
An Investigation into the Mechanism of Alkaline Extraction-Isoelectric Point Precipitation (AE-IEP) of High-Thiol Plant Proteins


Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
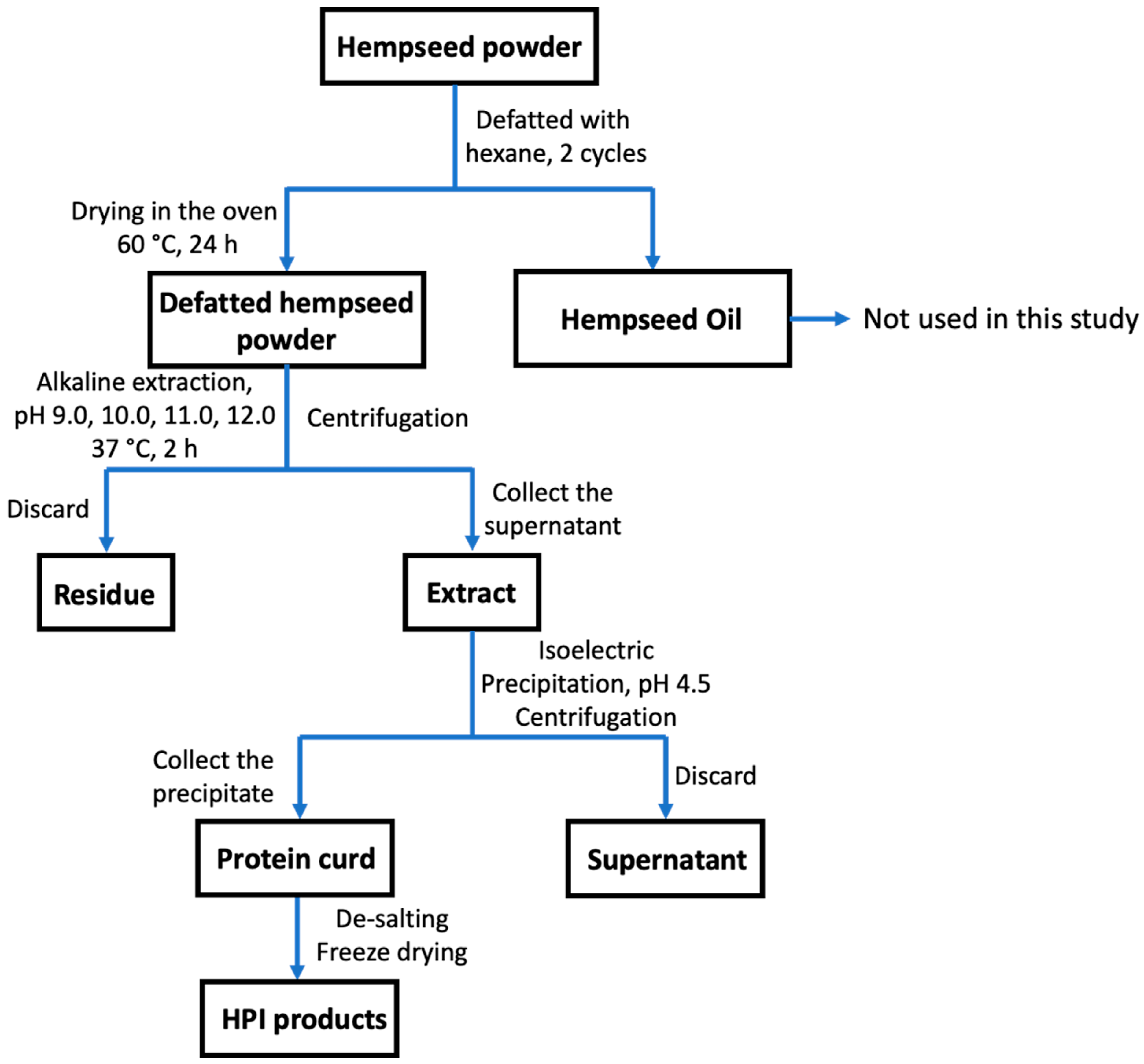
2.2. Hempseed Protein Alkaline Extraction
2.2.1. The Preparation of HPI Products
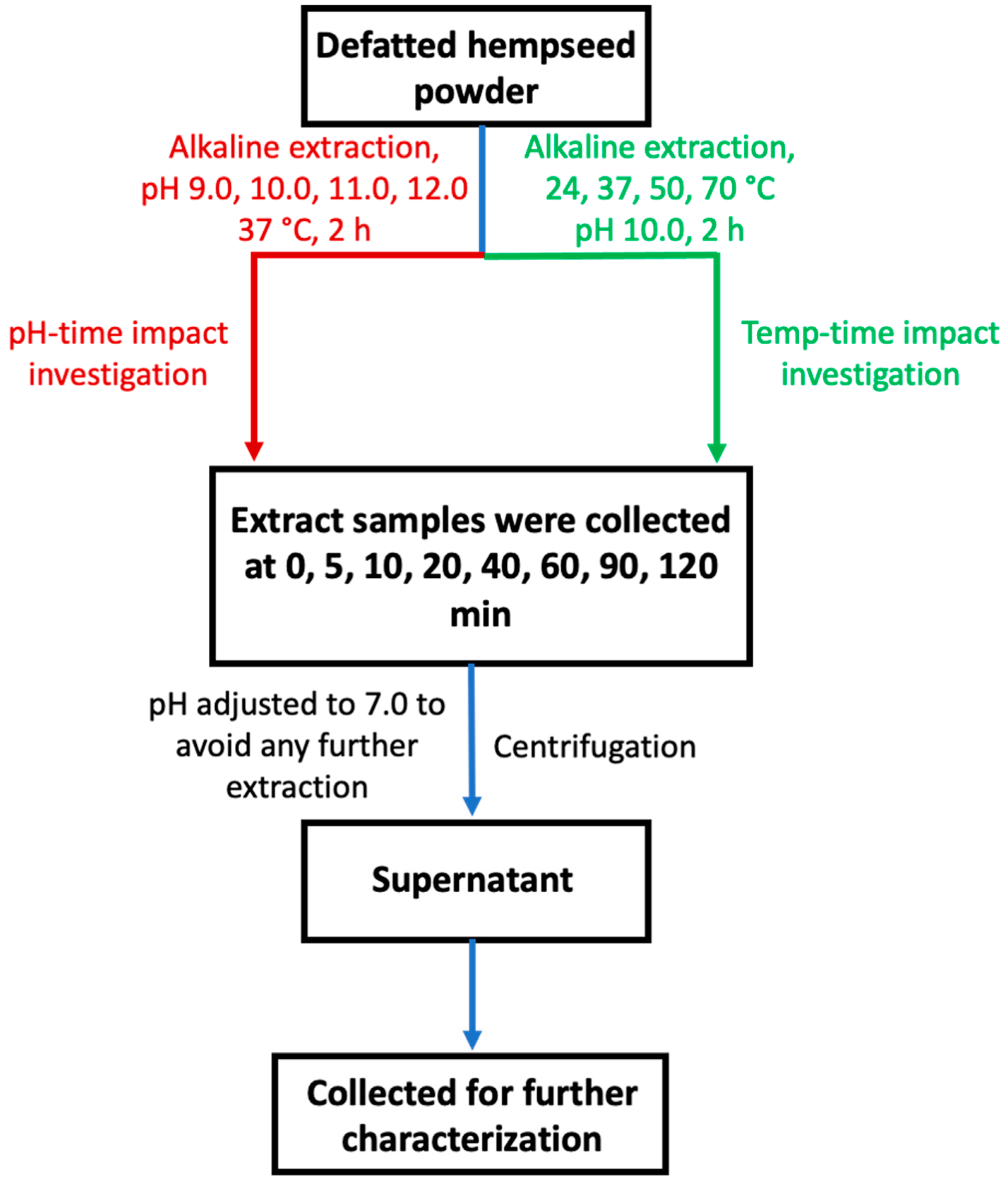
2.2.2. Mechanistic Investigation
2.3. Sodium Dodecyl Sulphate-Polyacrylamide Gel Electrophoresis (SDS-PAGE)
2.4. Protein Extraction Determination
2.5. Determination of Free Sulfhydryl (-SH) Group Content
2.6. Investigation of Hempseed Protein Functionality
2.6.1. Effect of Extraction Protocol on Protein Emulsifying Capacity
2.6.2. Particle Size and Microstructure Determination
2.6.3. Surface Tension
2.7. Percentage Protein Solubility
2.8. Zeta Potential
2.9. Extraction Protocol Optimization Based on Response Surface Methodology (RSM)
2.10. Statistical Analysis
3. Results and Discussions
3.1. Mechanistic Investigation of Hempseed Protein AE-IEP Extraction
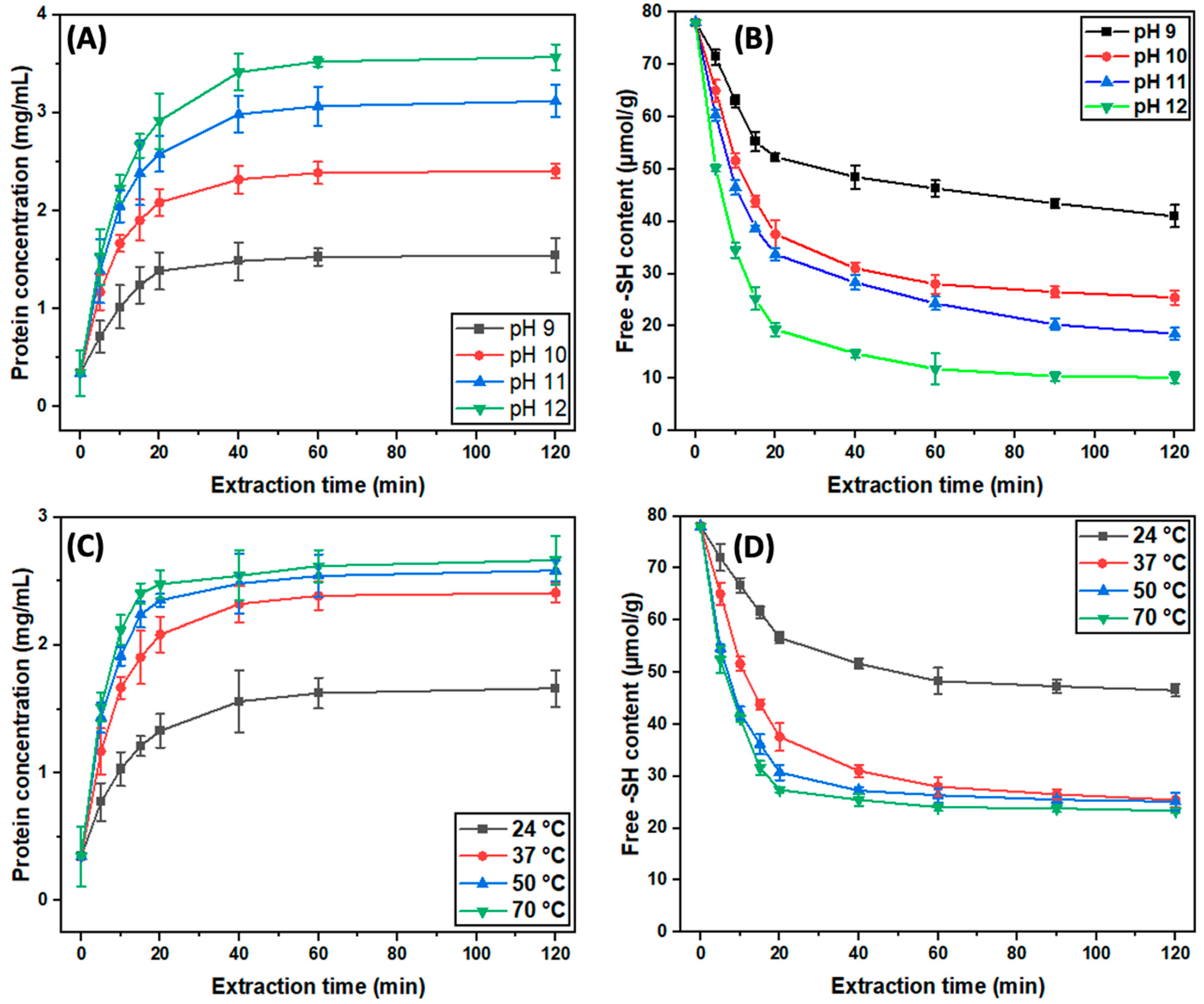
3.1.1. The Effect of pH and Temperature on Protein Extraction Kinetics
3.1.2. The Effect of pH and Temperature on Free Sulfhydryl Content during Extraction
3.2. Evidence for Intermolecular Disulphide Bond Formation
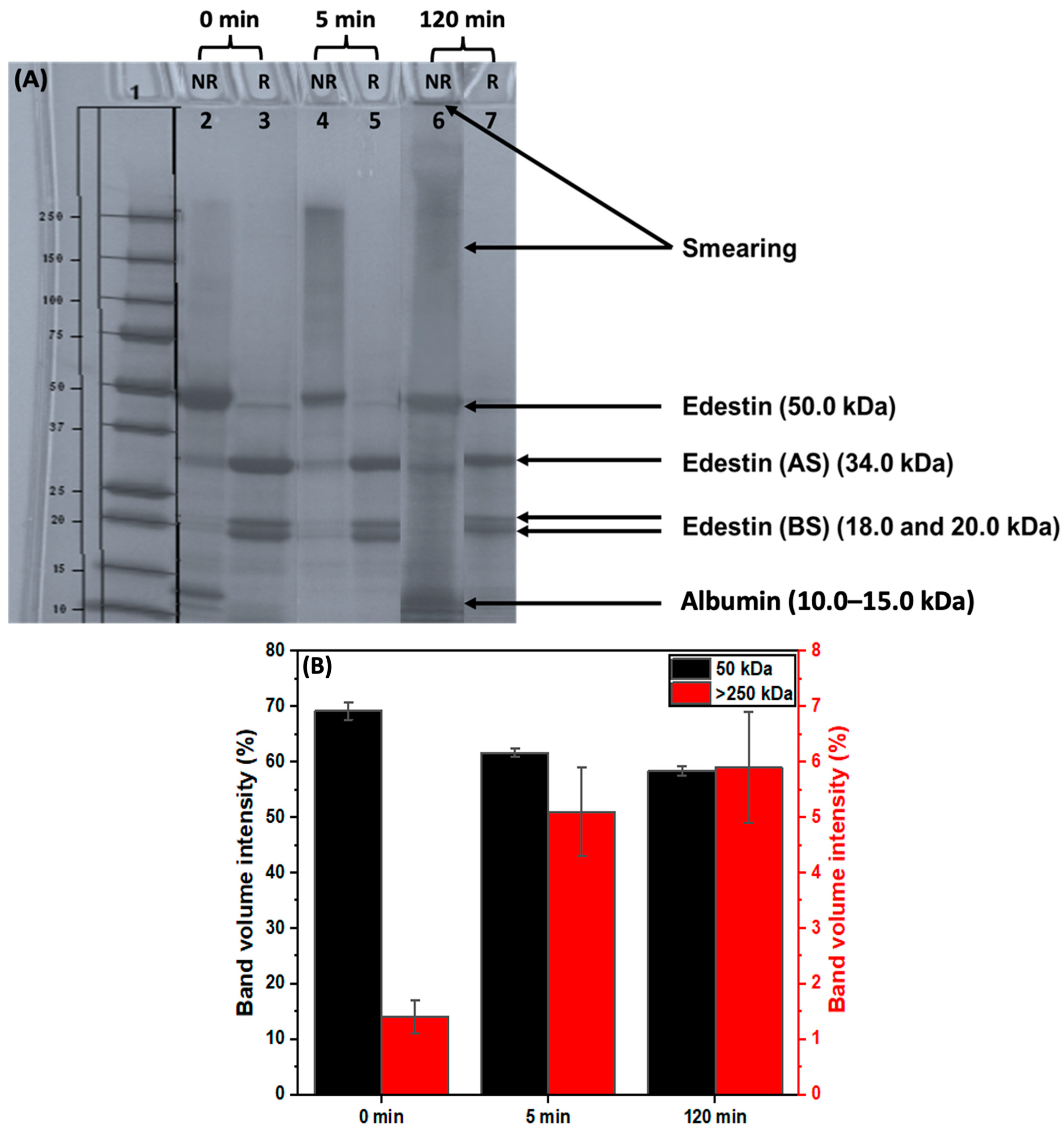
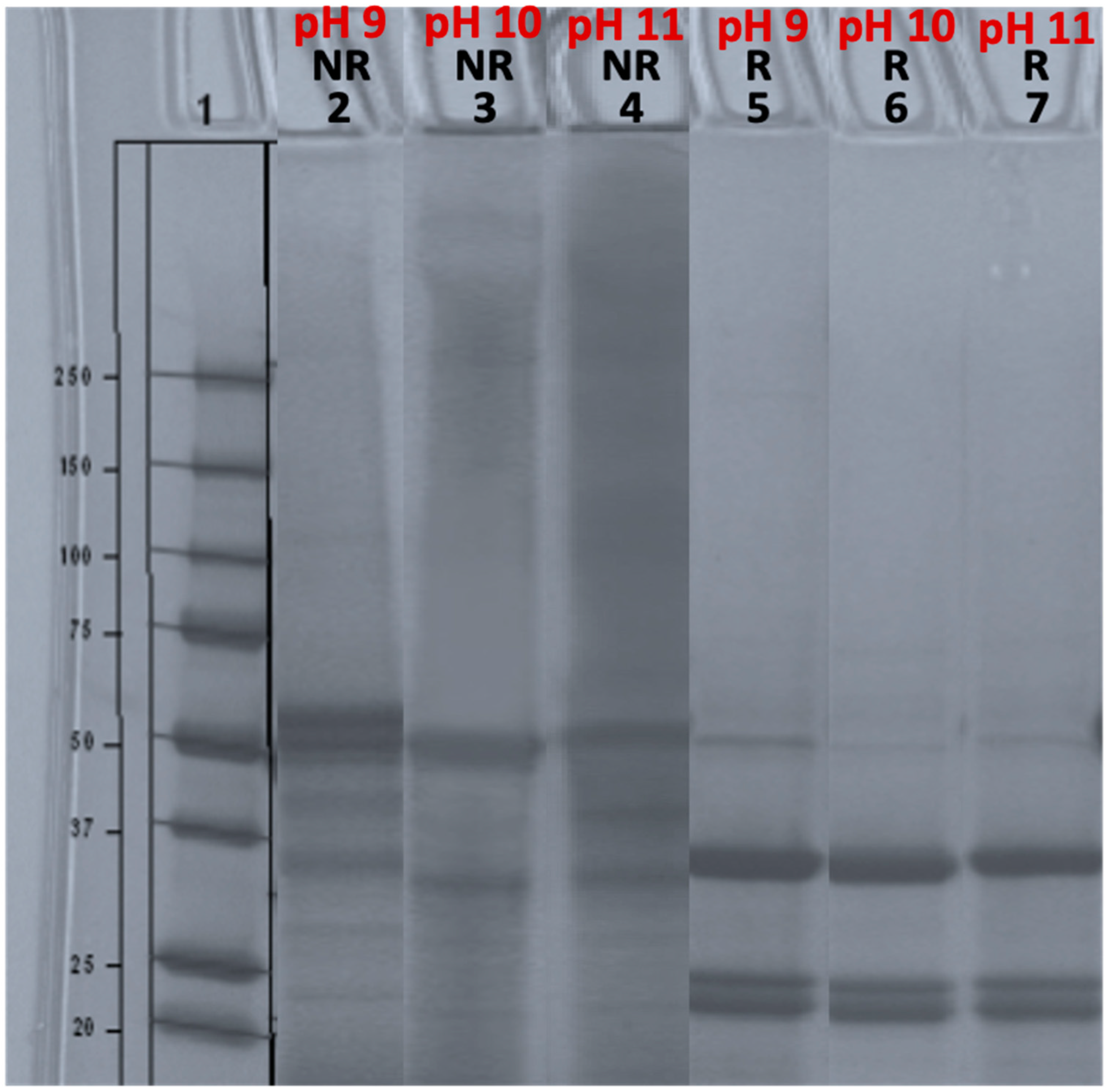
3.2.1. The Impact of Extraction on Hempseed Protein Subunits
3.2.2. The Impact of Extraction pH on Hempseed Protein Subunits
3.3. The Influence of Disulphide Bond Formation on Protein Functionality
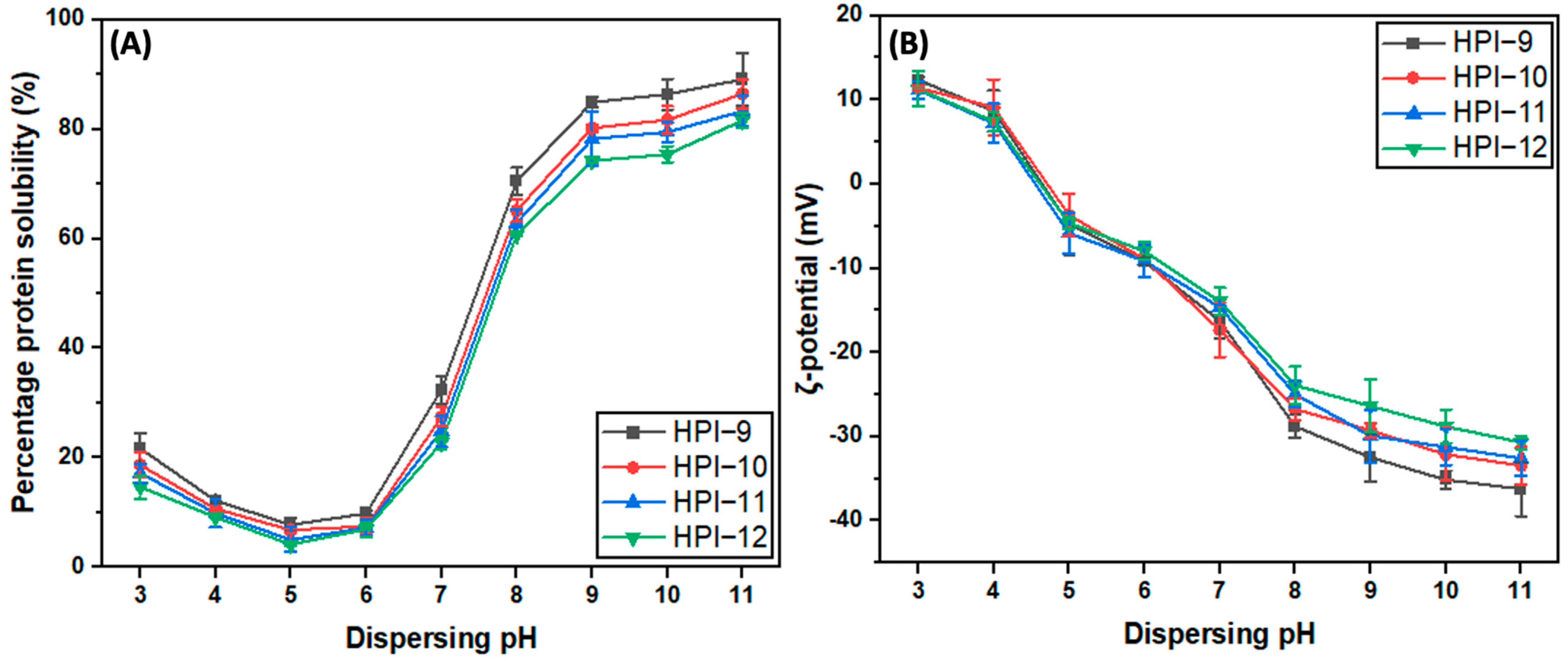
3.3.1. Protein Solubility and ζ-Potential
3.3.2. Emulsifying Properties
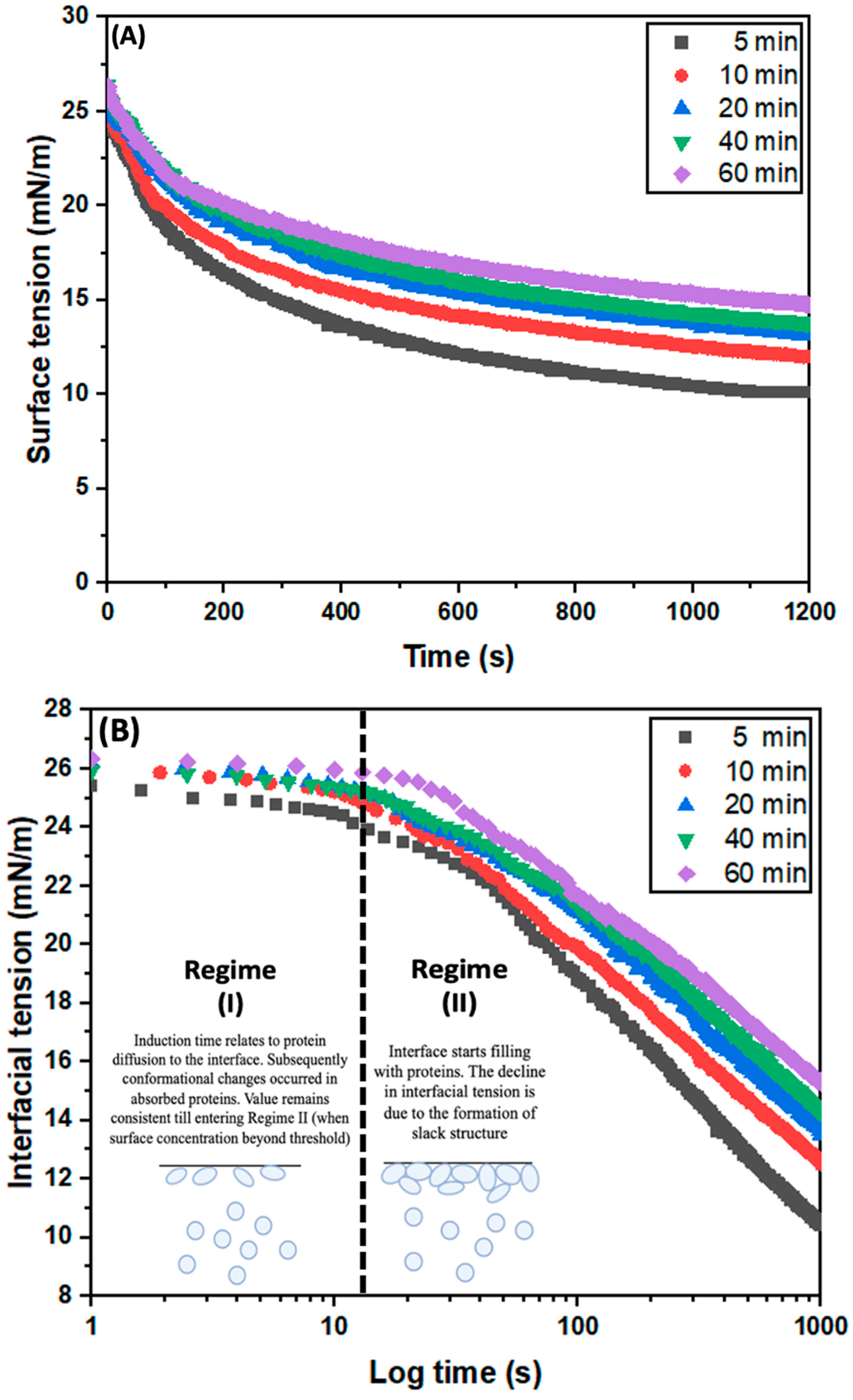
3.3.3. Surface Tension (γ)

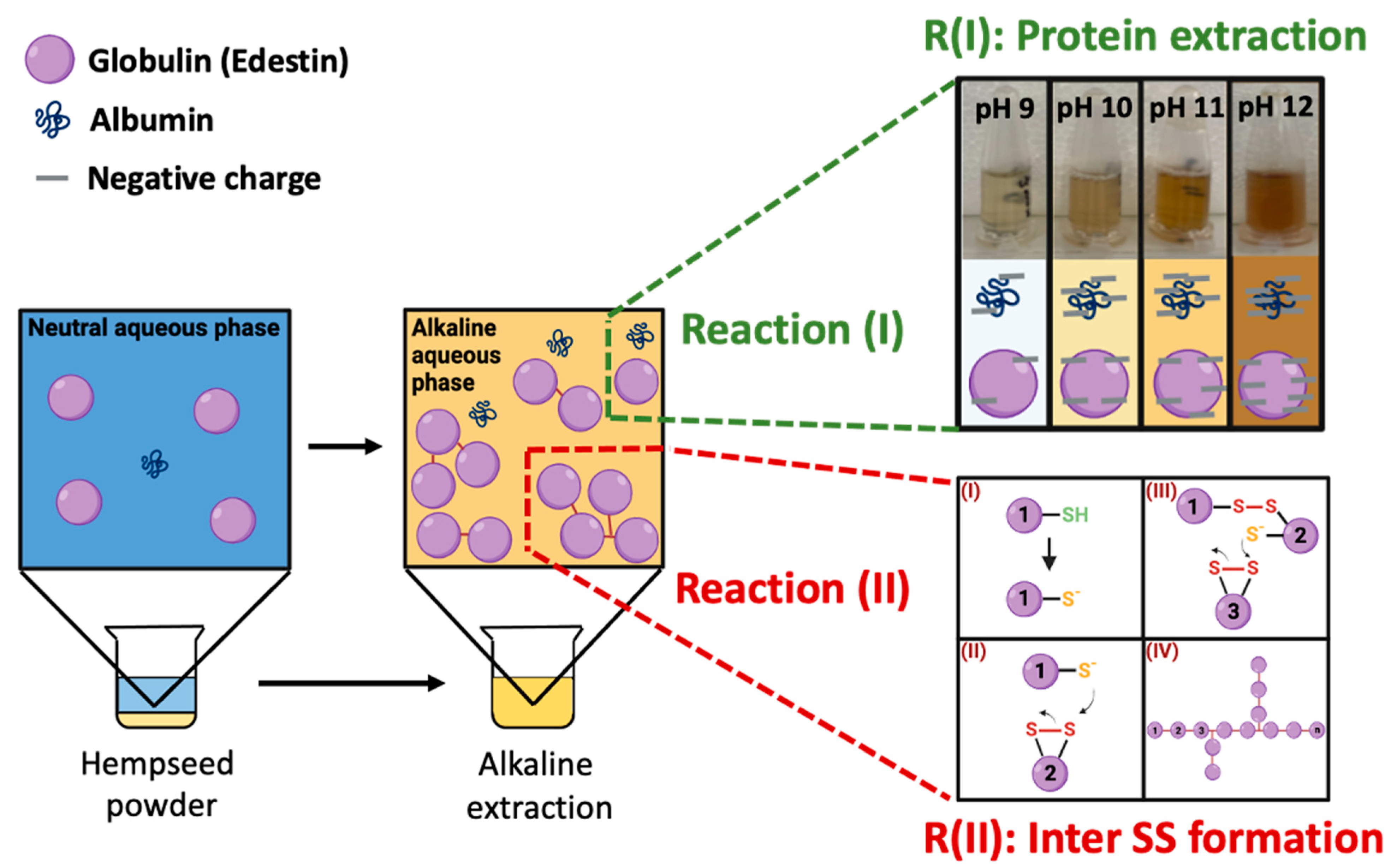
3.4. Proposed Mechanism Underlying Hempseed Protein Extraction
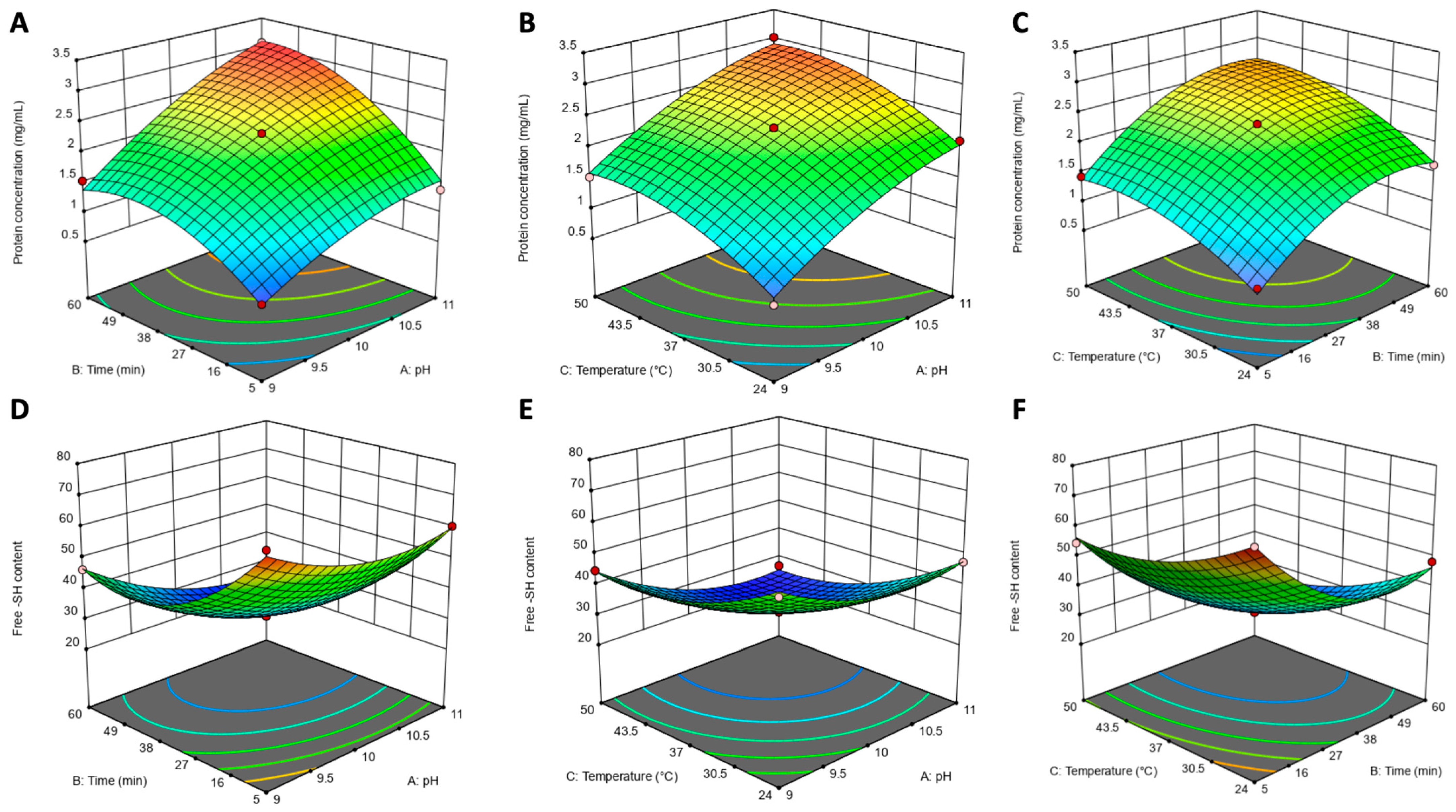
3.5. RSM Optimization of Hempseed Protein Extraction
3.5.1. Model Fitting
3.5.2. The Potential Impact of Optimising Extraction Conditions on HPI Functionality
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Callaway, J.C. Hempseed as a nutritional resource: An overview. Euphytica 2004, 140, 65–72. [Google Scholar] [CrossRef]
- Shen, P.; Gao, Z.; Xu, M.; Ohm, J.-B.; Rao, J.; Chen, B. The impact of hempseed dehulling on chemical composition, structure properties and aromatic profile of hemp protein isolate. Food Hydrocoll. 2020, 106, 105889. [Google Scholar] [CrossRef]
- Dapčević-Hadnađev, T.; Hadnađev, M.; Dizdar, M.; Lješković, N.J. Functional and Bioactive Properties of Hemp Proteins. In Sustainable Agriculture Reviews; Springer: Berlin/Heidelberg, Germany, 2020; Volume 42, pp. 239–263. [Google Scholar]
- Malomo, S.A.; He, R.; Aluko, R.E. Structural and Functional Properties of Hemp Seed Protein Products. J. Food Sci. 2014, 79, C1512–C1521. [Google Scholar] [CrossRef]
