
Genetic Algorithms for Optimal Control of Lactic Fermentation: Modelling the Lactobacillus paracasei CBA L74 Growth on Rice Flour Substrate

 , , , , ,
, , , , ,
Abstract
:1. Introduction
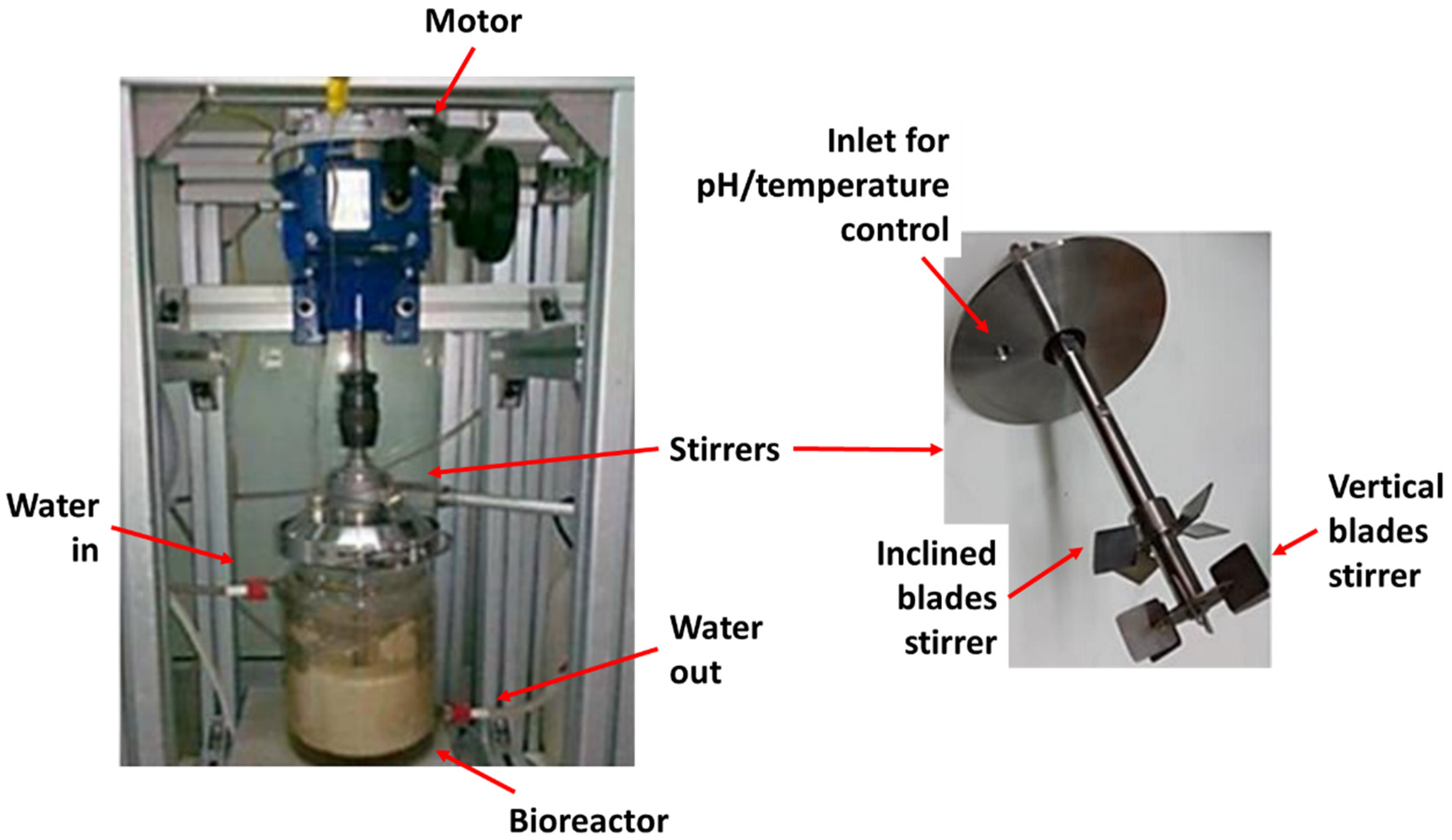
2. Materials and Methods
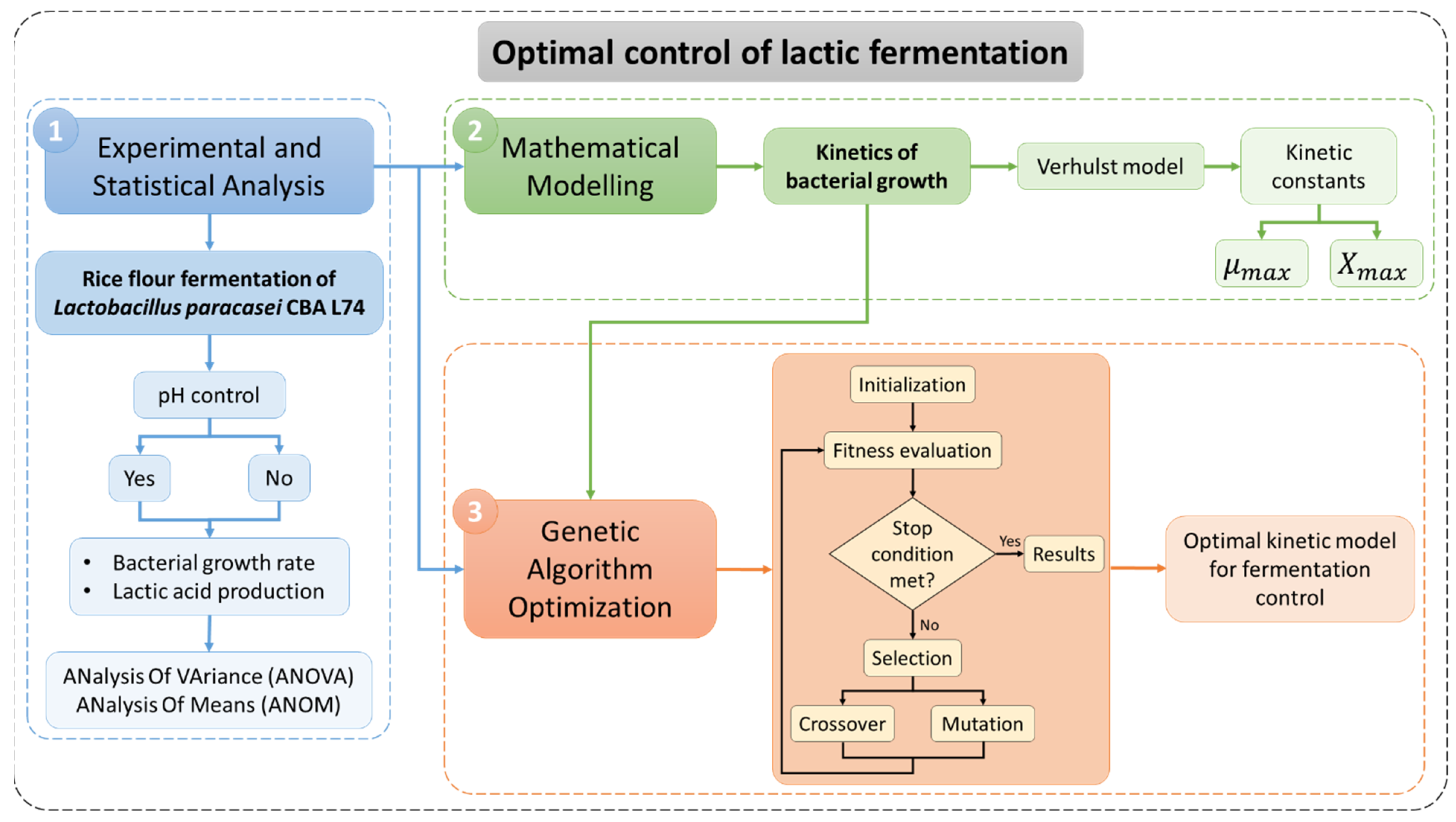
- The first step concerned the experimental and statistical investigation of the bacterial growth and subsequent lactic acid production of the Lactobacillus paracasei CBA L74 during rice flour fermentation by assessing the effect of the pH control through statistical tools as ANalysis Of VAriance (ANOVA) and ANalysis Of Means (ANOM). This phase is also aimed at creating the experimental data set to be used for the development of the optimal models during the following steps;
- The second step dealt with the definition of the kinetics of the bacterial growth by using the Verhulst non-structured mathematical model, which relates the specific growth rate as a function of the biomass concentration (only) during the exponential phase;
- Finally, a genetic algorithm-based approach is proposed with the aim of defining the optimal empirical model able to reproduce the kinetics of the bacterial growth based on the experimental data and to overcome the limitations of the conventional non-structured mathematical models that are generally able to accurately reproduce only the exponential phase.
