1. Introduction
Powered exoskeletons are becoming widely spread and reliable tools in the field of neurorehabilitation for spinal cord injuries, with multiple studies highlighting the beneficial neural plastic effects potentiated through their use. Previous studies have demonstrated that neurorehabilitation protocols combining motor-imagery-based brain–machine interfaces (BMIs) for exoskeleton control have led to significant neurological improvements [
1,
2,
3]. Specifically, the combination of tactile feedback with motor imagery is becoming more common, thereby emphasizing the relevance of tactile processing for neurorehabilitation.
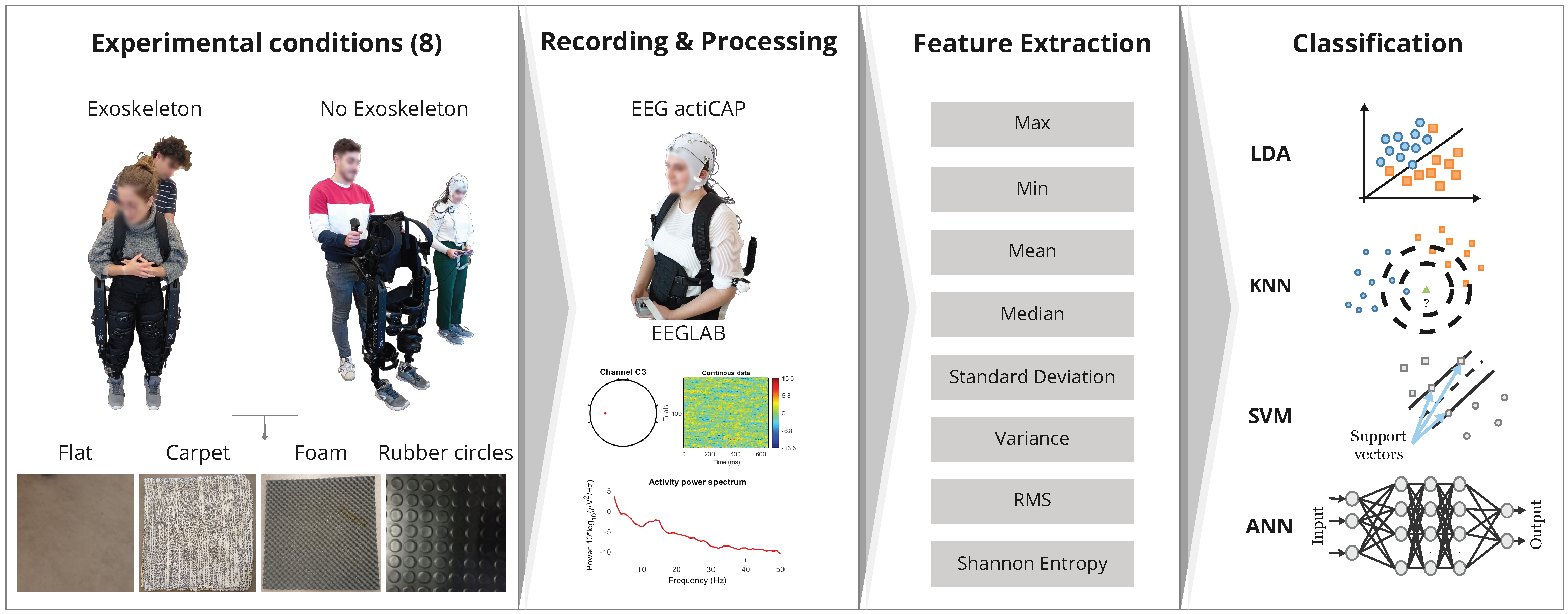
Currently, the utilization of an exoskeleton (refer to
Figure 1) is often associated with the use of bulky equipment, which often necessitates close contact between the exoskeleton and the soles of the feet. This contact is achieved through artificial soles that provide support to the lower limbs. However, the presence of an additional component between the foot sole and the ground has the potential to interfere with the processing of the somatosensory information related to textures. Therefore, BMIs that integrate exoskeletons and somatosensory processing can offer numerous advantages, but the use of exoskeletons can have two potential effects: (i) they may reduce the number of somatosensory signals transmitted to the spinal cord, and (ii) they may introduce additional changes to the dynamics of somatosensory processing. Both of these effects have the potential to alter the performance of BMIs. Considering the possibility of diminished differences in somatosensory neural activity due to the aforementioned factors, it is important to evaluate the following: (a) the general patterns of neural activity associated with exoskeleton use and (b) the performance of common classifiers decoding the neural activity of individuals controlling an exoskeleton when encountering different textured pavements.