- Potin, F.; Lubbers, S.; Husson, F.; Saurel, R. Hemp (Cannabis sativa L.) Protein Extraction Conditions Affect Extraction Yield and Protein Quality. J. Food Sci. 2019, 84, 3682–3690. [Google Scholar] [CrossRef] [PubMed]
- Hadnađev, M.; Dapčević-Hadnađev, T.; Lazaridou, A.; Moschakis, T.; Michaelidou, A.-M.; Popović, S.; Biliaderis, C.G. Hempseed meal protein isolates prepared by different isolation techniques. Part I. physicochemical properties. Food Hydrocoll. 2018, 79, 526–533. [Google Scholar] [CrossRef]
- Wang, Q.; Xiong, Y.L. Processing, nutrition, and functionality of hempseed protein: A review. Compr. Rev. Food Sci. Food Saf. 2019, 18, 936–952. [Google Scholar] [CrossRef]
- Malomo, S.A.; Aluko, R.E. A comparative study of the structural and functional properties of isolated hemp seed (Cannabis sativa L.) albumin and globulin fractions. Food Hydrocoll. 2015, 43, 743–752. [Google Scholar] [CrossRef]
- Dapčević-Hadnađev, T.; Dizdar, M.; Pojić, M.; Krstonošić, V.; Zychowski, L.M.; Hadnađev, M. Emulsifying properties of hemp proteins: Effect of isolation technique. Food Hydrocoll. 2019, 89, 912–920. [Google Scholar] [CrossRef]
- Teh, S.-S.; Bekhit, A.E.-D.; Carne, A.; Birch, J. Effect of the defatting process, acid and alkali extraction on the physicochemical and functional properties of hemp, flax and canola seed cake protein isolates. J. Food Meas. Charact. 2014, 8, 92–104. [Google Scholar] [CrossRef]
- Odani, S.; Odani, S. Isolation and primary structure of a methionine-and cystine-rich seed protein of Cannabis sativa. Biosci. Biotechnol. Biochem. 1998, 62, 650–654. [Google Scholar] [CrossRef]
- Patel, S.; Cudney, R.; McPherson, A. Crystallographic characterization and molecular symmetry of edestin, a legumin from hemp. J. Mol. Biol. 1994, 235, 361–363. [Google Scholar] [CrossRef] [PubMed]
- Docimo, T.; Caruso, I.; Ponzoni, E.; Mattana, M.; Galasso, I. Molecular characterization of edestin gene family in Cannabis sativa L. Plant Physiol. Biochem. 2014, 84, 142–148. [Google Scholar] [CrossRef] [PubMed]
- Beilinson, V.; Chen, Z.; Shoemaker, R.; Fischer, R.; Goldberg, R.; Nielsen, N. Genomic organization of glycinin genes in soybean. Theor. Appl. Genet. 2002, 104, 1132–1140. [Google Scholar] [CrossRef] [PubMed]
- Tang, C.-H.; Ten, Z.; Wang, X.-S.; Yang, X.-Q. Physicochemical and functional properties of hemp (Cannabis sativa L.) protein isolate. J. Agric. Food Chem. 2006, 54, 8945–8950. [Google Scholar] [CrossRef]
- Wang, X.-S.; Tang, C.-H.; Yang, X.-Q.; Gao, W.-R. Characterization, amino acid composition and in vitro digestibility of hemp (Cannabis sativa L.) proteins. Food Chem. 2008, 107, 11–18. [Google Scholar] [CrossRef]
- Wang, S.; Wang, J.; Xue, F.; Li, C. Effects of heating or ultrasound treatment on the enzymolysis and the structure characterization of hempseed protein isolates. J. Food Sci. Technol. 2019, 56, 3337–3346. [Google Scholar] [CrossRef]