2.1. Experimental and Statistical Analysis
2.2. Mathematical Modelling
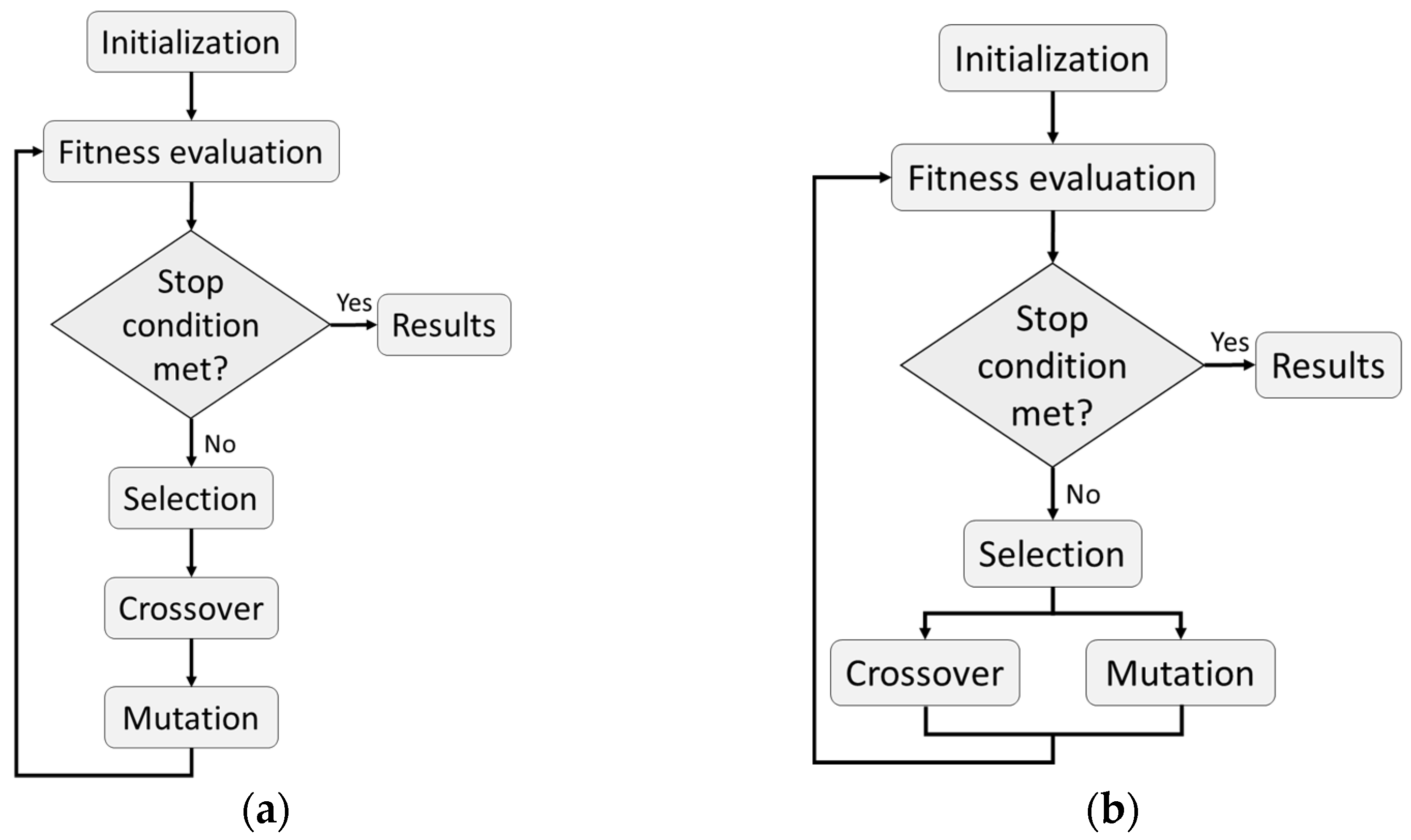
2.3. Genetic Algorithm Optimization
3. Results and Discussion
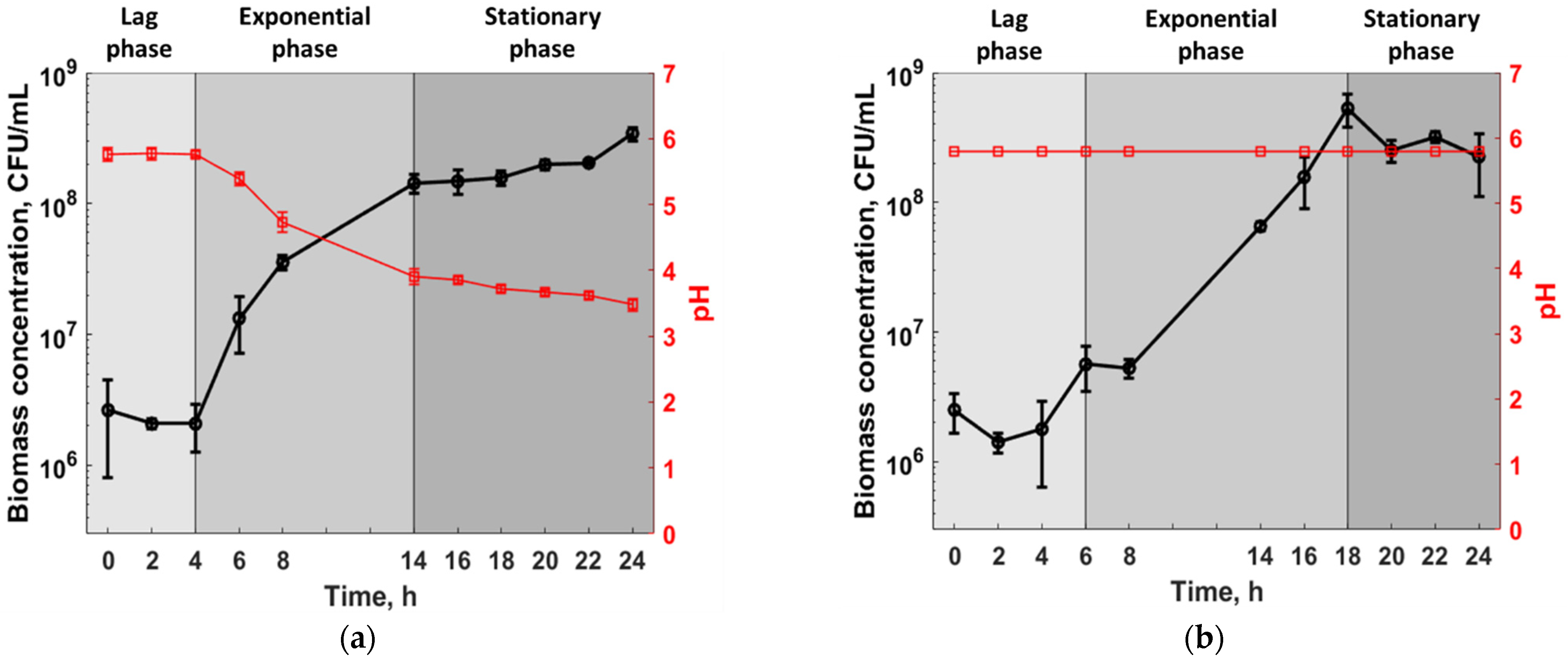
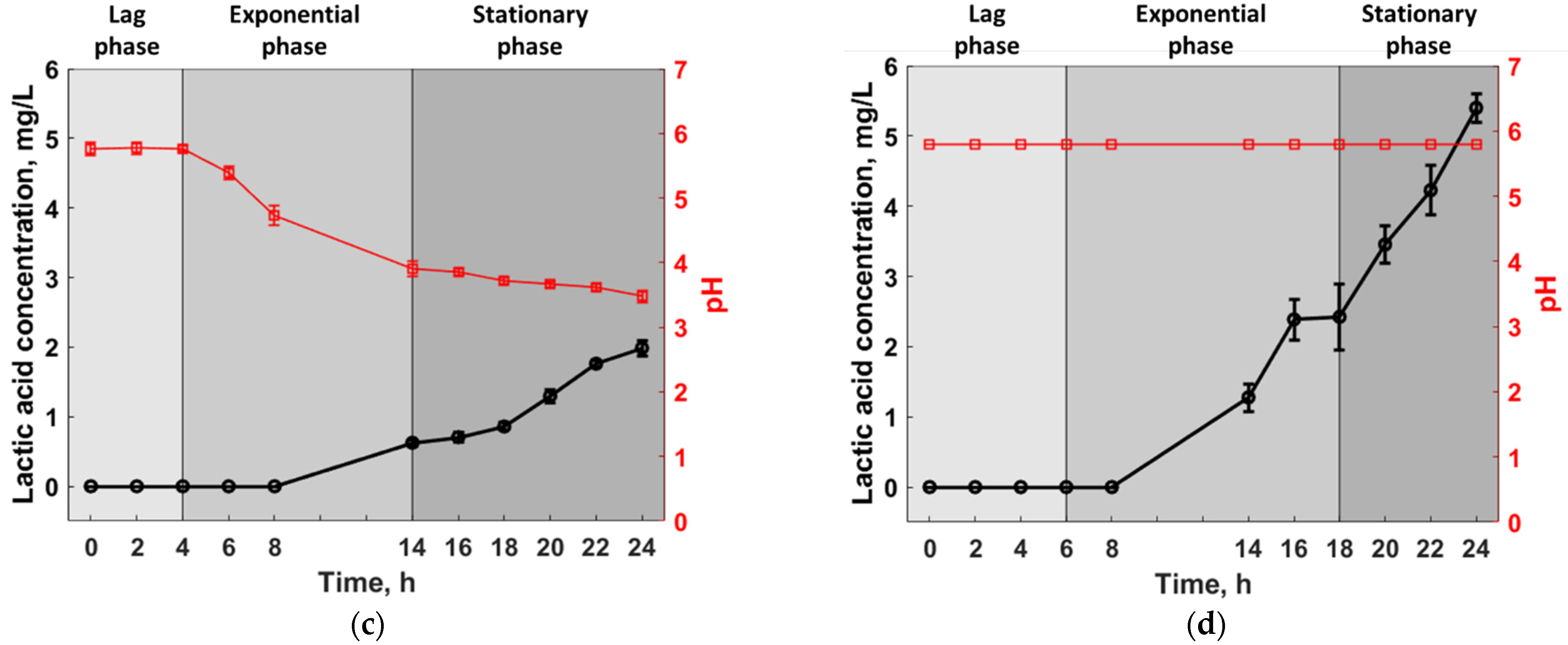