Neural correlates of tactile sensory processing have been extensively studied using electroencephalography [
4,
5,
6,
7], but there has been a growing interest in complementing the study of neural signals with the use of classifiers [
8,
9,
10,
11,
12,
13,
14,
15]. Some of the most popular classifiers for neural decoding include the linear support vector machine (L-SVM), the K-nearest neighbor algorithm (KNN), linear discriminant analysis (LDA), and the artificial neural network (ANN) [
12,
13,
14].
The SVM creates a separation hyperplane to separate the two different input classes into the appropriate category. The data points nearest to the hyperplane are designated by support vectors, and the classification process is based on the mapping of the new input data point. This model evaluates on which side of the hyperplane the new value is located, which it then attributes to the corresponding class according to nearest support vector. This algorithm possesses lower computational complexity when compared with other supervised algorithms, such as the ANN and KNN [
15,
16,
17].
The KNN is a nonparametric supervised algorithm that classifies different objects based on their K-closest neighbors. Therefore, for a new data point, this model determines the class by calculating the majority of the neighbours class labels. The letter “K” identifies the number of chosen neighbours, and, consequently, the selected value plays an important part in the model’s performance [
15,
16,
17].
LDA is classified as a linear technique that uses, as with the SVM model, a hyperplane to separate the different classes and to classify the input data values. The main goal of this model is to reduce the input value space while maximizing the separation between classes in the data. This classifier has a low computational cost [
15,
16,
17].
ANN models are inspired by the functioning of the human brain, more specifically, by how the neurons communicate between them. The basic unit of the ANN is the artificial neuron, which is also designated as the perceptron (neural network unit). The basic functioning of these units consists of input values that are multiplied by a specific weight. Afterwards, these values are summed, and this weighted sum is then passed to an activation function that controls the output, since it is responsible for concluding if the weighted sum value is smaller or bigger than the threshold value [
15,
16,
17].
Several other types of classifiers based in tensor methods, deep learning, and Riemannian geometry are now gaining importance due to their ability to be used with smaller training data sets, their higher signal-to-noise ratios, and their capability for transfer learning [
14,
15].
In this study, we conducted a preliminary investigation to address the following questions: (1) whether there were clearly discernible patterns of neural activity recorded from electrodes at the C3 and C4 positions when subjects wore an exoskeleton, (2) whether neural activity exhibited variations when subjects walked on different textured surfaces, and (3) how four commonly used classifiers (SVM, KNN, LDA, and ANN) performed in the classification of exoskeleton use (i.e., Exoskeleton/No Exoskeleton) on four distinct types of pavements (flat surface, carpet, foam, and rubber circles).
We formulated the following hypotheses for this study: Hypothesis (1) proposed that the use of an exoskeleton would lead to changes in the power of specific frequency bands, as it has been demonstrated that exoskeleton control can interfere with other brain functions [
18,
19]. Hypothesis (2) proposed that stepping on less stable textures, which require greater stability during walking, would result in an increase in the central (C3, C4) alpha and theta frequency bands [
20]. These frequency bands are typically associated with tasks involving the vestibular system. Lastly, Hypothesis (3) proposed that all four classifiers would demonstrate the ability to classify the eight-class problem (with/without Exoskeleton for all four pavements) above a chance level based solely on neural activity.
To gather preliminary evidence for these hypotheses, we conducted experiments that involved five subjects. The subjects performed walking tasks with and without an exoskeleton on four different textured pavements: flat (representing regular ground), carpet, foam, and rubber circles (refer to
Figure 1). During the experiments, the subjects’ neural activity was recorded using an electroencephalogram (EEG). When the subjects were not using the exoskeleton, a member of the research team (referred to as exoskeleton operator) replicated the subjects’ movements with the exoskeleton. This was done by holding the exoskeleton near the subject, ensuring that the frequency and sounds associated with the exoskeleton’s movements remained similar between the measurements with and without the exoskeleton. (see
Figure 1; also see Materials and Methodsbelow).