- Wang, Q.; Jin, Y.; Xiong, Y.L. Heating-Aided pH Shifting Modifies Hemp Seed Protein Structure, Cross-Linking, and Emulsifying Properties. J. Agric. Food Chem. 2018, 66, 10827–10834. [Google Scholar] [CrossRef]
- Yin, S.-W.; Tang, C.-H.; Cao, J.-S.; Hu, E.-K.; Wen, Q.-B.; Yang, X.-Q. Effects of limited enzymatic hydrolysis with trypsin on the functional properties of hemp (Cannabis sativa L.) protein isolate. Food Chem. 2008, 106, 1004–1013. [Google Scholar] [CrossRef]
- Kim, K.S.; Rhee, J.S. Effects of Acetylation on Physicochemical Properties of J1s Soy Protein. J. Food Biochem. 1989, 13, 187–199. [Google Scholar] [CrossRef]
- Ma, C.-Y.; Wood, D.F. Functional properties of oat proteins modified by acylation, trypsin hydrolysis or linoleate treatment. J. Am. Oil Chem. Soc. 1987, 64, 1726–1731. [Google Scholar] [CrossRef]
- Yin, S.W.; Tang, C.H.; Wen, Q.B.; Yang, X.Q. Functional and structural properties and in vitro digestibility of acylated hemp (Cannabis sativa L.) protein isolates. Int. J. Food Sci. Technol. 2009, 44, 2653–2661. [Google Scholar] [CrossRef]
- Shaaban, S.; Negm, A.; Sobh, M.A.; Wessjohann, L.A. Organoselenocyanates and symmetrical diselenides redox modulators: Design, synthesis and biological evaluation. Eur. J. Med. Chem. 2015, 97, 190–201. [Google Scholar] [CrossRef]
- Li, Y.; Cheng, Y.; Zhang, Z.; Wang, Y.; Mintah, B.K.; Dabbour, M.; Jiang, H.; He, R.; Ma, H. Modification of rapeseed protein by ultrasound-assisted pH shift treatment: Ultrasonic mode and frequency screening, changes in protein solubility and structural characteristics. Ultrason. Sonochem. 2020, 69, 105240. [Google Scholar] [CrossRef] [PubMed]
- Matsudomi, N.; Sasaki, T.; Kato, A.; Kobayashi, K. Conformational changes and functional properties of acid-modified soy protein. Agric. Biol. Chem. 1985, 49, 1251–1256. [Google Scholar]
- Nissen, S.H.; Schmidt, J.M.; Gregersen, S.; Hammershøj, M.; Møller, A.H.; Danielsen, M.; Stødkilde, L.; Nebel, C.; Dalsgaard, T.K. Increased solubility and functional properties of precipitated Alfalfa protein concentrate subjected to pH shift processes. Food Hydrocoll. 2021, 119, 106874. [Google Scholar] [CrossRef]
- Jiang, J.; Zhu, B.; Liu, Y.; Xiong, Y.L. Interfacial structural role of pH-shifting processed pea protein in the oxidative stability of oil/water emulsions. J. Agric. Food Chem. 2014, 62, 1683–1691. [Google Scholar] [CrossRef]
- Gao, Z.; Shen, P.; Lan, Y.; Cui, L.; Ohm, J.-B.; Chen, B.; Rao, J. Effect of alkaline extraction pH on structure properties, solubility, and beany flavor of yellow pea protein isolate. Food Res. Int. 2020, 131, 109045. [Google Scholar] [CrossRef]
- Ren, Y.; Liang, K.; Jin, Y.; Zhang, M.; Chen, Y.; Wu, H.; Lai, F. Identification and characterization of two novel α-glucosidase inhibitory oligopeptides from hemp (Cannabis sativa L.) seed protein. J. Funct. Foods 2016, 26, 439–450. [Google Scholar] [CrossRef]