3.1. Experimental Analysis
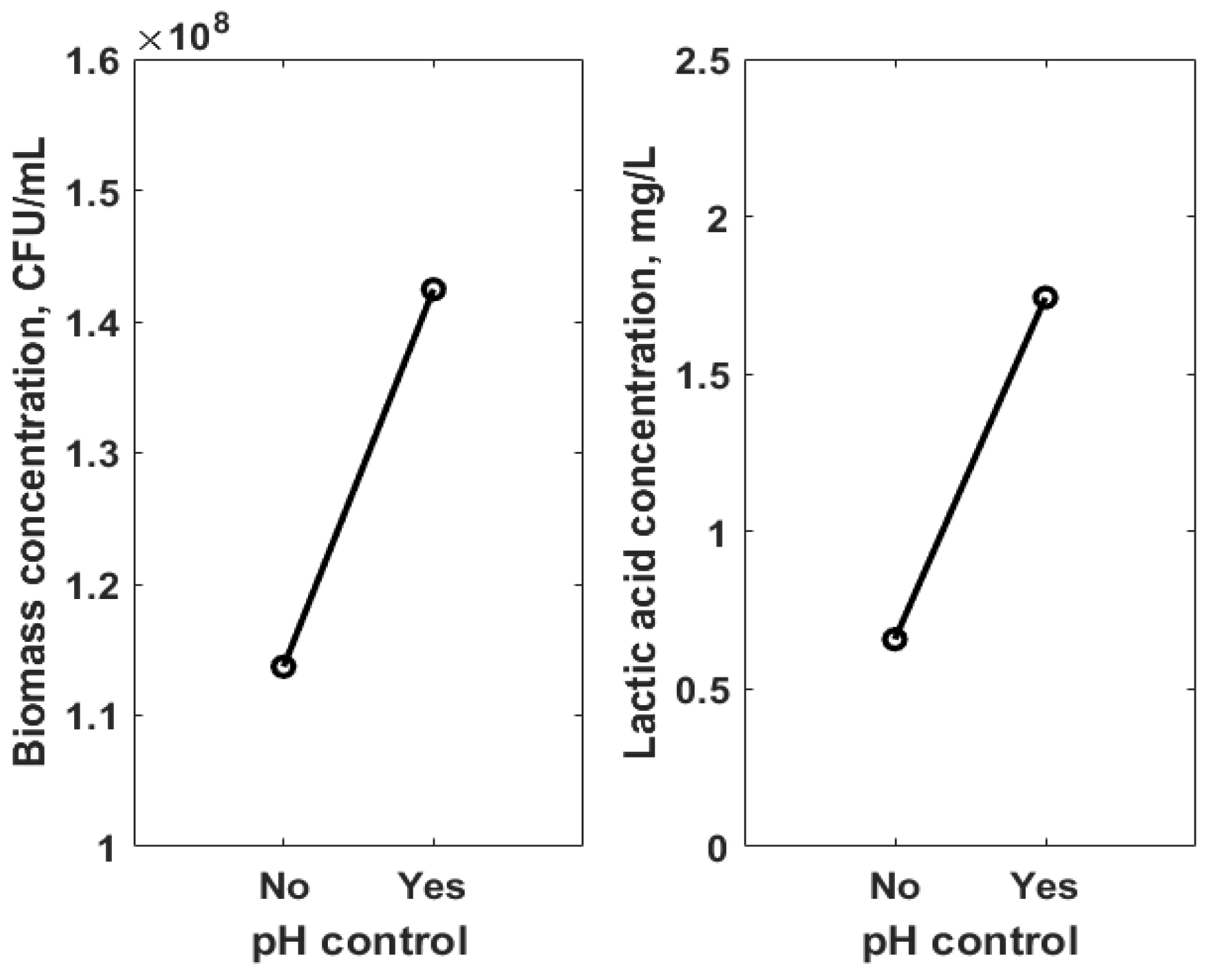
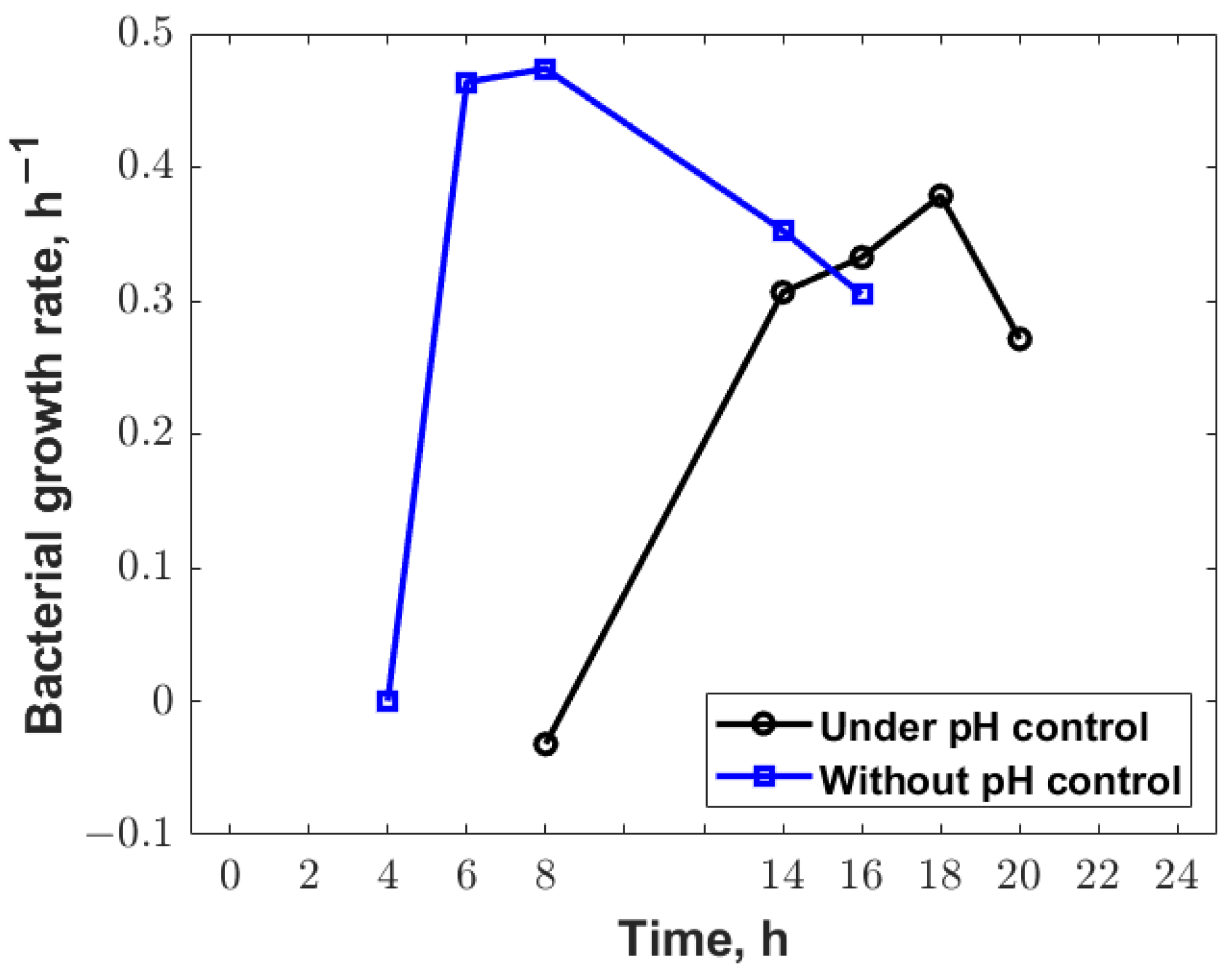
3.2. Statistical Analysis
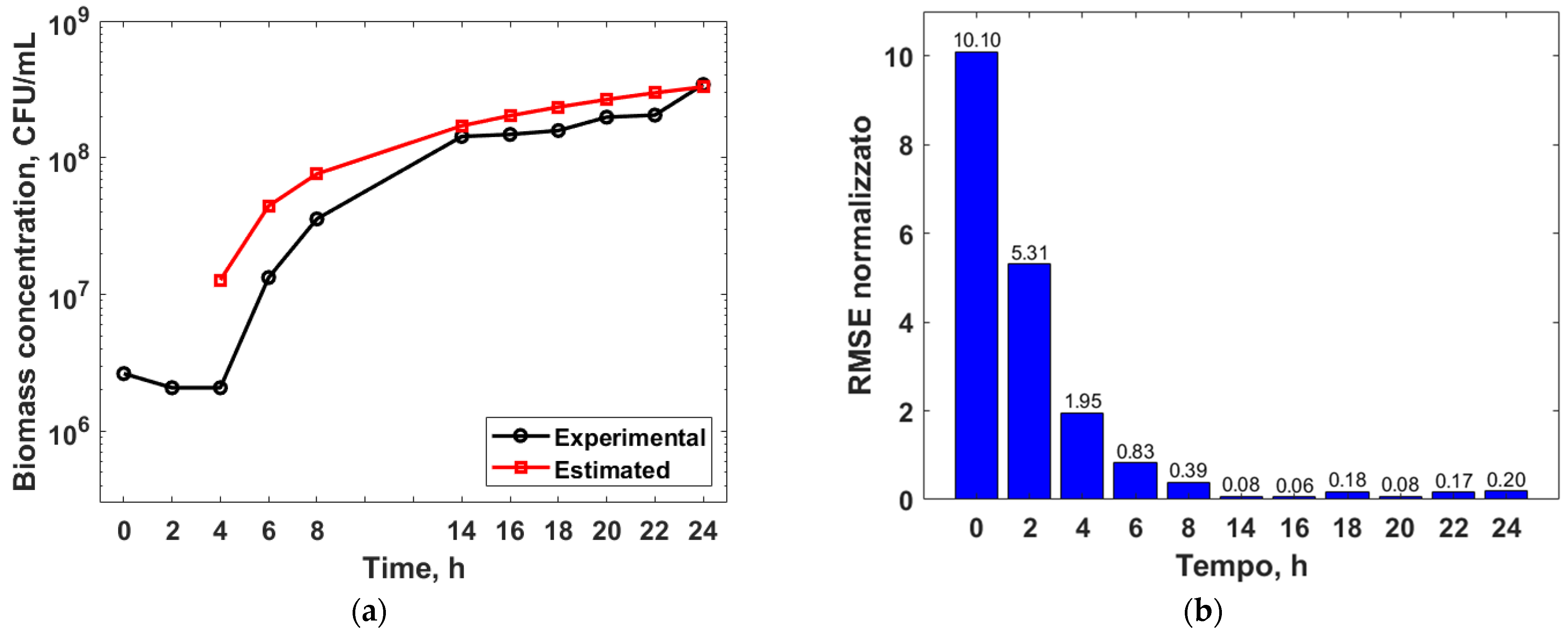
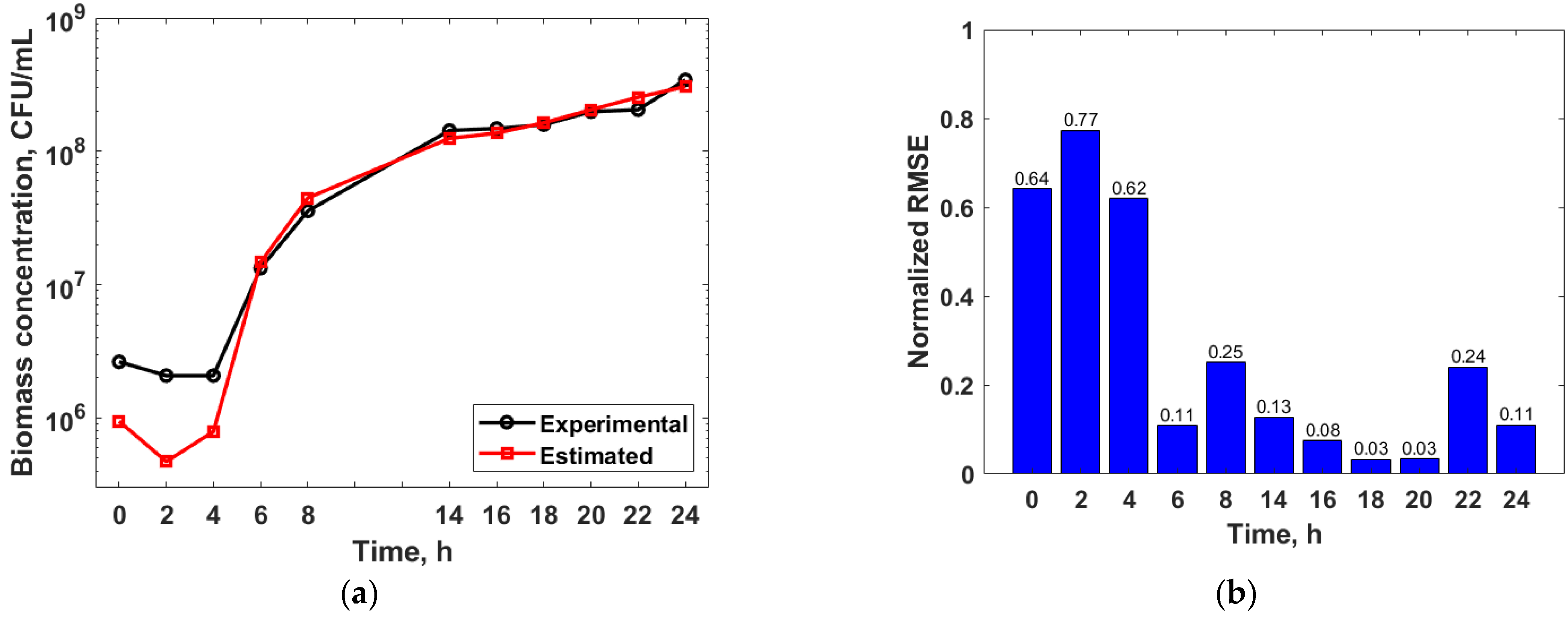