The developed work provides information on the neurophysiological basis of tactile processing during exoskeleton control. Additionally, this study also offers relevant data for the development of tactile-sensation-based BMI systems that aim to provide sensory feedback to their users.
The document is organized into five different chapters. The current
Section 1 intends to describe the significance of this work and its relevance to the scientific community.
Section 2 describes the methods used in this work in order to acquire, process, and classify the biosignal. In the following
Section 3, the obtained results are accessed and
Section 4 and the results are discussed while taking into account the main goal of this work. The
Section 5 presents the conclusions that can be drawn from the developed study.
3. Results
The results section is divided in two parts: (i) the pilot analysis of the power in the most common frequency bands (delta, theta, alpha, beta, and gamma) under different experimental conditions and (ii) the analysis of neural signals in different experimental conditions using four different classifiers (LDA, SVM, ANN, and KNN).
3.1. Pilot Analysis
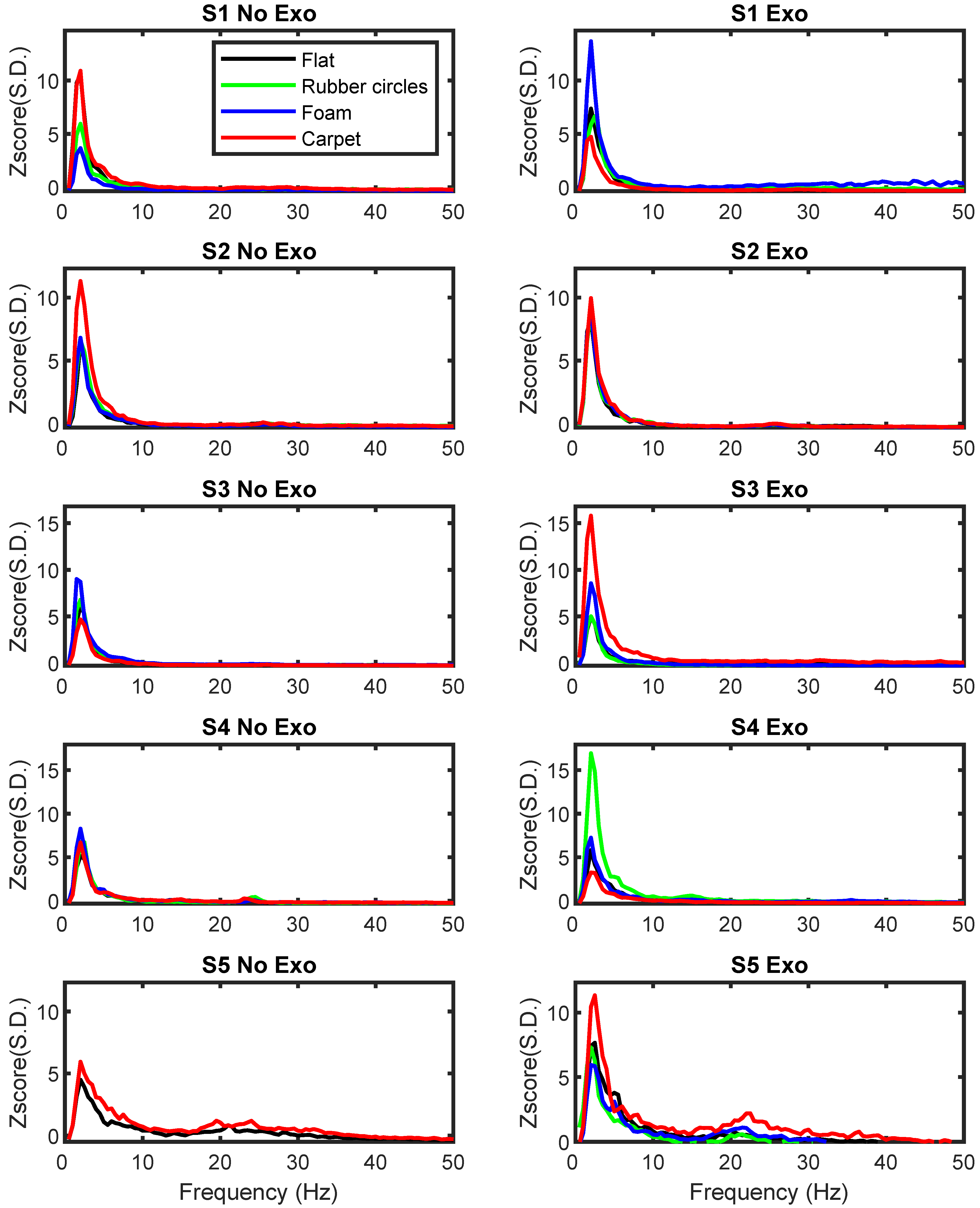
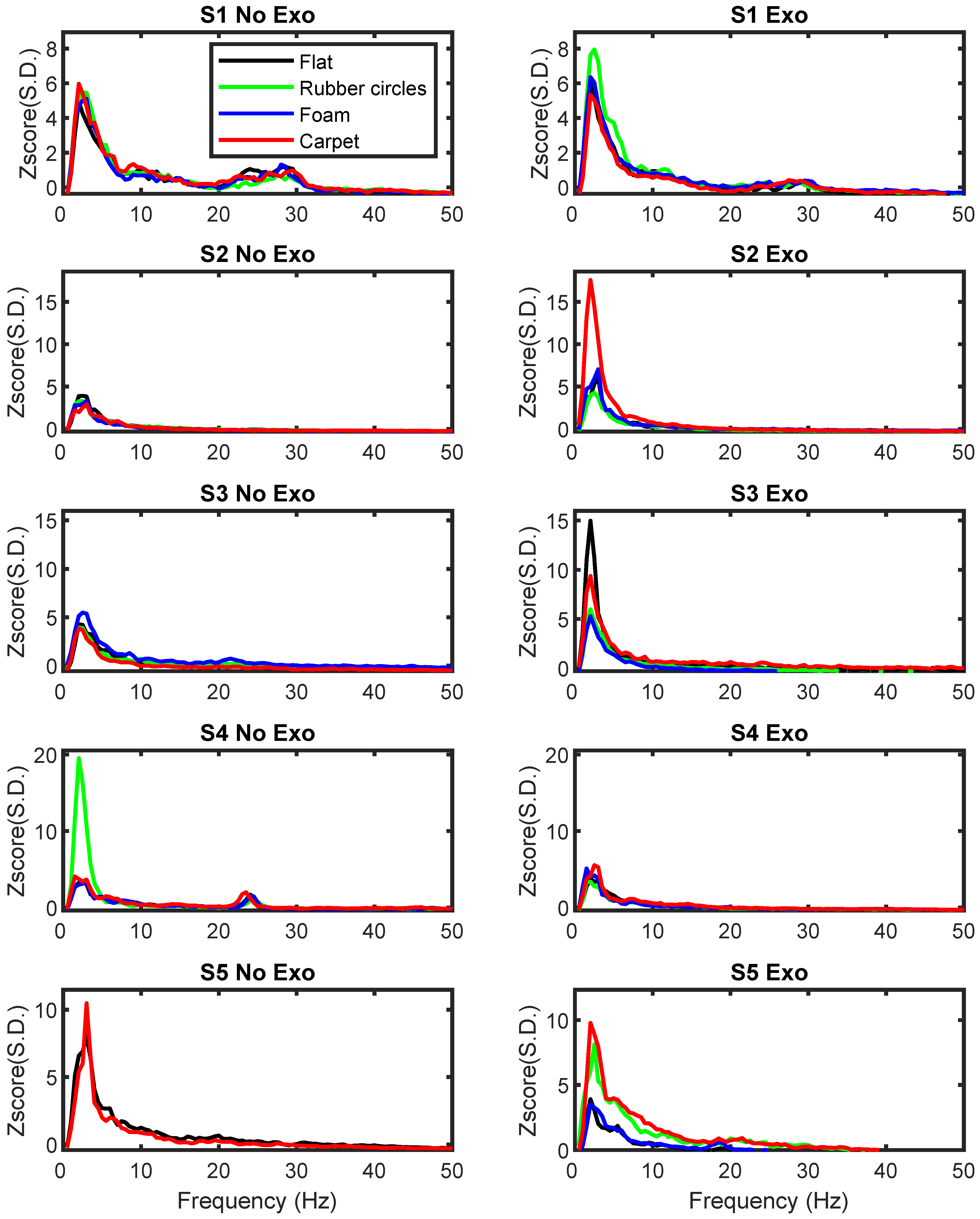
The pilot analysis of the power in different frequency bands at electrodes C3 and C4 was initially performed for the flat pavement to allow for a comparison between the Exo and No Exo conditions. As indicated in the methods section, no statistical tests were performed due to the small number of subjects studied and the large number of conditions. The pilot analysis of the EEG signal power at each frequency band for electrodes C3 and C4 revealed that the Exoskeleton and No Exoskeleton conditions in the flat texture differed mainly in the delta frequency band (see
Figure A1 and
Figure A2 in
Appendix A.2). This difference was particularly noticeable for electrode C3 in subjects S3 (delta—No exo: 2.0097
V
/Hz; Exo: 4.3148
V
/Hz) and S5 (delta—No exo: 3.7755
V
/Hz; Exo: 1.6442
V
/Hz), and for electrode C4 in subjects S1 (delta—No exo: 2.8825
V
/Hz; Exo: 2.1630
V
