
Benincasa hispida Extract Prevents Ovariectomy-Induced Osteoporosis in Female ICR Mice

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Sample Preparation
2.2. Animal and Experimental Design
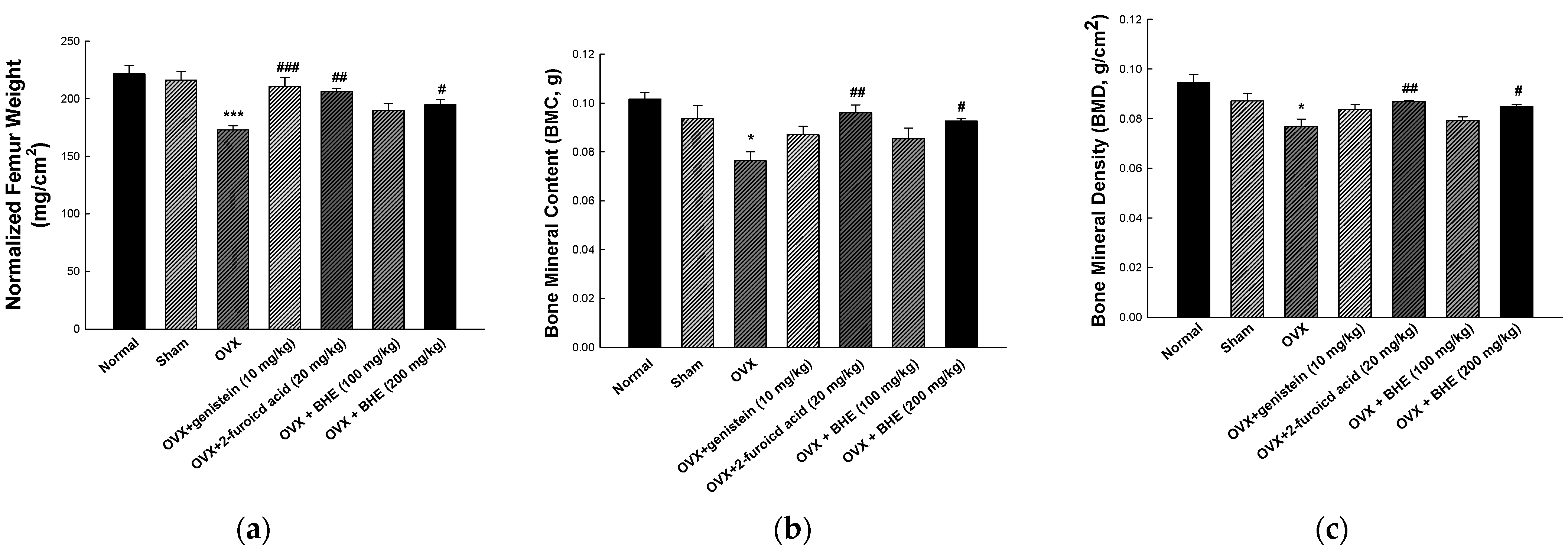
2.3. Assessment of Bone Mineral Density
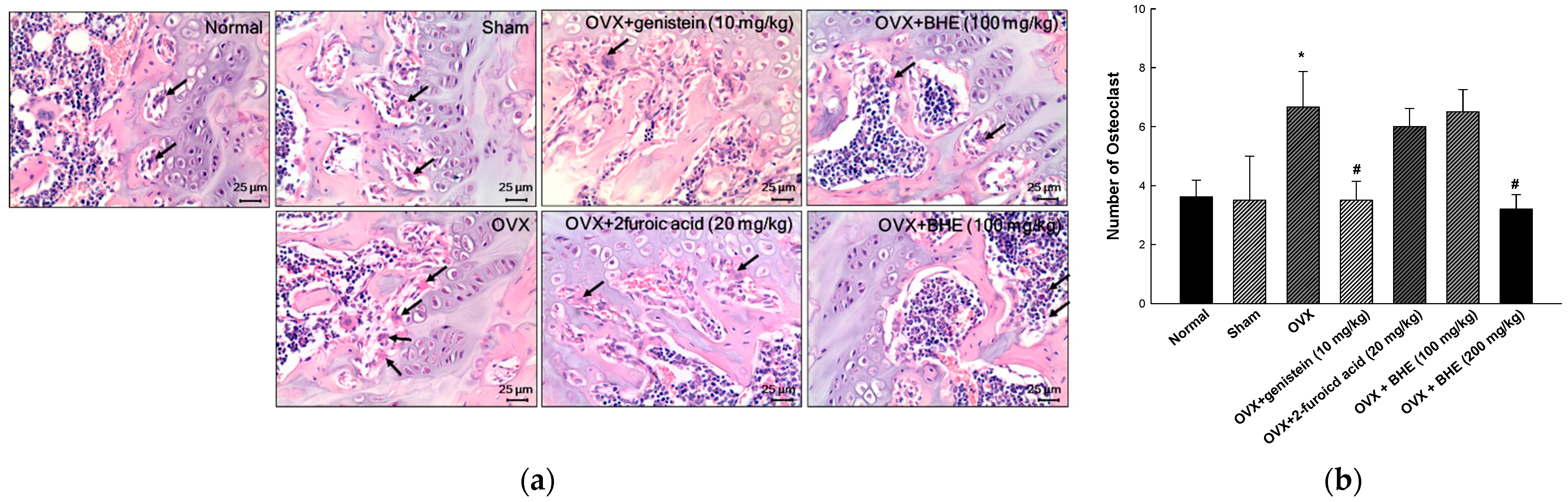
2.4. Histological Assessment of Osteoclast
2.5. Assessment of Serum Biochemical Parameter
2.6. Statistical Analysis
3. Results
3.1. Body Weight and Food Intake
3.2. Effect of BHE and 2-Furoic Acid on Bone Morphology
3.3. Effect of BHE and 2-Furoic Acid on Osteoclast Formation
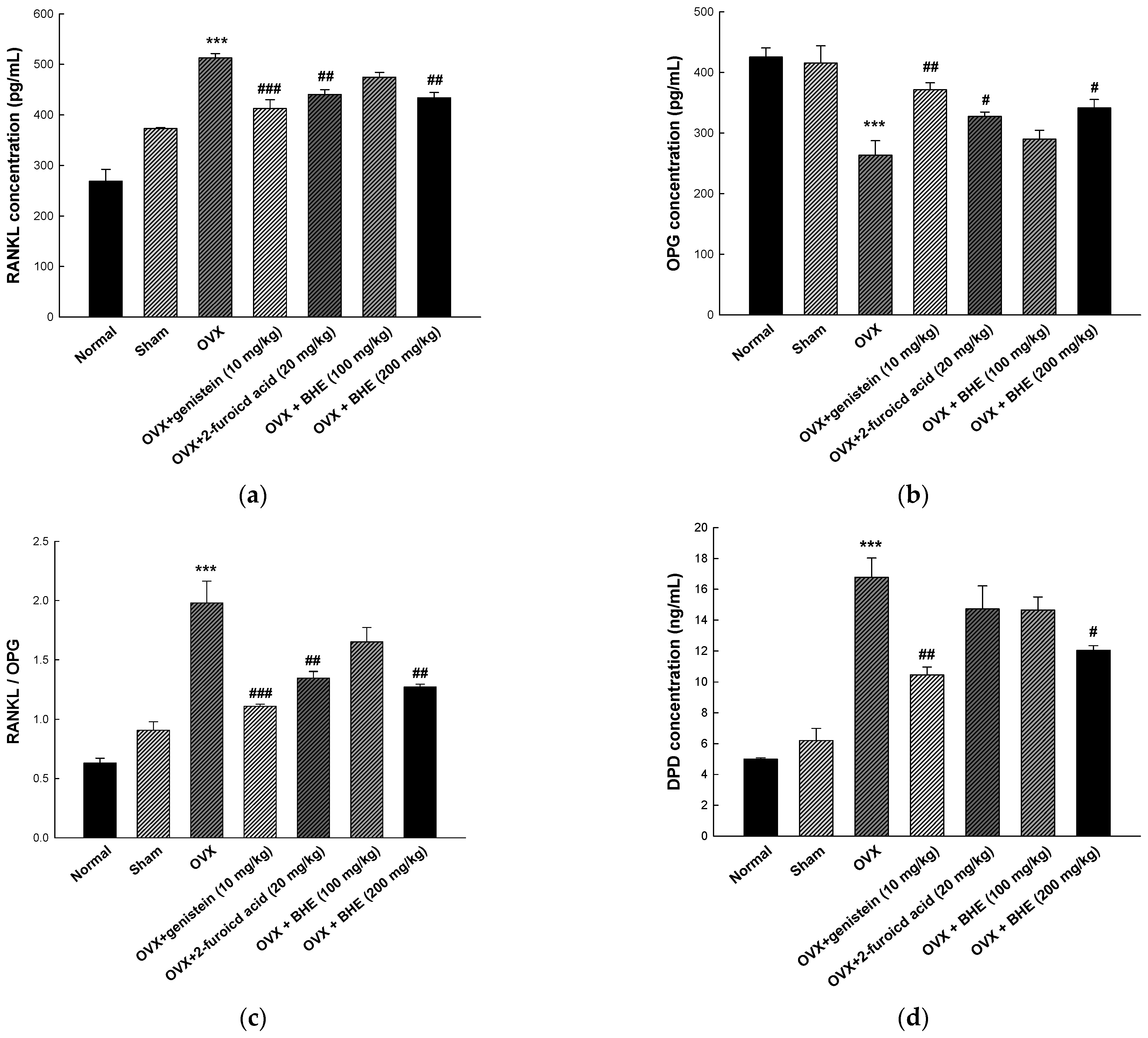
3.4. Effect of BHE and 2-Furoic Acid on Bone Resorption Markers
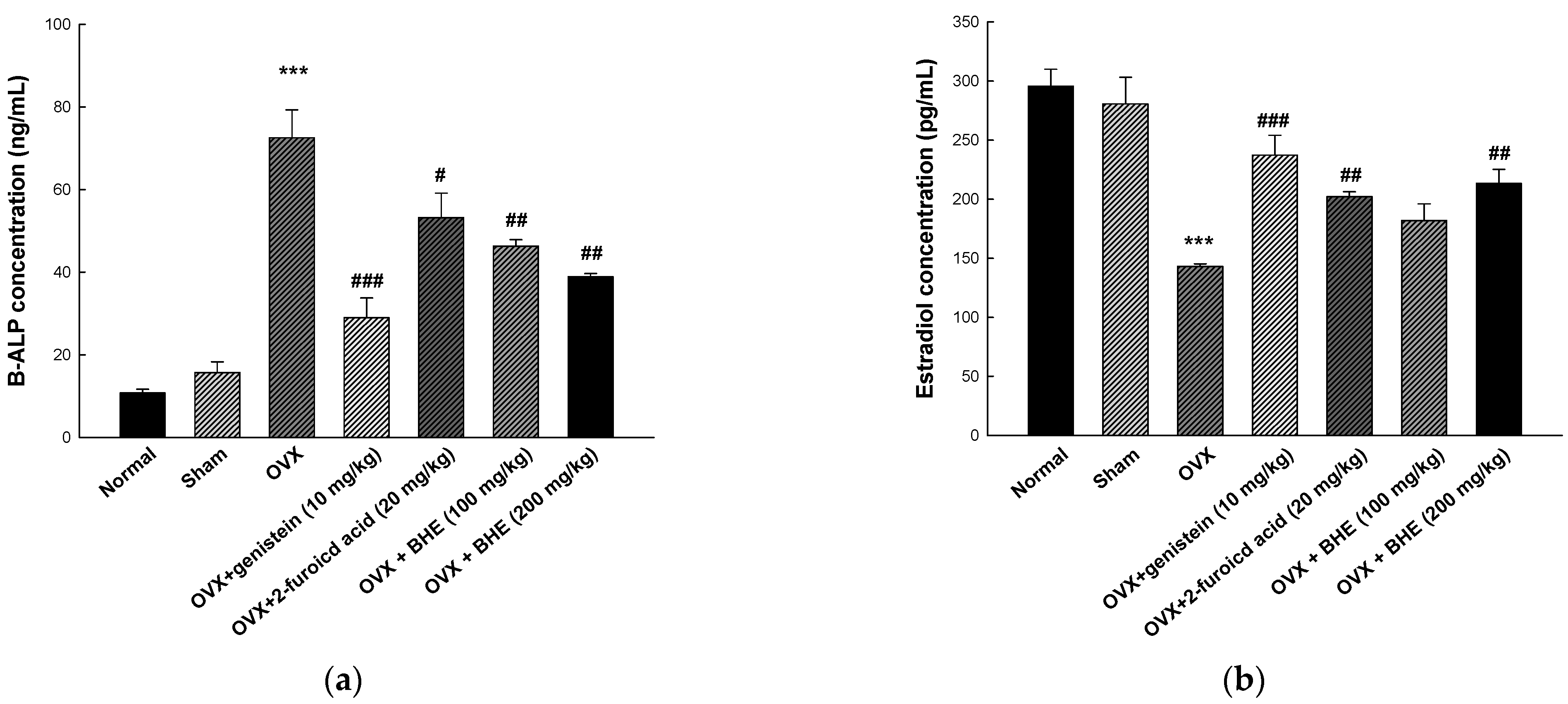
3.5. Effect of BHE and 2-Furoic Acid on Bone Formation Markers
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rudnicka, E.; Napierała, P.; Podfigurna, A.; Męczekalski, B.; Smolarczyk, R.; Grymowicz, M. The World Health Organization (WHO) approach to healthy ageing. Maturitas 2020, 139, 6–11. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Hou, X.; Zhang, J.; Li, J.; Wu, P.; Yan, L.; Qian, H. Diagnostic and Therapeutic Roles of Extracellular Vesicles in Aging-Related Diseases. Oxidative Med. Cell. Longev. 2022, 2022, 6742792. [Google Scholar] [CrossRef] [PubMed]
- Greco, E.A.; Pietschmann, P.; Migliaccio, S. Osteoporosis and sarcopenia increase frailty syndrome in the elderly. Front. Endocrinol. 2019, 10, 255. [Google Scholar] [CrossRef] [PubMed]
- Le, B.Q.; Nurcombe, V.; Cool, S.M.; Van Blitterswijk, C.A.; De Boer, J.; LaPointe, V.L.S. The components of bone and what they can teach us about regeneration. Materials 2017, 11, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saad, F.A. Novel insights into the complex architecture of osteoporosis molecular genetics. Ann. N. Y. Acad. Sci. 2019, 1462, 37–52. [Google Scholar] [CrossRef]