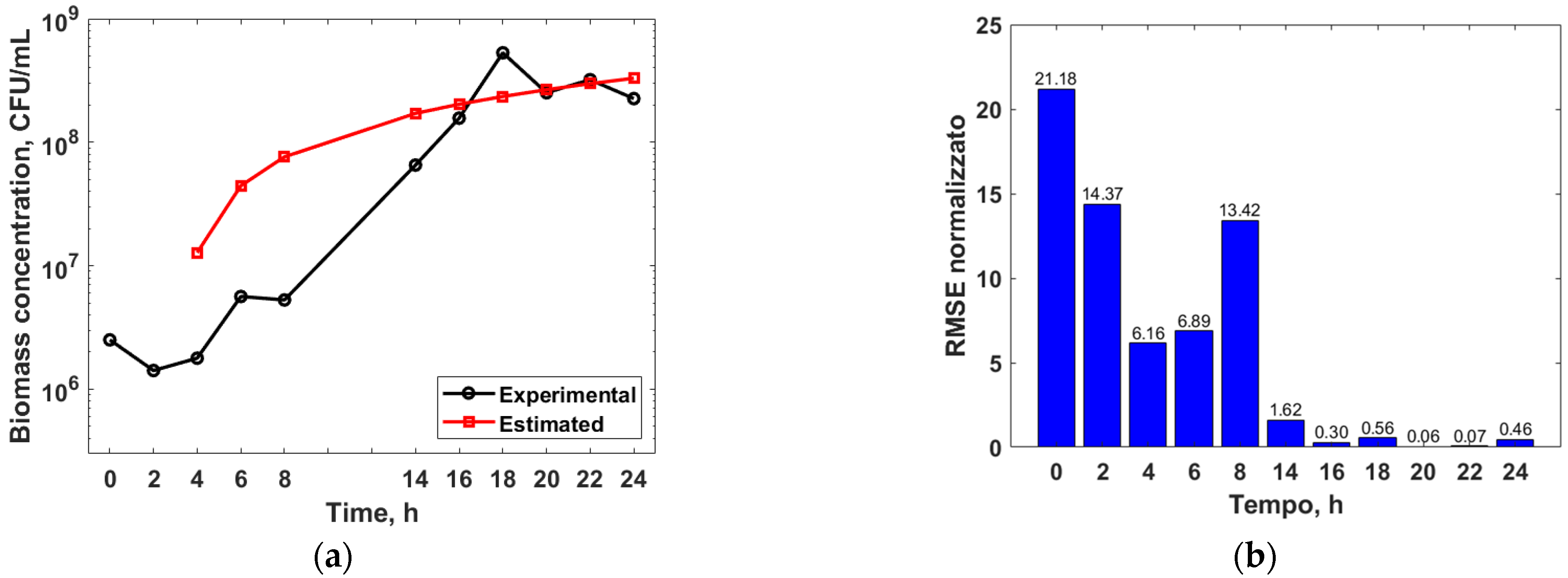
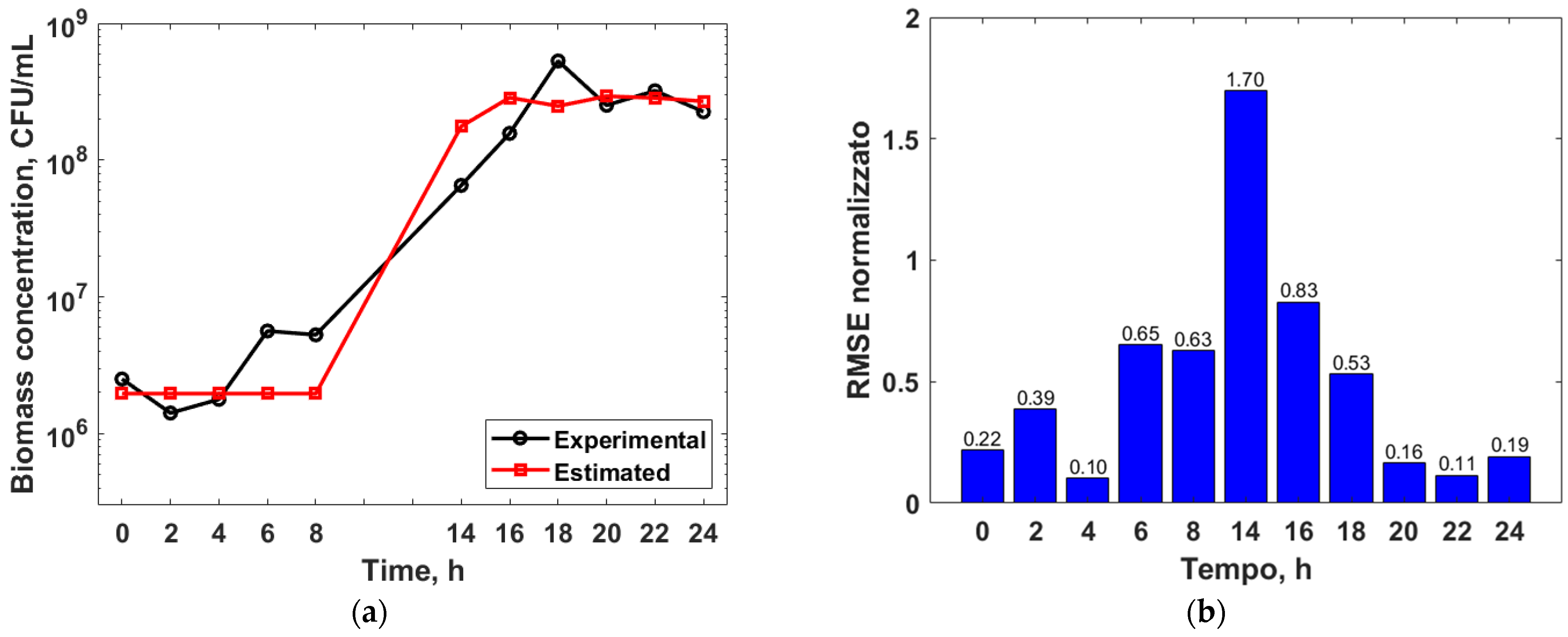
3.3. Mathematical Modelling
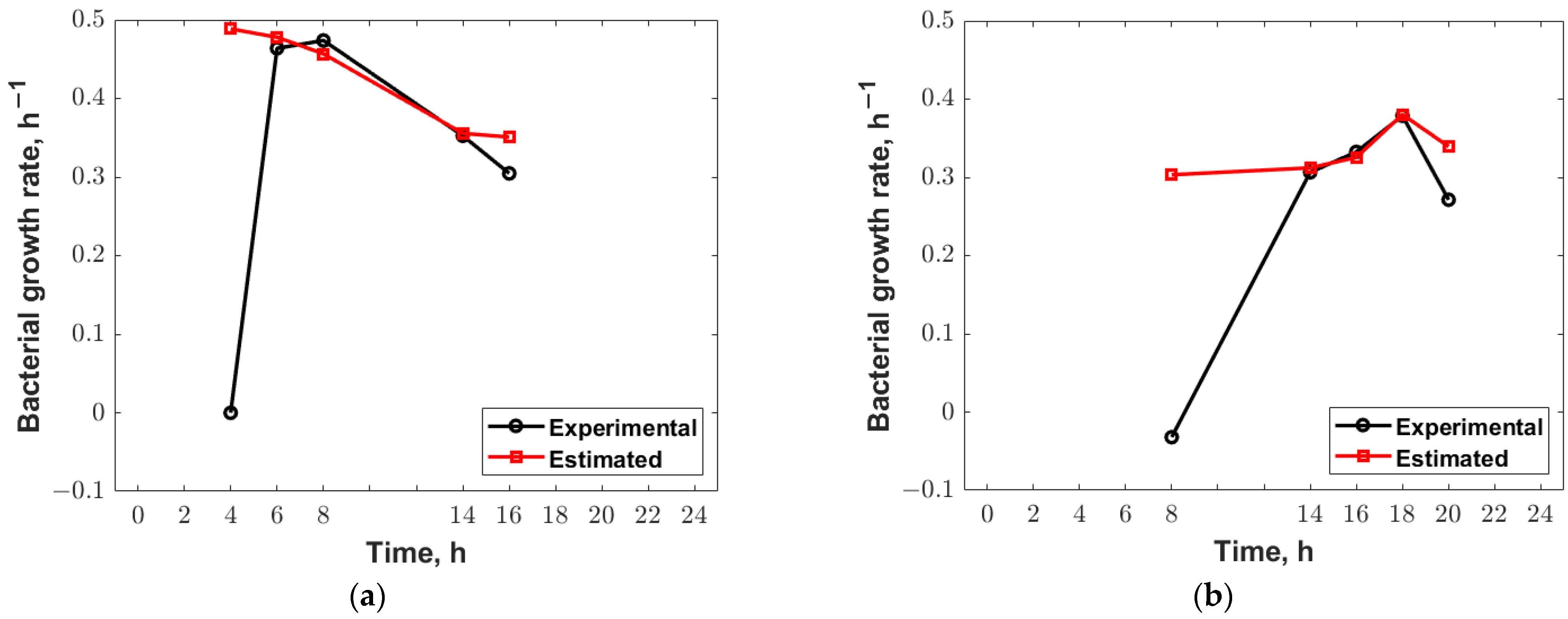
3.4. Genetic Algorithm Optimization
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Barros, C.P.; Guimarães, J.T.; Esmerino, E.A.; Duarte, M.C.K.; Silva, M.C.; Silva, R.; Ferreira, B.M.; Sant’Ana, A.S.; Freitas, M.Q.; Cruz, A.G. Paraprobiotics and Postbiotics: Concepts and Potential Applications in Dairy Products. Curr. Opin. Food Sci. 2020, 32, 1–8. [Google Scholar] [CrossRef]
- Bundele, J.; Deshmukh, R. Probiotics Market Growth, Size & Share|Global Report, 2030. 2021. Available online: https://www.alliedmarketresearch.com/probiotics-market (accessed on 8 March 2022).