/Hz) and S2 (delta—No exo: 1.7088
V
/Hz; Exo: 2.2709
V
/Hz). No clear difference was found for subject S4 (delta—No exo: 1.7232
V
/Hz; Exo: 1.9131
V
/Hz).
These findings demonstrated no clear group pattern, with some subjects presenting an increase in the power of specific frequency bands, while others showed a decrease or no change. Expanding this pilot analysis to the PSD values obtained in different conditions and textures further supported the notion that the major differences between textures occurred in the delta frequency band (see
Table 3 and
Figure A1 and
Figure A2 in
Appendix A.2).
Although the differences were visible, no clear pattern could be identified. The analysis of electrode C3 revealed that subjects S2 and S3 presented an increase in power when specific textures were used during the Exoskeleton condition (S1: rubber circles; S2: carpet; S3: flat and carpet), while subject S5 showed a decrease (S5: flat). Similarly, the analysis of electrode C4 revealed increases in the delta frequency bands for all subjects (S1: foam; S2: flat, rubber circles, and foam; S3: carpet; S4: rubber circles; S5: carpet) and decreases for subjects S1 and S2 (S1: carpet; S2: carpet; S4: carpet).
In the remaining conditions, no major differences were found for each of the frequency bands.
3.2. Classifiers’ Performance Outcomes during Exoskeleton Control
on Different Pavements
To complement the previous pilot analysis of the results, the neural activity recorded during the various conditions (exoskeleton and control) while walking on different pavements (flat, rubber circles, carpet, and foam) that formed an eight-class problem was examined using four different classifiers: ANN, KNN, SVM, and LDA.
The performance metrics used to assess the models’ classification abilities are presented in the
Table 4. Note that only one value is presented for each classifier (i.e., no average or standard deviation was calculated), because a single classification was performed, which included all subjects in all eight different conditions simultaneously.
As shown in
Table 4 and