- Orio, L.P.; Boschin, G.; Recca, T.; Morelli, C.F.; Ragona, L.; Francescato, P.; Arnoldi, A.; Speranza, G. New ACE-inhibitory peptides from hemp seed (Cannabis sativa L.) proteins. J. Agric. Food Chem. 2017, 65, 10482–10488. [Google Scholar] [CrossRef]
- Lu, R.-R.; Qian, P.; Sun, Z.; Zhou, X.-H.; Chen, T.-P.; He, J.-F.; Zhang, H.; Wu, J. Hempseed protein derived antioxidative peptides: Purification, identification and protection from hydrogen peroxide-induced apoptosis in PC12 cells. Food Chem. 2010, 123, 1210–1218. [Google Scholar] [CrossRef]
- Raikos, V.; Duthie, G.; Ranawana, V. Denaturation and Oxidative Stability of Hemp Seed (Cannabis sativa L.) Protein Isolate as Affected by Heat Treatment. Plant Foods Hum. Nutr. 2015, 70, 304–309. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Zheng, Y.; Zhong, Y.; Wang, D.; Deng, Y. Casein-hempseed protein complex via cross-link catalyzed by transglutaminase for improving structural, rheological, emulsifying and gelation properties. Food Chem. 2022, 383, 132366. [Google Scholar] [CrossRef] [PubMed]
- Potin, F.; Goure, E.; Lubbers, S.; Husson, F.; Saurel, R. Functional properties of hemp protein concentrate obtained by alkaline extraction and successive ultrafiltration and spray-drying. Int. J. Food Sci. Technol. 2022, 57, 436–446. [Google Scholar] [CrossRef]
- Mamone, G.; Picariello, G.; Ramondo, A.; Nicolai, M.A.; Ferranti, P. Production, digestibility and allergenicity of hemp (Cannabis sativa L.) protein isolates. Food Res. Int. 2019, 115, 562–571. [Google Scholar] [CrossRef]
- Drinić, Z.; Vladic, J.; Koren, A.; Zeremski, T.; Stojanov, N.; Tomić, M.; Vidović, S. Application of conventional and high-pressure extraction techniques for the isolation of bioactive compounds from the aerial part of hemp (Cannabis sativa L.) assortment Helena. Ind. Crop. Prod. 2021, 171, 113908. [Google Scholar] [CrossRef]
- Wall, S.B.; Oh, J.-Y.; Diers, A.R.; Landar, A. Oxidative Modification of Proteins: An Emerging Mechanism of Cell Signaling. Front. Physiol. 2012, 3, 369. [Google Scholar] [CrossRef]
- LoPachin, R.M.; Gavin, T. Reactions of electrophiles with nucleophilic thiolate sites: Relevance to pathophysiological mechanisms and remediation. Free. Radic. Res. 2016, 50, 195–205. [Google Scholar] [CrossRef]
- Yao, S.; Li, W.; Wu, Y.; Martin, G.J.O.; Ashokkumar, M. The Impact of High-Intensity Ultrasound-Assisted Extraction on the Structural and Functional Properties of Hempseed Protein Isolate (HPI). Foods 2023, 12, 348. [Google Scholar] [CrossRef]
- Wu, Y.; Li, W.; Martin, G.J.; Ashokkumar, M. Mechanism of low-frequency and high-frequency ultrasound-induced inactivation of soy trypsin inhibitors. Food Chem. 2021, 360, 130057. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Beveridge, T.; Toma, S.; Nakai, S. Determination of SH- and SS-groups in some food proteins using Ellman’s reagent. J. Food Sci. 1974, 39, 49–51. [Google Scholar] [CrossRef]