- Gallo, M.; Passannanti, F.; Cante, R.C.; Nigro, F.; Salameh, D.; Schiattarella, P.; Schioppa, C.; Budelli, A.; Nigro, R. Effects of the Glucose Addition during Lactic Fermentation of Rice, Oat and Wheat Flours. Appl. Food Biotechnol. 2020, 7, 21–30. [Google Scholar] [CrossRef]
- Borah, T.; Gogoi, B.; Khataniar, A.; Gogoi, M.; Das, A.; Borah, D. Probiotic Characterization of Indigenous Bacillus Velezensis Strain DU14 Isolated from Apong, a Traditionally Fermented Rice Beer of Assam. Biocatal. Agric. Biotechnol. 2019, 18, 101008. [Google Scholar] [CrossRef]
- Salameh, D.; Nigro, F.; Cante, R.C.; Passannanti, F.; Gallo, M.; Budelli, A.; Marzocchella, A.; Nigro, R. Fermentation of Rice Flour Supernatant Using Lactobacillus Paracasei CBA L74. Chem. Eng. Trans. 2019, 75, 289–294. [Google Scholar] [CrossRef]
- Mättö, J.; Alakomi, H.L.; Vaari, A.; Virkajärvi, I.; Saarela, M. Influence of Processing Conditions on Bifidobacterium Animalis Subsp. Lactis Functionality with a Special Focus on Acid Tolerance and Factors Affecting It. Int. Dairy J. 2006, 16, 1029–1037. [Google Scholar] [CrossRef]
- Saarela, M.; Virkajärvi, I.; Alakomi, H.L.; Mattila-Sandholm, T.; Vaari, A.; Suomalainen, T.; Mättö, J. Influence of Fermentation Time, Cryoprotectant and Neutralization of Cell Concentrate on Freeze-Drying Survival, Storage Stability, and Acid and Bile Exposure of Bifidobacterium Animalis Ssp. Lactis Cells Produced without Milk-Based Ingredients. J. Appl. Microbiol. 2005, 99, 1330–1339. [Google Scholar] [CrossRef]
- Kanurić, K.G.; Hrnjez, D.v.; Ranogajec, M.I.; Milanović, S.D.; Iliĉić, M.D.; Vukić, V.R.; Milanović, M.L. The Effect of Fermentation Temperature on the Functional Dairy Product Quality. Acta Period. Technol. 2011, 42, 63–70. [Google Scholar] [CrossRef]
- Ross, R.P.; Hill, D.; Sugrue, I.; Arendt, E.; Hill, C.; Stanton, C. Recent Advances in Microbial Fermentation for Dairy and Health. F1000Research 2017, 6, 751. [Google Scholar]
- Gallo, M.; Nigro, F.; Passannanti, F.; Dana, S.; Schiattarella, P.; Budelli, A.; Nigro, R. Lactic Fermentation of Cereal Flour: Feasibility Tests on Rice, Oat and Wheat. Appl. Food Biotechnol. 2019, 6, 165–172. [Google Scholar] [CrossRef]
- Min, M.; Bunt, C.R.; Mason, S.L.; Hussain, M.A. Non-Dairy Probiotic Food Products: An Emerging Group of Functional Foods. Crit. Rev. Food Sci. Nutr. 2019, 59, 2626–2641. [Google Scholar] [CrossRef]
- Gallo, M.; Passannanti, F.; Schiattarella, P.; Esposito, A.; Colucci Cante, R.; Nigro, F.; Budelli, A.; Nigro, R. Banana Puree Lactic Fermentation: The Role of Ripeness, Heat Treatment, and Ascorbic Acid. Appl. Sci. 2021, 11, 5153. [Google Scholar] [CrossRef]
- Prasad, J.; Gill, H.; Smart, J.; Gopal, P.K. Selection and Characterisation of Lactobacillus and Bifidobacterium Strains for Use as Probiotics. Int. Dairy J. 1998, 8, 993–1002. [Google Scholar] [CrossRef]
- Cante, R.C.; Gallo, M.; Nigro, F.; Passannanti, F.; Budelli, A.; Nigro, R. Mathematical Modeling of Lactobacillus Paracasei Cba L74 Growth during Rice Flour Fermentation Performed with and without Ph Control. Appl. Sci. 2021, 11, 2921. [Google Scholar] [CrossRef]
- Ghimire, A.; Kumar Sah, A.; Poudel, R. Kinetics and Modeling of Growth and Lactic Acid Production in Gundruk, a Himalayan Fermented Vegetable Dish. Food Sci. Nutr. 2020, 8, 5591. [Google Scholar] [CrossRef] [PubMed]
- Rezvani, F.; Ardestani, F.; Najafpour, G. Growth Kinetic Models of Five Species of Lactobacilli and Lactose Consumption in Batch Submerged Culture. Braz. J. Microbiol. 2017, 48, 251. [Google Scholar] [CrossRef] [PubMed]
- Monod, J. The Growth of Bacterial Cultures. Annu. Rev. Microbiol. 1949, 3, 371–394. [Google Scholar] [CrossRef] [Green Version]
- Muloiwa, M.; Nyende-Byakika, S.; Dinka, M. Comparison of Unstructured Kinetic Bacterial Growth Models. S. Afr J. Chem. Eng. 2020, 33, 141–150. [Google Scholar] [CrossRef]
- Borojeni, M.S.; Shahmansourian, A. Genetic Algorithm Is a More Accurate Approach to Estimate Maximum Growth Rate of Bacteria Growth Pattern. In Proceedings of the 2nd International Conference on Mathematical Sciences (ICMS 2014), Istanbul, Turkey, 9–11 July 2014. [Google Scholar]
- Pedrozo, H.A.; Dallagnol, A.M.; Schvezov, C.E. Genetic Algorithm Applied to Simultaneous Parameter Estimation in Bacterial Growth. J. Bioinform. Comput. Biol. 2021, 19, 2050045. [Google Scholar] [CrossRef]
- García-Gimeno, R.M.; Hervás-Martínez, C.; de Silóniz, M.I. Improving Artificial Neural Networks with a Pruning Methodology and Genetic Algorithms for Their Application in Microbial Growth Prediction in Food. Int. J. Food Microbiol. 2002, 72, 19–30. [Google Scholar] [CrossRef]
- Gallo, M.; Nigro, F.; Passannanti, F.; Nanayakkara, M.; Lania, G.; Parisi, F.; Salameh, D.; Budelli, A.; Barone, M.V.; Nigro, R. Effect of PH Control during Rice Fermentation in Preventing a Gliadin P31-43 Entrance in Epithelial Cells. Int. J. Food Sci. Nutr. 2019, 70, 950–958. [Google Scholar] [CrossRef]