- Aspray, T.J.; Hill, T.R. Osteoporosis and the ageing skeleton. Subcell. Biochem. 2019, 91, 453–476. [Google Scholar]
- Johnston, C.B.; Dagar, M. Osteoporosis in older adults. Med. Clin. N. Am. 2020, 104, 873–884. [Google Scholar] [CrossRef]
- Hobson, E.E.; Ralsteo, S.H. Role of genetic factors in the pathophysiology and management of osteoporosis. Clin. Endocrinol. 2001, 54, 1–9. [Google Scholar] [CrossRef]
- Garnero, P.; Delmas, P.D. New developments in biochemical markers for osteoporosis. Calcif. Tissue Int. 1996, 1, S2–S9. [Google Scholar] [CrossRef]
- Johansen, J.S.; Riis, B.J.; Delmas, P.D.; Christiansen, C. Plasma BGP: An indicator of spontaneous bone loss and of the effect of oestrogen treatment in postmenopausal women. Eur. J. Clin. Investig. 1988, 18, 191–195. [Google Scholar] [CrossRef]
- Cauley, J.A.; Gutai, J.P.; Sandier, R.B.; LaPorte, A.E.; Kuller, L.H.; Sashin, D. The relationship of endogenous estrogen to bone density and bone area in normal poetmenopausal women. Am. J. Epldemiol. 1986, 124, 752–761. [Google Scholar] [CrossRef]
- Ohta, H.; Masuzawa, T.; Ikeda, T.; Suda, Y.; Makita, K.; Nozawa, S. Which is more osteoporosis inducing, menopause or oophorectomy? Bone Miner. 1992, 19, 273–285. [Google Scholar] [CrossRef]
- Li, M.; Shen, Y.; Wronski, T.J. Time course of femoral neck osteopenia in ovariectomized rats. Bone 1997, 20, 55–61. [Google Scholar] [CrossRef]
- Valero, M.A.; Loinaz, C.; Larrodera, L.; Leon, M.; Moreno, E.; Hawkins, F. Calcitonin and bisphosphonates treatment in bone loss after liver transplantation. Calcif. Tissue Int. 1995, 57, 15–19. [Google Scholar] [CrossRef]
- Ross, R.K.; Paganini-Hill, A.; Wan, P.C.; Pike, M.C. Effect of hormone replacement therapy on breast cancer risk: Estrogen versus estrogen plus progestin. J. Natl. Cancer Inst. 2000, 92, 328–332. [Google Scholar] [CrossRef]
- Ziel, H.K. Estrogen’s role in endometrial cancer. Obstet. Gynecol. 1982, 60, 509–515. [Google Scholar]