Figure 3, the LDA model demonstrated the highest performance, wherein it achieved an accuracy of 85.71%, a recall of 86.71%, a precision of 86.44%, and an F1 score of 0.87.
4. Discussion
A small sample of subjects controlling an exoskeleton was tested on different textured pavements while their neural activity was recorded. To the best of our knowledge, this is the first study to investigate the neural activity associated with different pavement textures while subjects controlled an exoskeleton. The analysis of the neural activity associated with tactile processing revealed an overall pattern of increases and decreases in power within the delta frequency bands of the C3 and C4 electrodes when the exoskeleton was used. However, no clear pattern was observed with respect to the different textured pavements. When utilizing four different classifiers, the results showed high levels of decoding accuracy, with the LDA classifier demonstrating the best performance. Collectively, these findings suggest that the use of an exoskeleton is associated with changes in the neural activity in electrodes C3 and C4, as well as that LDA is effective in classifying these differences.
Exoskeleton control was associated with changes in neural activity. The pilot analysis of the power in the C3 and C4 electrodes revealed that the use of the exoskeleton in different pavements was associated with changes occurring mostly in the delta band. This finding from the pilot analysis was further supported by the the classifiers’ performance outcomes. Our results are in line with previous studies highlighting the role of the delta frequency band in upper and lower limb exoskeleton control [
38,
39,
40,
41,
42]. An alternative explanation for these findings could be the increases in attention, perception, and decision making [
43,
44,
45]. This alternative explanation also fits with self-reports of subjects indicating the need to pay attention to the exoskeleton movements and balance during the task. It will be relevant in future studies to determine if the continuous use of the exoskeleton results in a decrease in the power of the delta frequency band for this electrode.