- Colucci Cante, R.; Gallo, M.; Nigro, F.; Passannanti, F.; Salameh, D.; Budelli, A.; Nigro, R. Lactic Fermentation of Cooked Navy Beans by Lactobacillus Paracasei CBA L74 Aimed at a Potential Production of Functional Legume-Based Foods. Can. J. Chem. Eng. 2020, 98, 1955–1961. [Google Scholar] [CrossRef]
- Gallo, M.; Passannanti, F.; Colucci Cante, R.; Nigro, F.; Schiattarella, P.; Zappulla, S.; Budelli, A.; Nigro, R. Lactic Fermentation of Cereals Aqueous Mixture of Oat and Rice Flours with and without Glucose Addition. Heliyon 2020, 6, e04920. [Google Scholar] [CrossRef] [PubMed]
- Hetényi, K.; Németh, Á.; Sevella, B. Role of PH-Regulation in Lactic Acid Fermentation: Second Steps in a Process Improvement. Chem. Eng. Process. Process. Intensif. 2011, 50, 293–299. [Google Scholar] [CrossRef]
- Miller, R.G.; Brown, B.W. Beyond ANOVA: Basics of Applied Statistics, 1st ed.; Chapman & Hall/CRC: Boca Raton, FL, USA, 1997; ISBN 9780412070112. [Google Scholar]
- Pallmann, P. Analysis of Means: Examples Using Package ANOM. 2016. Available online: https://cran.microsoft.com/snapshot/2015-09-02/web/packages/ANOM/vignettes/ANOM.pdf (accessed on 8 March 2022).
- Mohr, D.L.; Wilson, W.J.; Freund, R.J. Probability and Sampling Distributions. In Statistical Methods; Elsevier: Amsterdam, The Netherlands, 2022; pp. 65–122. ISBN 978-0-12-823043-5. [Google Scholar]
- Ramirez-Cando, L.J.; Alvarez-Mendoza, C.I. Verhulst-Pearl Growth Model versus Malthusian Growth Model for In Vitro Evaluation of Lead Removal in Wastewater by Photobacterium Sp. F1000Research 2018, 7, 491. [Google Scholar] [CrossRef] [Green Version]
- Dao, S.D.; Abhary, K.; Marian, R. A Bibliometric Analysis of Genetic Algorithms throughout the History. Comput. Ind. Eng. 2017, 110, 395–403. [Google Scholar] [CrossRef]
- Aguirre, H.; Tanaka, K. A Model for Parallel Operators in Genetic Algorithms. In Parallel Evolutionary Computations; Springer: Berlin/Heidelberg, Germany, 2006; pp. 3–31. [Google Scholar]
- Genna, S.; Giannini, O.; Guarino, S.; Ponticelli, G.S.; Tagliaferri, F. Laser Texturing of AISI 304 Stainless Steel: Experimental Analysis and Genetic Algorithm Optimisation to Control the Surface Wettability. Int. J. Adv. Manuf. Technol. 2020, 110, 3005–3022. [Google Scholar] [CrossRef]
- Verotti, M.; Di Giamberardino, P.; Belfiore, N.P.; Giannini, O. A Genetic Algorithm-Based Method for the Mechanical Characterization of Biosamples Using a MEMS Microgripper: Numerical Simulations. J. Mech. Behav. Biomed. Mater. 2019, 96, 88–95. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Output | Time, h | Source | F-Value | p-Value | Π% |
|---|---|---|---|---|---|
| Bacterial growth rate | 2 | pH control | 14.434 | 0.019 | 78.29 |
| Error | 21.71 | ||||
| 8 | pH control | 131.630 | 0.000 | 97.20 | |
| Error | 2.80 | ||||
| 14 | pH control | 33.741 | 0.004 | 89.60 | |
| Error | 10.40 | ||||
| 18 | pH control | 18.758 | 0.012 | 82.54 | |
| Error | 17.46 | ||||
| 22 | pH control | 42.854 | 0.003 | 91.40 | |
| Error | 8.60 | ||||
| Lactic acid concentration | 14 | pH control | 31.199 | 0.005 | 88.56 |
| Error | 11.44 | ||||
| 16 | pH control | 94.330 | 0.001 | 95.93 | |
| Error | 4.07 | ||||
| 18 | pH control | 33.142 | 0.005 | 89.23 | |
| Error | 10.77 | ||||
| 20 | pH control | 178.937 | 0.000 | 97.81 | |
| Error | 2.19 | ||||
| 22 | pH control | 144.268 | 0.000 | 97.31 | |
| Error | 2.69 | ||||
| 24 | pH control | 671.710 | 0.000 | 99.41 | |
| Error | 0.59 |
| Kinetic Constants | Without pH Control | Under pH Control | ||
|---|---|---|---|---|
| Experimental | Estimated | Experimental | Estimated | |
| , h−1 | 0.4738 | 0.4905 | 0.3787 | 0.3025 |
| , CFU/mL | 3.43 × 108 | 5.20 × 108 | 2.26 × 108 | 2.06 × 109 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ponticelli, G.S.; Gallo, M.; Cacciotti, I.; Giannini, O.; Guarino, S.; Budelli, A.; Nigro, R. Genetic Algorithms for Optimal Control of Lactic Fermentation: Modelling the Lactobacillus paracasei CBA L74 Growth on Rice Flour Substrate. Appl. Sci. 2023, 13, 582. https://doi.org/10.3390/app13010582
Ponticelli GS, Gallo M, Cacciotti I, Giannini O, Guarino S, Budelli A, Nigro R. Genetic Algorithms for Optimal Control of Lactic Fermentation: Modelling the Lactobacillus paracasei CBA L74 Growth on Rice Flour Substrate. Applied Sciences. 2023; 13(1):582. https://doi.org/10.3390/app13010582
Chicago/Turabian StylePonticelli, Gennaro Salvatore, Marianna Gallo, Ilaria Cacciotti, Oliviero Giannini, Stefano Guarino, Andrea Budelli, and Roberto Nigro. 2023. "Genetic Algorithms for Optimal Control of Lactic Fermentation: Modelling the Lactobacillus paracasei CBA L74 Growth on Rice Flour Substrate" Applied Sciences 13, no. 1: 582. https://doi.org/10.3390/app13010582