- Nelson, H.D.; Humphrey, L.L.; Nygren, P.; Teutsch, S.M.; Allan, J.D. Postmenopausal hormone replacement therapy: Scientific review. J. Am. Med. Assoc. 2002, 288, 872–881. [Google Scholar] [CrossRef]
- Mohammad, N.A.; Anwar, F.; Mehmood, T.; Hamid, A.A.; Muhammad, K.; Saari, N. Phenolic compounds, tocochromanols profile and antioxidant properties of winter melon [Benincasa hispida (Thunb.) Cogn.] seed oils. J. Food Meas. Charact. 2019, 13, 940–948. [Google Scholar] [CrossRef]
- Park, G.R.; Lee, J.A. Anti-oxidant, anti-inflammatory and Whitening effect of Benincasa hispida seed extract. J. Converg. Inf. Technol. 2020, 10, 249–256. [Google Scholar]
- Lim, S.J.; Jeong, J.G.; Kim, M.W.; Choi, S.S.; Han, H.K.; Park, J.E. Effects of Benincasa hispida intake on blood glucose and lipid level in streptozotocin induced diabetic rats. Korean J. Nutr. 2003, 36, 335–343. [Google Scholar]
- You, Y.H.; Jun, W.J. Effects of fractions from Benincasa hispida on inhibition of adipogenesis in 3T3-L1 preadipocytes. J. Korean Soc. Food Sci. Nutr. 2012, 41, 895–900. [Google Scholar] [CrossRef]
- Grover, J.K.; Adiga, G.; Vats, V.; Rathi, S.S. Extracts of Benincasa hispida prevent development of experimental ulcers. J. Ethnopharmacol. 2001, 78, 159–164. [Google Scholar] [CrossRef]
- Kim, M.W. Effects of benincasa hispida seed supplementation on glyeogen status and lipid peroxidatin in streptozotecin-induced diabetic rats. J. Nutr. Health 2004, 37, 865–871. [Google Scholar]
- Lim, T.K. Edible medicinal and non-medicinal plants. In Modified Stems, Roots, Bulbs; Springer: Berlin/Heidelberg, Germany, 2012; Volume 11, pp. 285–292. [Google Scholar]
- Gill, N.S.; Dhiman, K.; Bajwa, J.; Sharma, P.; Sood, S. Evaluation of free radical scavenging, anti-inflammatory and analgesic potential of Benincasa hispida seed extract. Int. J. Pharmacol. 2010, 6, 652–657. [Google Scholar] [CrossRef] [Green Version]