- Ladjal-Ettoumi, Y.; Boudries, H.; Chibane, M.; Romero, A. Pea, Chickpea and Lentil Protein Isolates: Physicochemical Characterization and Emulsifying Properties. Food Biophys. 2016, 11, 43–51. [Google Scholar] [CrossRef]
- Amagliani, L.; O’Regan, J.; Kelly, A.L.; O’Mahony, J.A. The composition, extraction, functionality and applications of rice proteins: A review. Trends Food Sci. Technol. 2017, 64, 1–12. [Google Scholar] [CrossRef]
- Pearce, K.N.; Kinsella, J.E. Emulsifying properties of proteins: Evaluation of a turbidimetric technique. J. Agric. Food Chem. 1978, 26, 716–723. [Google Scholar] [CrossRef]
- Yatipanthalawa, B.; Li, W.; Hill, D.R.; Trifunovic, Z.; Ashokkumar, M.; Scales, P.J.; Martin, G.J. Interplay between interfacial behaviour, cell structure and shear enables biphasic lipid extraction from whole diatom cells (Navicula sp.). J. Colloid Interface Sci. 2021, 589, 65–76. [Google Scholar] [CrossRef] [PubMed]
- Guo, P.; Cao, Z.; Chen, S.; Chen, C.; Liu, J.; Meng, J. Application of Design-Expert Response Surface Methodology for the Optimization of Recycled Asphalt Mixture with Waste Engine Oil. J. Mater. Civ. Eng. 2021, 33, 04021075. [Google Scholar] [CrossRef]
- Shen, P.; Gao, Z.; Xu, M.; Rao, J.; Chen, B. Physicochemical and structural properties of proteins extracted from dehulled industrial hempseeds: Role of defatting process and precipitation pH. Food Hydrocoll. 2020, 108, 106065. [Google Scholar] [CrossRef]
- Sari, Y.W.; Mulder, W.J.; Sanders, J.P.M.; Bruins, M.E. Towards plant protein refinery: Review on protein extraction using alkali and potential enzymatic assistance. Biotechnol. J. 2015, 10, 1138–1157. [Google Scholar] [CrossRef]
- Momen, S.; Alavi, F.; Aider, M. Alkali-mediated treatments for extraction and functional modification of proteins: Critical and application review. Trends Food Sci. Technol. 2021, 110, 778–797. [Google Scholar] [CrossRef]
- Ashori, A.; Ornelas, M.; Sheshmani, S.; Cordeiro, N. Influence of mild alkaline treatment on the cellulosic surfaces active sites. Carbohydr. Polym. 2012, 88, 1293–1298. [Google Scholar] [CrossRef]
- Feyzi, S.; Milani, E.; Golimovahhed, Q.A. Grass Pea (Lathyrus sativus L.) Protein Isolate: The Effect of Extraction Optimization and Drying Methods on the Structure and Functional Properties. Food Hydrocoll. 2018, 74, 187–196. [Google Scholar] [CrossRef]
- Östbring, K.; Nilsson, K.; Ahlström, C.; Fridolfsson, A.; Rayner, M. Emulsifying and Anti-Oxidative Properties of Proteins Extracted from Industrially Cold-Pressed Rapeseed Press-Cake. Foods 2020, 9, 678. [Google Scholar] [CrossRef] [PubMed]