In summary, based on the presented results, significant changes were observed in the delta frequency bands of the C3 and C4 electrodes within both the Exo and No Exo (control) conditions while walking on the four different pavements. Among the four classifiers used, LDA achieved the best performance. Additionally, it is worth noting that, although the neural patterns could not be inferred during the pilot analysis, all implemented machine-learning models were able to distinguish between all experimental conditions, albeit with varying degrees of performance.
4.1. Neural Correlates of Pavement Textures
No clear differences between pavement textures were present for any participant. Although previous studies have reported changes in the EEG power when different textures and stimuli are presented [
4,
28,
46,
47] the present findings did not reveal a clear pattern. It is noteworthy that, in all participants, the power for one or more textures was associated with a large difference from the remaining textures, but such differences could not consistently be associated with self-reports of increased attention, difficulty, preference, tiredness, nor novelty (i.e., being the first or last texture).
Previous studies support the notion that tactile processing is associated with power changes in the alpha and beta frequency bands in C3 and C4 electrodes, such as after tactile stimulation of the index finger [
4,
28,
48]. Meanwhile, increases in the delta frequency bands of the C3 and C4 electrodes when different tactile stimuli were delivered to subjects have recently been reported [
46]. Although previous reports have indicated that the most prominent changes occurred in the alpha and beta frequency bands, the aforementioned study has shown the presence of power changes in the delta and low gamma frequency bands, although in a less pronounced form [
46]. Therefore, the data gathered here supports H1 (that the use of the exoskeleton is associated with changes in neural activity), but the data does not support H2 (that the alpha and theta frequencies would be associated with less stable texture/pavements).
4.2. Eight-Class Classification Problem
The comparison of four different classifiers generally revealed good performance outcomes (accuracy values no lower than 63.61%), with linear discriminant analysis presenting the best performance outcomes across all parameters studied. LDA has been a popular choice in many brain–machine interface studies [
49,
50] due to its simplicity and ability to distinguish across different classes. Its performance with respect to binary classification problems makes this model suitable for this type of classification [
51,
52].
LDA and the SVM are among the most popular classifiers for BMI applications, with some studies reporting improved performance for SVM and others for LDA [
14,
36,
53]. Our present results are in line with these previous studies, as we observed good performance for both algorithms in our study, although LDA exhibited the best performance.
Despite the good performance outcomes, there is still room for improvement. For example, in future studies, it will be relevant to test different k values for the KNN model, as well as different numbers of hidden layers and neurons for the ANN classifier [
54,
55,
56,
57].
Altogether, the results from the ML algorithms support H3 (that the neural activity associated with different textured pavements could be identified using common classifiers).
4.3. Active and Passive Tactile Information Processing
The present study did not require subjects to actively process tactile information. Therefore, except for minor adjustments in posture or balance that could have been necessary on different pavements, the subjects performed the act of walking in place, mostly in the same manner, across the different pavements. Active and passive tactile discrimination are known to have a significant impact on EEG signals [
5,
46]; therefore, it is possible that future studies, namely, those involving brain–computer interfaces that combine the neural control of the exoskeleton with different textured pavements, may reveal other relevant neural correlates of pavement texture during exoskeleton control.
4.4. Relevance of Tactile Processing for Exoskeleton Control
Tactile processing during exoskeleton control is relevant, because exoskeletons (and brain-controlled exoskeletons) can cause severe accidents and lead to injuries [
58]. Therefore, incorporating tactile information into these robotic devices, as well as in brain–machine interfaces, can impact exoskeleton control [
1,
46]. Lastly, tactile processing and feedback during exoskeleton control are crucial for neurorehabilitation processes [
1]. The present study supports and extends these previous findings by demonstrating that texture information can be decoded from the C3 and C4 electrodes during exoskeleton control.