- Cho, B.Y.; Park, M.R.; Lee, J.H.; Ra, M.J.; Han, K.C.; Kang, I.J.; Lee, O.H. Standardized Cirsium setidens Nakai ethanolic extract suppresses adipogenesis and regulates lipid metabolisms in 3T3-L1 adipocytes and C57BL/6J mice fed high-fat diets. J. Med. Food 2017, 20, 763–776. [Google Scholar] [CrossRef]
- Jeon, S.Y.; Jeong, E.J.; Baek, J.H.; Cha, Y.J. Analytical method validation of quercetin in Changnyeong onion extract as a functional ingredient for functional health food. J. Korean Soc. Food Sci. Nutr. 2011, 40, 565–569. [Google Scholar] [CrossRef]
- Xie, J.; Zhang, A.-H.; Sun, H.; Yan, G.-L.; Wang, X.-J. Recent advances and effective strategies in the discovery and applications of natural products. RSC Adv. 2018, 8, 812–824. [Google Scholar] [CrossRef] [Green Version]
- Hall, I.H.; Williams, W.L.; Rhyne, K.A.; Knowles, M. The hypolipidemic activity of furoic acid and furylacrylic acid derivatives in rodents. Pharm. Res. 1985, 2, 233–238. [Google Scholar] [CrossRef]
- Hall, I.H.; Wong, O.T.; Reynolds, D.J.; Chang, J.J. Hypolipidemic effects of 2-uroic acid in sprague-dawley rats. Arch. Pharm. 1993, 326, 15–23. [Google Scholar] [CrossRef]
- Sajadi, Z.; Abrishami, M.M.; Chapman, J.M.; Hall, I.H. Synthesis and evaluation of the antitumor properties of esters of 2-furoic acid and 2-furylacrylic acid. J. Pharm. Sci. 1984, 73, 266–267. [Google Scholar] [CrossRef]
- Choi, S.I.; Han, X.; Men, X.; Lee, S.J.; Park, M.H.; Lee, O.H.; Yang, J.M.; Choi, Y.E.; Cho, J.H. Development and validation of an analytical method for 2-furoic acid in Benincasa hispida extracts (HR1901-W). J. Agri. Life Environ. Sci. 2021, 33, 311–320. [Google Scholar]
- Choi, Y.E.; Yang, J.M.; Cho, J.H. Benincasa hispida extract promotes proliferation, differentiation, and mineralization of MC3T3-E1 preosteoblasts and inhibits the differentiation of RAW 246.7 osteoclast precursors. Appl. Sci. 2022, 12, 8849. [Google Scholar] [CrossRef]