- Gadalkar, S.M.; Gogate, P.R.; Rathod, V.K. Recovery of Proteins from Rice Mill Industry Waste (Rice Bran) Using Alkaline or NaCl-Assisted Alkaline Extraction Processes. J. Food Process. Eng. 2017, 40, e12430. [Google Scholar] [CrossRef]
- Sari, Y.W.; Bruins, M.E.; Sanders, J.P. Enzyme assisted protein extraction from rapeseed, soybean, and microalgae meals. Ind. Crop. Prod. 2013, 43, 78–83. [Google Scholar] [CrossRef]
- Plati, F.; Ritzoulis, C.; Pavlidou, E.; Paraskevopoulou, A. Complex coacervate formation between hemp protein isolate and gum Arabic: Formulation and characterization. Int. J. Biol. Macromol. 2021, 182, 144–153. [Google Scholar] [CrossRef] [PubMed]
- Sereewatthanawut, I.; Prapintip, S.; Watchiraruji, K.; Goto, M.; Sasaki, M.; Shotipruk, A. Extraction of protein and amino acids from deoiled rice bran by subcritical water hydrolysis. Bioresour. Technol. 2008, 99, 555–561. [Google Scholar] [CrossRef]
- L’Hocine, L.; Boye, J.I.; Arcand, Y. Composition and Functional Properties of Soy Protein Isolates Prepared Using Alternative Defatting and Extraction Procedures. J. Food Sci. 2006, 71, C137–C145. [Google Scholar] [CrossRef]
- Stone, A.K.; Karalash, A.; Tyler, R.T.; Warkentin, T.D.; Nickerson, M.T. Functional attributes of pea protein isolates prepared using different extraction methods and cultivars. Food Res. Int. 2015, 76, 31–38. [Google Scholar] [CrossRef]
- Jarpa-Parra, M. Lentil protein: A review of functional properties and food application. An overview of lentil protein functionality. Int. J. Food Sci. Technol. 2018, 53, 892–903. [Google Scholar] [CrossRef]
- Avelar, Z.; Vicente, A.A.; Saraiva, J.A.; Rodrigues, R.M. The role of emergent processing technologies in tailoring plant protein functionality: New insights. Trends Food Sci. Technol. 2021, 113, 219–231. [Google Scholar] [CrossRef]
- Schmitt, C.; Bovay, C.; Rouvet, M.; Shojaei-Rami, S.; Kolodziejczyk, E. Whey Protein Soluble Aggregates from Heating with NaCl: Physicochemical, Interfacial, and Foaming Properties. Langmuir 2007, 23, 4155–4166. [Google Scholar] [CrossRef] [PubMed]
- Alavi, F.; Chen, L.; Wang, Z.; Emam-Djomeh, Z. Consequences of heating under alkaline pH alone or in the presence of maltodextrin on solubility, emulsifying and foaming properties of faba bean protein. Food Hydrocoll. 2021, 112, 106335. [Google Scholar] [CrossRef]
- Tan, S.H.; Mailer, R.J.; Blanchard, C.L.; Agboola, S.O.; Day, L. Gelling properties of protein fractions and protein isolate extracted from Australian canola meal. Food Res. Int. 2014, 62, 819–828. [Google Scholar] [CrossRef]
- Aryee, A.; Agyei, D.; Udenigwe, C. Impact of processing on the chemistry and functionality of food proteins. In Proteins in Food Processing; Elsevier: Amsterdam, The Netherlands, 2018; pp. 27–45. [Google Scholar] [CrossRef]