4.5. Technical Details and Caveats
A few caveats and technical details should be taken into account. We used a 16-electrode system, which did not allow for the identification of the source of the signals [
59]. Therefore, the results can only be discussed based on the recorded electrode locations and not on the specific brain regions underneath them. Additionally, our sample size was small, which consisted of only five female subjects. In future studies, it will be important to increase the number of electrodes and subjects while ensuring a better representation of sex and age.







{kind=link}
{kind=link}
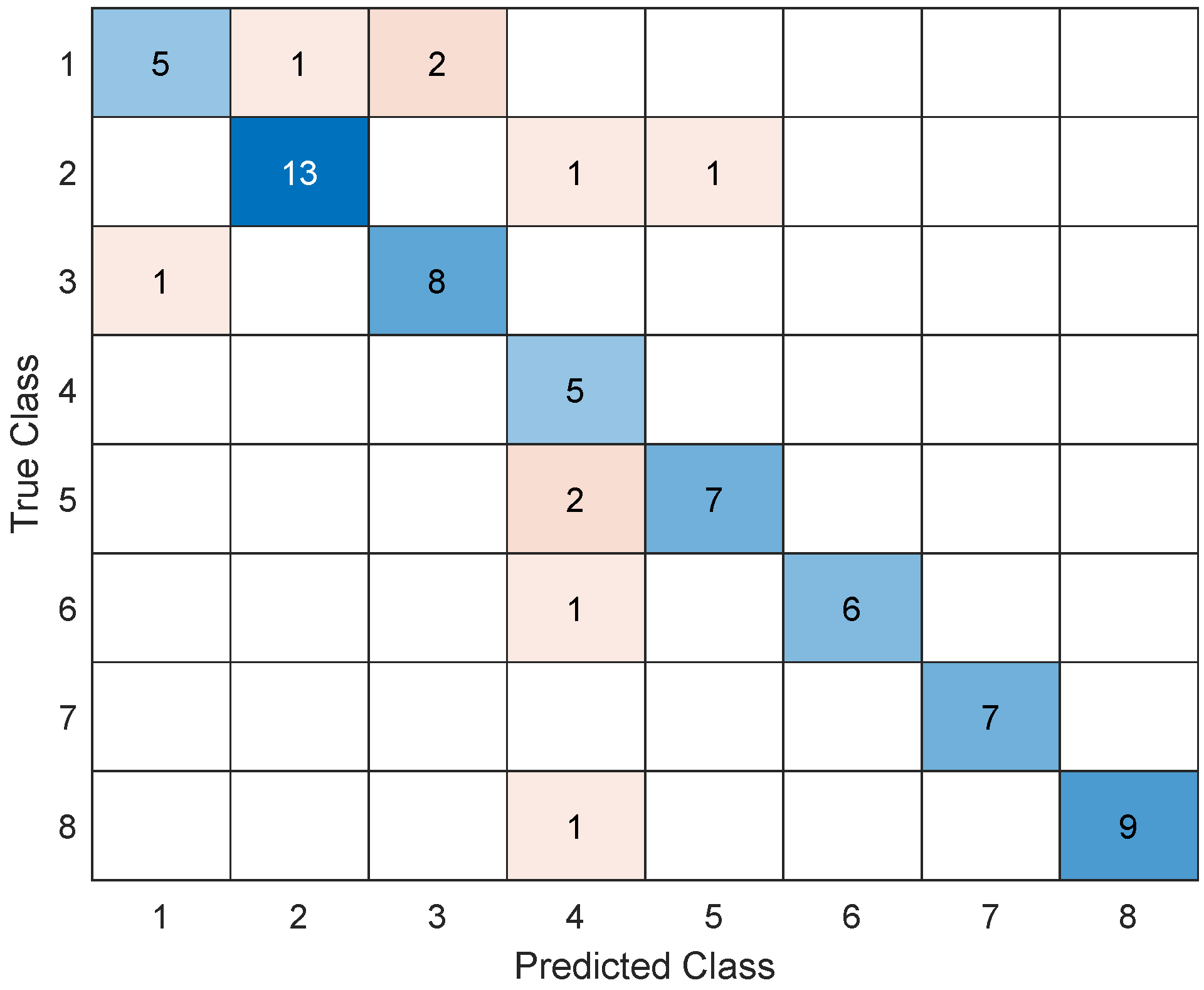{kind=link}
{kind=link}
{kind=link}