- Behloul, N.; Wu, G. Genistein: A promising therapeutic agent for obesity and diabetes treatment. Eur. J. Pharmacol. 2013, 698, 31–38. [Google Scholar] [CrossRef]
- Yang, L.; Yu, Z.; Qu, H.; Li, M. Comparative effects of hispidulin, genistein, and icariin with estrogen on bone tissue in ovariectomized rats. Cell Biochem. Biophys. 2014, 70, 485–490. [Google Scholar] [CrossRef]
- Udagawa, N.; Koide, M.; Nakamura, M.; Nakamichi, Y.; Yamashita, T.; Uehara, S.; Kobayashi, Y.; Furuya, Y.; Tasuda, H.; Fukuda, C.; et al. Osteoclast differentiation by RANKL and OPG signaling pathways. J. Bone Miner. Metab. 2021, 39, 19–26. [Google Scholar] [CrossRef]
- Seibel, M.J.; Cosman, F.; Shen, V.; Gordon, S.; Dempster, D.W.; Ratcliffe, A.; Lindsay, R. Urinary hydroxypyridinium crosslinks of collagen as markers of bone resorption and estrogen efficacy in postmenopausal osteoporosis. J. Bone Miner. Res. 1993, 8, 881–889. [Google Scholar] [CrossRef]
- Lane, N.E. Epidemiology, etiology, and diagnosis of osteoporosis. Am. J. Obstet. Gynecol. 2006, 194, S3–S11. [Google Scholar] [CrossRef]
- Campbell, S.E.; Febbraio, M.A. Effects of ovarian hormones on exercise metabolism. Curr. Opin. Clin. Nutr. Metab. Care 2001, 4, 515–520. [Google Scholar] [CrossRef]
- Misso, M.L.; Jang, C.; Adams, J.; Tran, J.; Murata, Y.; Bell, R.; Boon, W.C.; Simpson, E.R.; Davis, S.R. Differential expression of factors involved in fat metabolism with age and the menopause transition. Maturitas 2005, 51, 299–306. [Google Scholar] [CrossRef]
- Zoth, N.; Weigt, C.; Laudenbach-Leschowski, U.; Diel, P. Physical activity and estrogen treatment reduce visceral body fat and serum levels of leptin in an additive manner in a diet induced animal model of obesity. J. Steroid Biochem. Mol. Biol. 2010, 122, 100–105. [Google Scholar] [CrossRef]