- Malik, M.A.; Sharma, H.K.; Saini, C.S. High intensity ultrasound treatment of protein isolate extracted from dephenolized sunflower meal: Effect on physicochemical and functional properties. Ultrason. Sonochem. 2017, 39, 511–519. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.; Huang, Z.; Guo, Y.; Li, B.; Yu, W.; Zhou, L.; Jiang, L.; Teng, F.; Wang, Z. Effects of high-pressure homogenization on structural and emulsifying properties of thermally soluble aggregated kidney bean (Phaseolus vulgaris L.) proteins. Food Hydrocoll. 2021, 119, 106835. [Google Scholar] [CrossRef]
- Friberg, S.; Larsson, K.; Sjoblom, J. Food Emulsions; CRC Press: Boca Raton, FL, USA, 2003. [Google Scholar]
- Poon, S.; Clarke, A.E.; Schultz, C.J. Effect of Denaturants on the Emulsifying Activity of Proteins. J. Agric. Food Chem. 2001, 49, 281–286. [Google Scholar] [CrossRef]
- Ward, A.; Tordai, L. Time-dependence of boundary tensions of solutions I. The role of diffusion in time-effects. J. Chem. Phys. 1946, 14, 453–461. [Google Scholar] [CrossRef]
- Beverung, C.; Radke, C.J.; Blanch, H.W. Protein adsorption at the oil/water interface: Characterization of adsorption kinetics by dynamic interfacial tension measurements. Biophys. Chem. 1999, 81, 59–80. [Google Scholar] [CrossRef]
- Abu Baka, N.; Abu-Siada, A.; Islam, S.; El-Naggar, M.F. A new technique to measure interfacial tension of transformer oil using UV-Vis spectroscopy. IEEE Trans. Dielectr. Electr. Insul. 2015, 22, 1275–1282. [Google Scholar] [CrossRef]
- Anfinsen, C.B. Principles that Govern the Folding of Protein Chains. Science 1973, 181, 223–230. [Google Scholar] [CrossRef]
- Fernandes, P.A.; Ramos, M.J. Theoretical Insights into the Mechanism for Thiol/Disulfide Exchange. Chem. A Eur. J. 2004, 10, 257–266. [Google Scholar] [CrossRef]
- Taniyama, Y.; Kuroki, R.; Omura, F.; Seko, C.; Kikuchi, M. Evidence for intramolecular disulfide bond shuffling in the folding of mutant human lysozyme. J. Biol. Chem. 1991, 266, 6456–6461. [Google Scholar] [CrossRef]
- Coghlan, J.; Benet, A.; Kumaran, P.; Ford, M.; Veale, L.; Skilton, S.J.; Saveliev, S.; Schwendeman, A.A. Streamlining the Characterization of Disulfide Bond Shuffling and Protein Degradation in IgG1 Biopharmaceuticals Under Native and Stressed Conditions. Front. Bioeng. Biotechnol. 2022, 10, 311. [Google Scholar] [CrossRef]
- Giustarini, D.; Dalle-Donne, I.; Colombo, R.; Milzani, A.; Rossi, R. Is ascorbate able to reduce disulfide bridges? A cautionary note. Nitric Oxide 2008, 19, 252–258. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Independent Variables | Symbol | Coded Levels | ||
|---|---|---|---|---|
| −1 | 0 | 1 | ||
| Extraction pH | X1 | 9 | 10 | 11 |
| Extraction time (min) | X2 | 5 | 32.5 | 60 |
| Extraction Temp (°C) | X3 | 24 | 37 | 50 |
| Extraction pH | Extraction Yield (%) | Protein Recovery Yield (%) | Free -SH Group in HPI (μmol/g Soluble Protein) |
|---|---|---|---|
