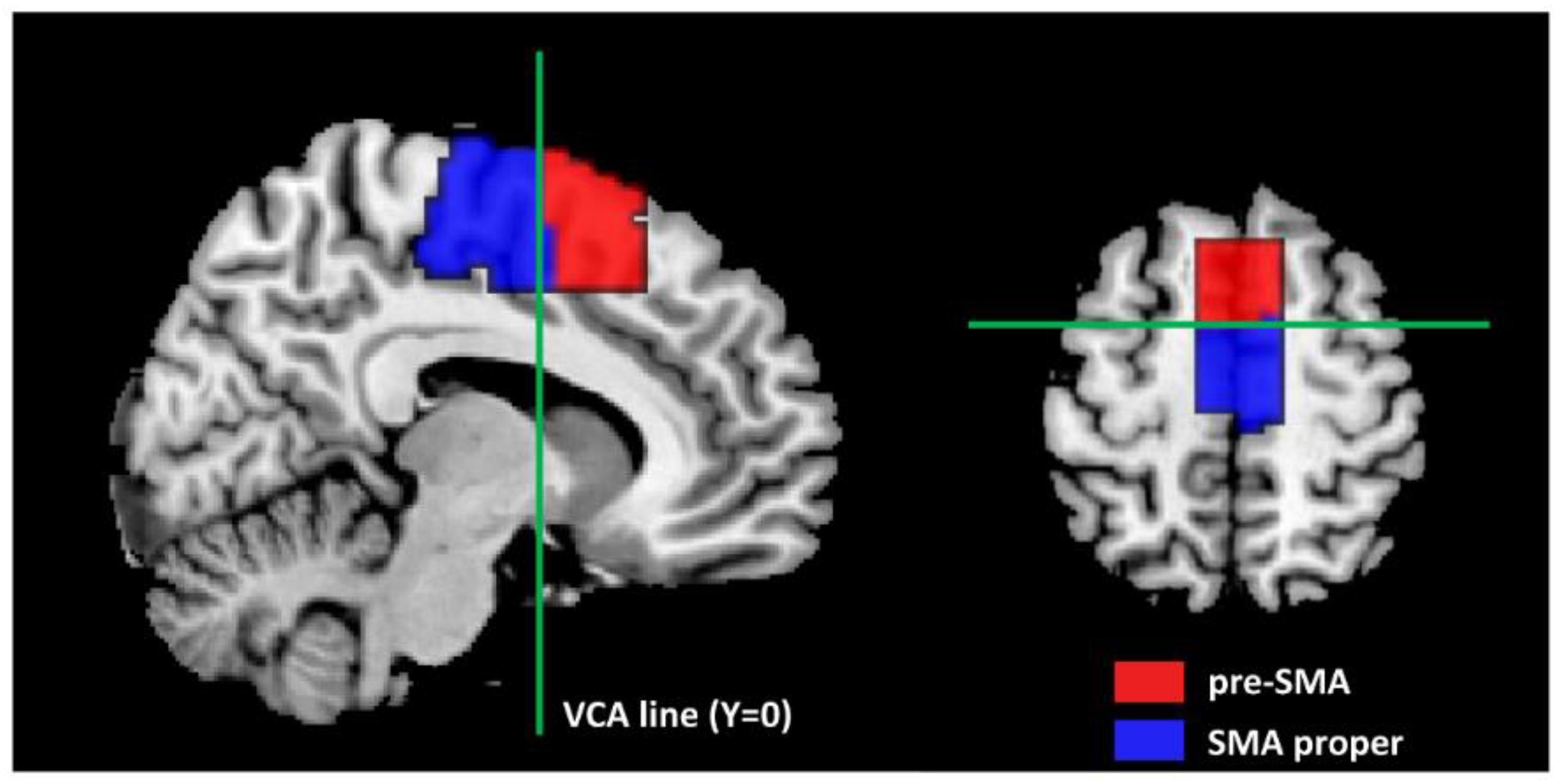
- Kim, J.H.; Lee, J.M.; Jo, H.J.; Kim, S.H.; Lee, J.H.; Kim, S.T.; Seo, S.W.; Cox, R.W.; Na, D.L.; Kim, S.I.; et al. Defining functional SMA and pre-SMA subregions in human MFC using resting state fMRI: Functional connectivity-based parcellation method. NeuroImage 2010, 49, 2375–2386. [Google Scholar] [CrossRef] [PubMed]
- Nachev, P.; Kennard, C.; Husain, M. Functional role of the supplementary and pre-supplementary motor areas. Nat. Rev. Neurosci. 2008, 9, 856–869. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wang, X.; Xia, M.; Liao, X.; Evans, A.; He, Y. GRETNA: A graph theoretical network analysis toolbox for imaging connectomics. Front. Hum. Neurosci. 2015, 9, 386. [Google Scholar]
- Wang, L.; Qiang, L.; Hong, L.; Hu, D. Functional Connectivity-Based Parcellation of Human Medial Frontal Cortex via Maximum Margin Clustering. In Proceedings of the International Conference on Intelligent Science and Intelligent Data Engineering, Nanjing, China, 15–17 October 2012. [Google Scholar]
- Chumbley, J.R.; Friston, K.J. False discovery rate revisited: FDR and topological inference using Gaussian random fields. NeuroImage 2009, 44, 62–70. [Google Scholar] [CrossRef]
- Smith, S.M.; Miller, K.L.; Salimi-Khorshidi, G.; Webster, M.; Beckmann, C.F.; Nichols, T.E.; Ramsey, J.D.; Woolrich, M.W. Network modelling methods for FMRI. NeuroImage 2011, 54, 875–891. [Google Scholar] [CrossRef]
- Burgess, P.W.; Dumontheil, I.; Gilbert, S.J. The gateway hypothesis of rostral prefrontal cortex (area 10) function. Trends Cogn. Sci. 2007, 11, 290–298. [Google Scholar] [CrossRef]
- Koechlin, E.; Hyafil, A. Anterior prefrontal function and the limits of human decision-making. Science 2007, 318, 594–598. [Google Scholar] [CrossRef]
- Friston, K. Learning and inference in the brain. Neural Netw. 2003, 16, 1325–1352. [Google Scholar] [CrossRef]
- Friston, K. A theory of cortical responses. Philos. Trans. R. Soc. Lond. 2005, 360, 815–836. [Google Scholar] [CrossRef]
- Stephan, K.E.; Iglesias, S.; Heinzle, J.; Diaconescu, A.O. Translational Perspectives for Computational Neuroimaging. Neuron 2015, 87, 716–732. [Google Scholar] [CrossRef] [PubMed]