- Zheng, W.; Rogoschin, J.; Niehoff, A.; Oden, K.; Kulling, S.E.; Xie, M.; Diel, P. Combinatory effects of phytoestrogens and exercise on body fat mass and lipid metabolism in ovariectomized female rats. J. Steroid Biochem. Mol. Biol. 2018, 178, 73–81. [Google Scholar] [CrossRef]
- Shen, H.H.; Huang, S.Y.; Kung, C.W.; Chen, S.Y.; Chen, Y.F.; Cheng, P.Y.; Lam, K.K.; Lee, Y.M. Genistein ameliorated obesity accompanied with adipose tissue browning and attenuation of hepatic lipogenesis in ovariectomized rats with high-fat diet. J. Nutr. Biochem. 2019, 67, 111–122. [Google Scholar] [CrossRef]
- Kim, H.K.; Nelson-Dooley, C.; Della-Fera, M.A.; Yang, J.Y.; Zhang, W.; Duan, J.; Hartzel, D.L.; Hamrick, M.W.; Baile, C.A. Genistein decreases food intake, body weight, and fat pad weight and causes adipose tissue apoptosis in ovariectomized female mice. J. Nutr. 2006, 136, 409–414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishimi, Y.; Arai, N.; Wang, X.; Wu, J.; Umegaki, K.; Miyaura, C.; Takeda, A.; Ikegami, S. Difference in effective dosage of genistein on bone and uterus in ovariectomized mice. Biochem. Biophys. Res. Commun. 2000, 274, 697–701. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.M.; Lee, H.S.; Jung, J.I.; Lim, S.M.; Lim, J.H.; Ha, W.H.; Jeon, C.L.; Lee, J.Y.; Kim, E.J. Effect of isoflavone-enriched whole soy milk powder supplementation on bone metabolism in ovariectomized mice. Nutr. Res. Pract. 2018, 12, 275–282. [Google Scholar] [CrossRef] [PubMed]
- Rodan, G.A.; Martin, T.J. Therapeutic approaches to bone diseases. Science 2000, 289, 1508–1514. [Google Scholar] [CrossRef]
- Marie, P.J.; Hott, M.; Modrowski, D.; De Pollak, C.; Guillemain, J.; Deloffre, P.; Tsouderos, Y. An uncoupling agent containing strontium prevents bone loss by depressing bone resorption and maintaining bone formation in estrogen-deficient rats. J. Bone Miner. Res. 1993, 8, 607–615. [Google Scholar] [CrossRef]