| 9.0 | 17.2 ± 0.3 c | 49.2 ± 0.8 c | 41.0 ± 0.2 a |
| 10.0 | 18.7 ± 0.6 c | 51.7 ± 1.6 bc | 25.5 ± 0.4 b |
| 11.0 | 20.7 ± 0.5 b | 55.4 ± 1.3 ab | 18.5 ± 1.1 c |
| 12.0 | 22.4 ± 0.4 a | 59.3 ± 1.0 a | 10.2 ± 0.2 d |
| Extraction Time (min) | Free -SH Content (μmol/g Soluble Protein) | 0-Day Droplet Size D4,3 (μm) | 7th-Day Droplet Size D4,3 (μm) | EAI (m2/g) | Creaming Index (%) |
|---|---|---|---|---|---|
| 5 | 65.0 ± 0.6 a | 11.0 ± 0.9 c | 11.6 ± 0.3 d | 7.1 ± 0.5 a | 14.2 |
| 10 | 51.7 ± 1.3 b | 12.5 ± 0.8 c | 13.6 ± 0.3 cd | 6.4 ± 0.1 b | 67.3 |
| 20 | 37.6 ± 0.7 c | 20.0 ± 4.0 bc | 22.6 ± 1.9 bc | 6.0 ± 0.1 c | 69.6 |
| 40 | 32.1 ± 2.4 d | 27.8 ± 1.8 ab | 31.6 ± 1.1 b | 5.9 ± 0.2 cd | 90.2 |
| 60 | 28.0 ± 0.9 d | 39.0 ± 9.4 a | 43.6 ± 7.2 a | 5.8 ± 0.1 d | 91.6 |
| Factor | Coefficient | SE Coefficient | F-Value | p-Value |
|---|---|---|---|---|
| Hempseed protein concentration | ||||
| pH | 0.6425 | 0.0434 | 219.54 | <0.0001 |
| Time | 0.5600 | 0.0434 | 166.78 | <0.0001 |
| Temperature | 0.4150 | 0.0434 | 91.59 | <0.0001 |
| pH × Time | 0.2175 | 0.0613 | 12.58 | 0.0094 |
| pH × Temp | 0.0175 | 0.0613 | 0.0814 | 0.7836 |
| Time × Temp | 0.0625 | 0.0613 | 1.04 | 0.3420 |
| pH × pH | −0.2038 | 0.0598 | 11.62 | 0.0113 |
| Time × Time | −0.4438 | 0.0598 | 55.12 | 0.0001 |
| Temp × Temp | −0.2837 | 0.0598 | 22.54 | 0.0021 |
| Free sulfhydryl (-SH) content | ||||
| pH | −7.60 | 0.5736 | 175.56 | <0.0001 |
| Time | −14.25 | 0.5736 | 617.10 | <0.0001 |
| Temperature | −9.19 | 0.5736 | 256.49 | <0.0001 |
| pH × Time | −2.72 | 0.8112 | 11.24 | 0.0122 |
| pH × Temp | −2.09 | 0.8112 | 6.61 | 0.0370 |
| Time × Temp | −1.35 | 0.8112 | 2.76 | 0.1406 |
| pH × pH | 6.71 | 0.7906 | 72.00 | <0.0001 |
| Time × Time | 12.89 | 0.7906 | 265.64 | <0.0001 |
| Temp × Temp | 6.21 | 0.7906 | 61.62 | 0.0001 |
| Selected Functionality | Original HPI | Optimised HPI |
|---|---|---|
| PS at pH 4.0 (%) | 11.1 ± 0.5 | 18.5 ± 3.8 |
| PS at pH 7.0 (%) | 29.4 ± 2.0 | 39.8 ± 4.1 |
| EAI (m2/g) | 5.8 ± 0.1 | 7.4 ± 1.2 |
| CI (%) | 91.6 | 12.9 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yao, S.; Li, W.; Martin, G.J.O.; Ashokkumar, M. An Investigation into the Mechanism of Alkaline Extraction-Isoelectric Point Precipitation (AE-IEP) of High-Thiol Plant Proteins. Appl. Sci. 2023, 13, 6469. https://doi.org/10.3390/app13116469
Yao S, Li W, Martin GJO, Ashokkumar M. An Investigation into the Mechanism of Alkaline Extraction-Isoelectric Point Precipitation (AE-IEP) of High-Thiol Plant Proteins. Applied Sciences. 2023; 13(11):6469. https://doi.org/10.3390/app13116469
Chicago/Turabian StyleYao, Shunyu, Wu Li, Gregory J. O. Martin, and Muthupandian Ashokkumar. 2023. "An Investigation into the Mechanism of Alkaline Extraction-Isoelectric Point Precipitation (AE-IEP) of High-Thiol Plant Proteins" Applied Sciences 13, no. 11: 6469. https://doi.org/10.3390/app13116469