- Rushworth, M.F.; Hadland, K.A.; Paus, T.; Sipila, P.K. Role of the human medial frontal cortex in task switching: A combined fMRI and TMS study. J. Neurophysiol. 2002, 87, 2577–2592. [Google Scholar] [CrossRef]
- Li, C.S.; Huang, C.; Constable, R.T.; Sinha, R. Imaging response inhibition in a stop-signal task: Neural correlates independent of signal monitoring and post-response processing. J. Neurosci. Off. J. Soc. Neurosci. 2006, 26, 186–192. [Google Scholar] [CrossRef]
- Hikosaka, O.; Sakai, K.; Miyauchi, S.; Takino, R.; Sasaki, Y.; Putz, B. Activation of human presupplementary motor area in learning of sequential procedures: A functional MRI study. J. Neurophysiol. 1996, 76, 617–621. [Google Scholar] [CrossRef] [PubMed]
- Lau, H.C.; Rogers, R.D.; Ramnani, N.; Passingham, R.E. Willed action and attention to the selection of action. NeuroImage 2004, 21, 1407–1415. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Daunizeau, J.; Stephan, K.E.; Penny, W.; Hu, D.; Friston, K. Generalised filtering and stochastic DCM for fMRI. NeuroImage 2011, 58, 442–457. [Google Scholar] [CrossRef] [PubMed]
- Wei, H.; Jafarian, A.; Zeidman, P.; Litvak, V.; Friston, K.J. Bayesian fusion and multimodal DCM for EEG and fMRI. NeuroImage 2020, 211, 116595. [Google Scholar] [CrossRef]
- Goncalves, M.S.; Hall, D.A. Connectivity analysis with structural equation modelling: An example of the effects of voxel selection. NeuroImage 2003, 20, 1455–1467. [Google Scholar] [CrossRef]
- Behzadi, Y.; Restom, K.; Liau, J.; Liu, T.T. A component based noise correction method (CompCor) for BOLD and perfusion based fMRI. NeuroImage 2007, 37, 90–101. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Drivers | Controls | p Value |
|---|---|---|---|
| Number of subjects | 20 | 20 | |
| Age (years) | 39.5 ± 5.8 | 41.1 ± 5.0 | 0.34 a |
| Sex (male/female) | 20/0 | 18/2 | 0.15 b |
| Education (years) | 9.5 ± 1.8 | 9.0 ± 1.4 | 0.37 a |
| Duration of taxi driving (years) | 4.9 ± 3.5 | ||
| Duration of total driving (years) | 11.7 ± 4.9 |