- Bowman, B.M.; Miller, S.C. Elevated progesterone during pseudopregnancy may prevent bone loss associated with low estrogen. J. Bone Miner. Res. 1996, 11, 15–21. [Google Scholar] [CrossRef]
- Khosla, S.; Oursler, M.J.; Monroe, D.G. Estrogen and the skeleton. Trends Endocrinol. Metab. 2012, 23, 576–581. [Google Scholar] [CrossRef] [Green Version]
- Clarke, B.L.; Khosla, S. Female reproductive system and bone. Arch. Biochem. Biophys. 2010, 503, 118–128. [Google Scholar] [CrossRef] [Green Version]
- Hou, J.M.; Xue, Y.; Lin, Q.M. Bovine lactoferrin improves bone mass and microstructure in ovariectomized rats via OPG/RANKL/RANK pathway. Acta Pharmacol. Sin. 2012, 33, 1277–1284. [Google Scholar] [CrossRef] [Green Version]
- Rissanen, J.P.; Suominen, M.I.; Peng, Z.; Halleen, J.M. Secreted tartrate-resistant acid phosphatase 5b is a Marker of osteoclast number in human osteoclast cultures and the rat ovariectomy model. Calcif. Tissue Int. 2008, 82, 108–115. [Google Scholar] [CrossRef]
- Mulari, M.; Vääräniemi, J.; Väänänen, H.K. Intracellular membrane trafficking in bone resorbing osteoclasts. Microsc. Res. Tech. 2003, 61, 496–503. [Google Scholar] [CrossRef]
- Noh, D.; Lim, Y.; Lee, H.; Kim, H.; Kwon, O. Soybean-hop alleviates estrogen deficiency-related bone loss and metabolic dysfunction in ovariectomized rats fed a high-fat diet. Molecules 2018, 23, 1205. [Google Scholar] [CrossRef] [Green Version]
- Christenson, R.H. Biochemical markers of bone metabolism: An overview. Clin. Biochem. 1997, 30, 573–593. [Google Scholar] [CrossRef]
- Qadrie, Z.L.; Tayebhawisan, N.; Alikhan, M.W.; Samuel, M.; Anandan, R. Antinociceptive and anti-pyretic activity of Benincasa hispida (Thunb) Cogn. in Wistar albino rats. Pak. J. Pharm. Sci. 2009, 22, 287–290. [Google Scholar]