| Anatomical Area | Size | MNI Coordinates | T Value | p Value (FDR Corrected) | Brodmann’s Area | ||
|---|---|---|---|---|---|---|---|
| x | y | z | |||||
| Pre-SMA | |||||||
| Driver > control | |||||||
| Left orbital frontal cortex | 113 | −21 | 57 | −9 | 5.62 | 0.003 | 10, 11 |
| Left superior frontal gyrus | 63 | −18 | 60 | 21 | 4.52 | 0.011 | 9, 10 |
| Left middle temporal gyrus | 51 | −60 | −21 | −9 | 4.43 | 0.011 | 21 |
| Driver < control | |||||||
| Right middle frontal gyrus | 27 | 39 | 6 | 39 | 4.47 | 0.189 | 6, 9 |
| Right inferior parietal lobule | 33 | 51 | −30 | 36 | 4.15 | 0.138 | 2, 40 |
| SMA proper | |||||||
| No significance | |||||||
| Whole SMA | |||||||
| Driver > control | |||||||
| Left superior frontal gyrus | 29 | −21 | 57 | 21 | 4.12 | 0.497 | 10 |
| Effective Connectivity | Sensitivity (%) | Specificity (%) | Accuracy (%) | Correlation | Significance (p Value) |
|---|---|---|---|---|---|
| Pre-SMA→pre-SMA | 80 | 60 | 70 | 0.3352 | 0.0172 |
| Pre-SMA→lOFC | 65 | 55 | 60 | 0.0977 | 0.2744 |
| Pre-SMA→lSFG | 80 | 65 | 72.5 | 0.4057 | 0.0047 |
| lOFC→lSFG | 70 | 55 | 62.5 | 0.0556 | 0.3666 |
| lSFG→lSFG | 60 | 55 | 57.5 | −0.2116 | 0.9050 |
| lMTG→lOFC | 55 | 50 | 52.5 | 0.1609 | 0.1606 |
| lMTG→lMTG | 80 | 45 | 62.5 | −0.0222 | 0.5542 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wei, H.; Wang, L.; Peng, L.; Li, C.; Ma, T.; Hu, D. Effective Connectivity Changes among Brain Hierarchical Architecture of Pre-Supplementary Motor Area in Taxi Drivers. Appl. Sci. 2023, 13, 11471. https://doi.org/10.3390/app132011471
Wei H, Wang L, Peng L, Li C, Ma T, Hu D. Effective Connectivity Changes among Brain Hierarchical Architecture of Pre-Supplementary Motor Area in Taxi Drivers. Applied Sciences. 2023; 13(20):11471. https://doi.org/10.3390/app132011471
Chicago/Turabian StyleWei, Huilin, Lubin Wang, Limin Peng, Chenming Li, Tian Ma, and Dewen Hu. 2023. "Effective Connectivity Changes among Brain Hierarchical Architecture of Pre-Supplementary Motor Area in Taxi Drivers" Applied Sciences 13, no. 20: 11471. https://doi.org/10.3390/app132011471