- Jayasree, T.; Kishore, K.K.; Vinay, M.; Vasavi, P.; Dixit, R.; Rajanikanth, M.; Manohar, V.S. Diuretic effect of the chloroform extract of the Benincasa hispida rind (Pericarp) extract in Sprague-Dawley rats. Int. J. Appl. Biol. Pharm. 2011, 2, 94–99. [Google Scholar]
- Shakya, A.; Chaudhary, S.K.; Bhat, H.R.; Ghosh, S.K. Acute and sub-chronic toxicity studies of Benincasa hispida (Thunb.) cogniaux fruit extract in rodents. Regul. Toxicol. Pharmacol. 2020, 118, 104785. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Normal | Sham | OVX | OVX + GEN 10 mg/kg | OVX + 2-FA 20 mg/kg | OVX + BHE 100 mg/kg | OVX + BHE 200 mg/kg | |
|---|---|---|---|---|---|---|---|---|
| Variables | ||||||||
| Initial body weight (g) | 30.42 ± 0.67 | 29.43 ± 1.71 | 27.58 ± 1.40 | 28.80 ± 0.96 | 28.72 ± 0.16 | 28.68 ± 0.52 | 29.22 ± 0.15 | |
| Final body weight (g) | 40.08 ± 1.53 | 36.70 ± 1.91 | 37.38 ± 3.70 | 41.24 ± 2.56 | 42.28 ± 2.44 | 40.10 ± 2.73 | 40.77 ± 1.16 | |
| Weight gain (g) | 9.66 ± 1.53 | 7.27 ± 0.94 | 9.80 ± 2.62 | 12.44 ± 1.89 | 13.56 ± 2.41 | 11.43 ± 2.27 | 11.55 ± 1.13 | |
| FER (%) | 2.35 ± 0.37 | 2.11 ± 0.27 | 2.77 ± 0.74 | 3.50 ± 0.53 | 3.40 ± 0.60 | 3.36 ± 0.67 | 3.15 ± 0.31 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choi, S.-I.; Han, X.; Men, X.; Lee, S.-J.; Oh, G.; Choi, Y.-E.; Yang, J.-M.; Cho, J.-H.; Lee, O.-H. Benincasa hispida Extract Prevents Ovariectomy-Induced Osteoporosis in Female ICR Mice. Appl. Sci. 2023, 13, 832. https://doi.org/10.3390/app13020832
Choi S-I, Han X, Men X, Lee S-J, Oh G, Choi Y-E, Yang J-M, Cho J-H, Lee O-H. Benincasa hispida Extract Prevents Ovariectomy-Induced Osteoporosis in Female ICR Mice. Applied Sciences. 2023; 13(2):832. https://doi.org/10.3390/app13020832
Chicago/Turabian StyleChoi, Sun-Il, Xionggao Han, Xiao Men, Se-Jeong Lee, Geon Oh, Ye-Eun Choi, Jung-Mo Yang, Ju-Hyun Cho, and Ok-Hwan Lee. 2023. "Benincasa hispida Extract Prevents Ovariectomy-Induced Osteoporosis in Female ICR Mice" Applied Sciences 13, no. 2: 832. https://doi.org/10.3390/app13020832