
Effect of Commercial Microbial Preparations Containing Paenibacillus azotofixans, Bacillus megaterium and Bacillus subtilis on the Yield and Photosynthesis of Winter Wheat and the Nitrogen and Phosphorus Content in the Soil


Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design, Growing Conditions and Treatments
2.2. Yield and Quality Analysis Samplings
2.3. Leaf Gas Exchange, SPAD Index Measurement
- (1)
- stem elongation–first node at least 1 cm above tillering node (BBCH31)
- (2)
- inflorescence emergence, heading–beginning of heading (BBCH51).
2.4. Physicochemical Soil Analyses
2.5. Statistical Analysis
3. Results and Discussion
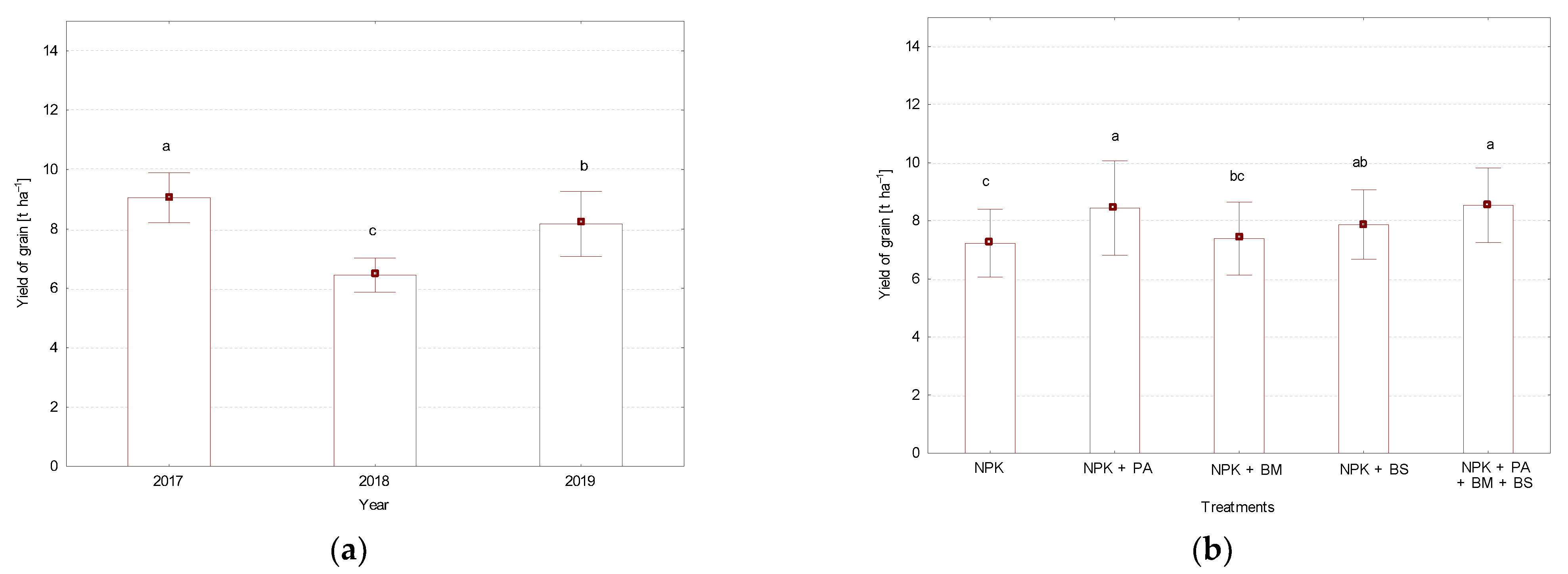
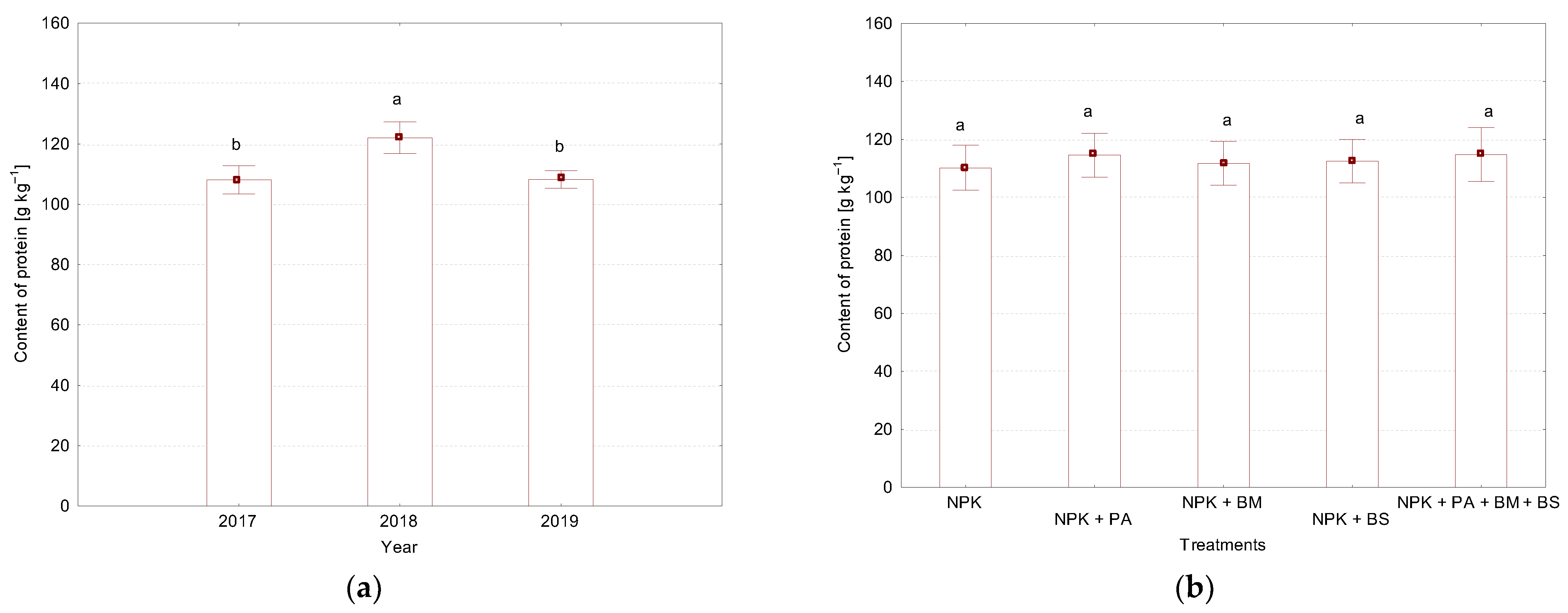
3.1. Grain Yield and Protein Content
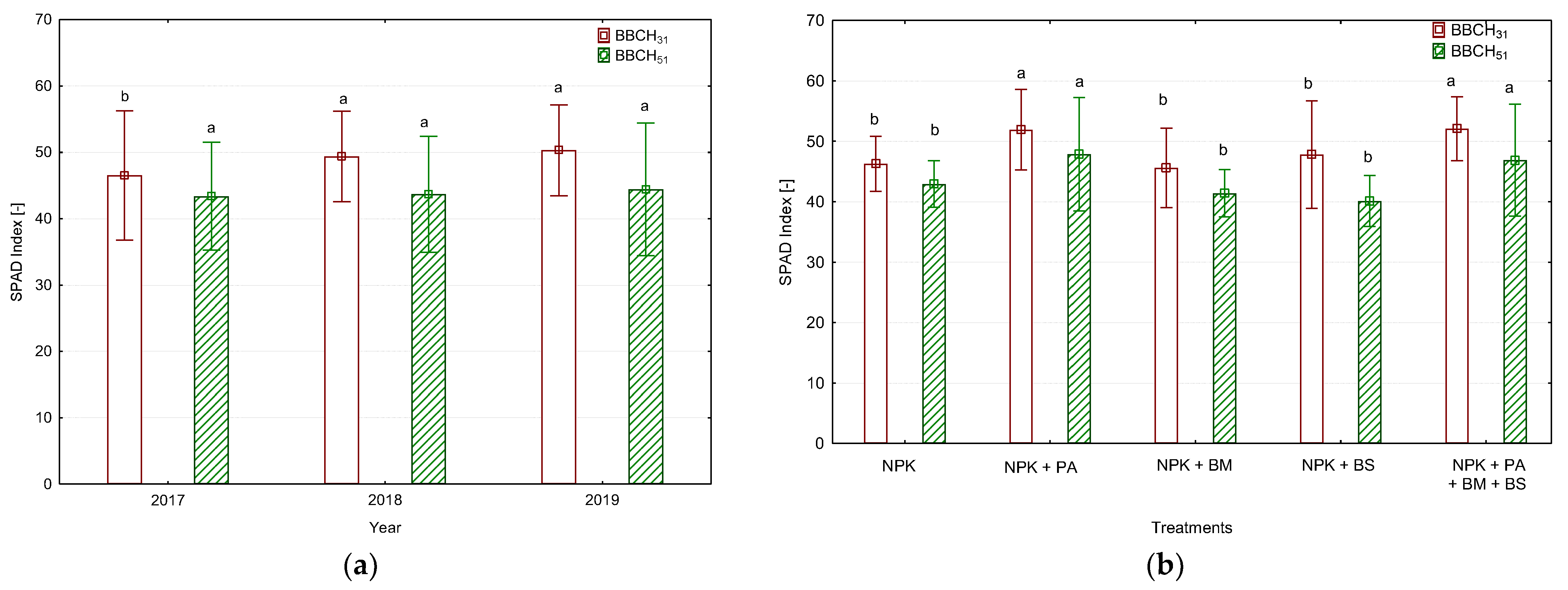
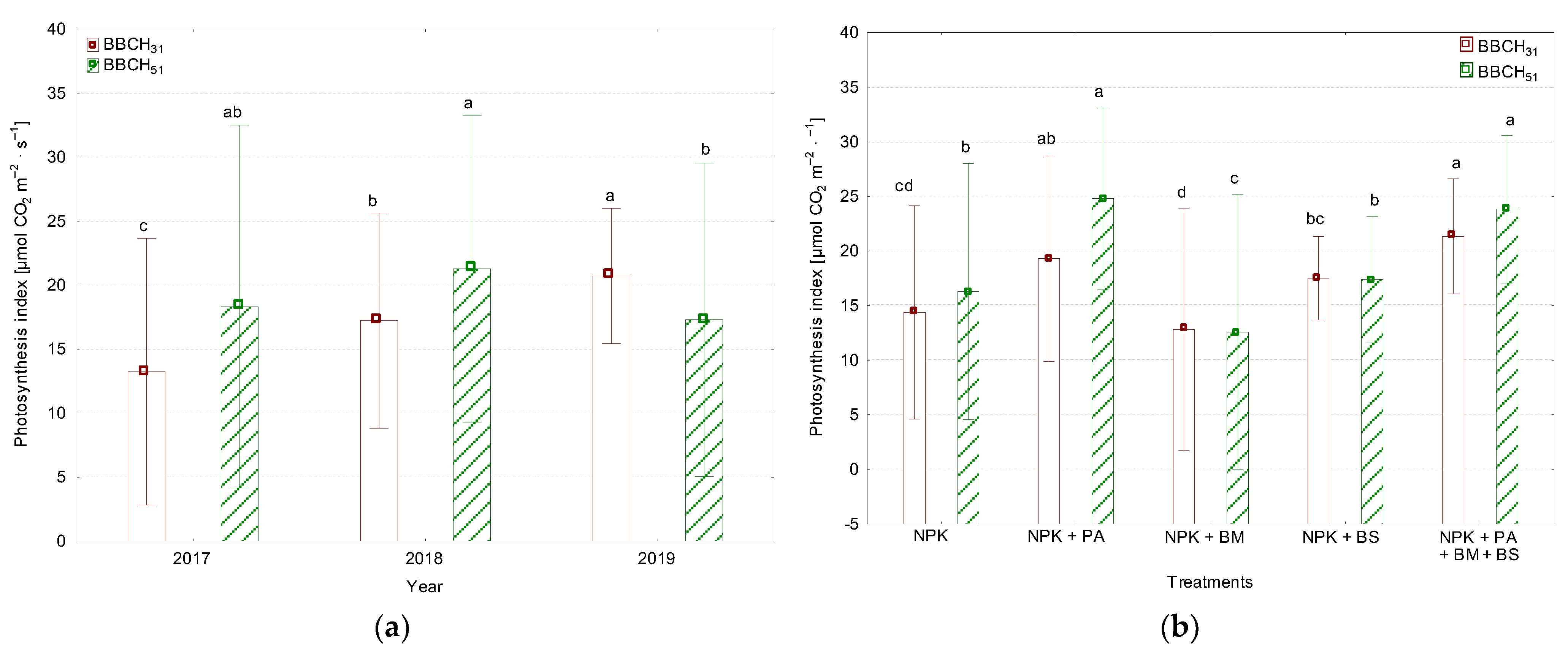
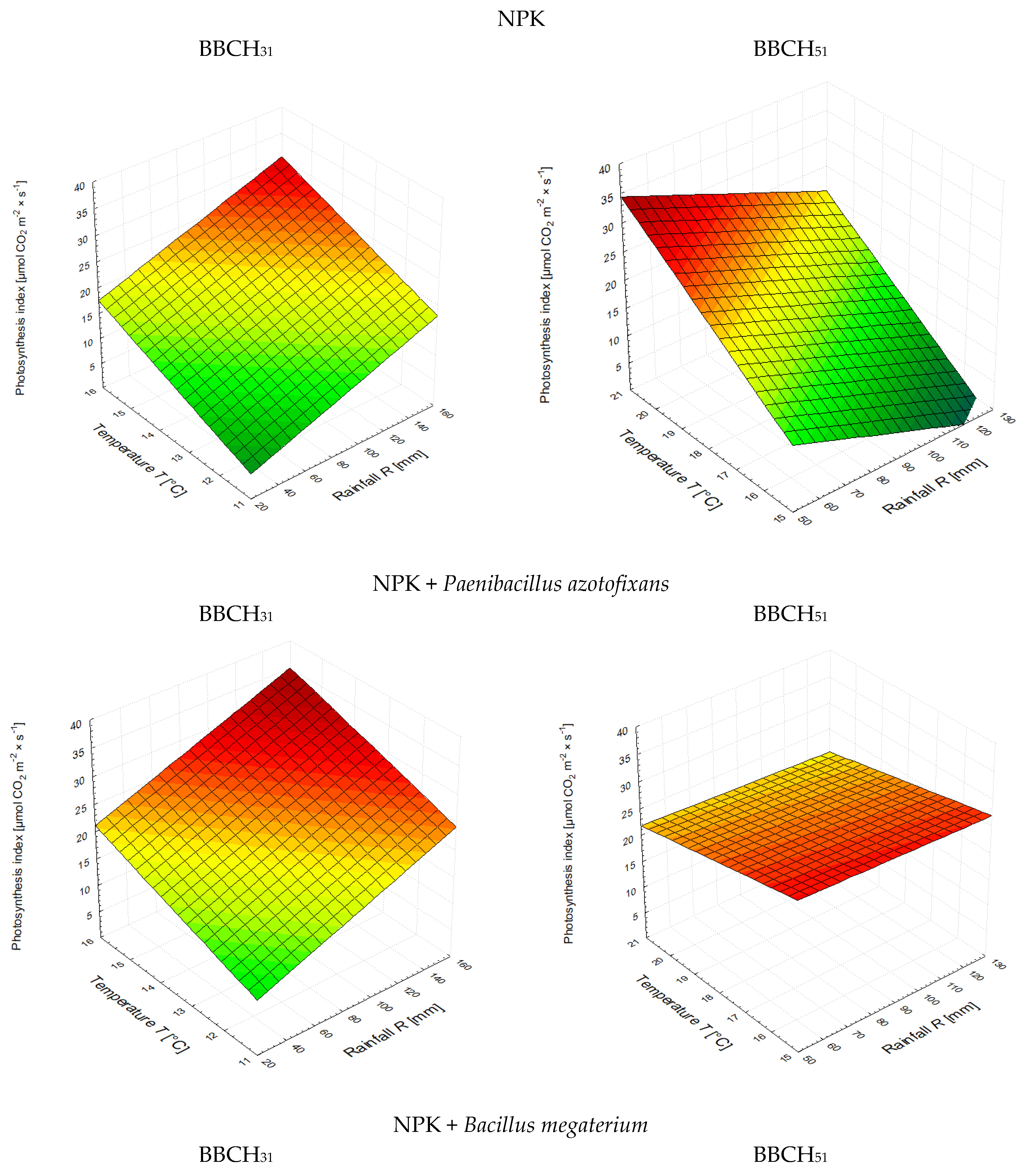
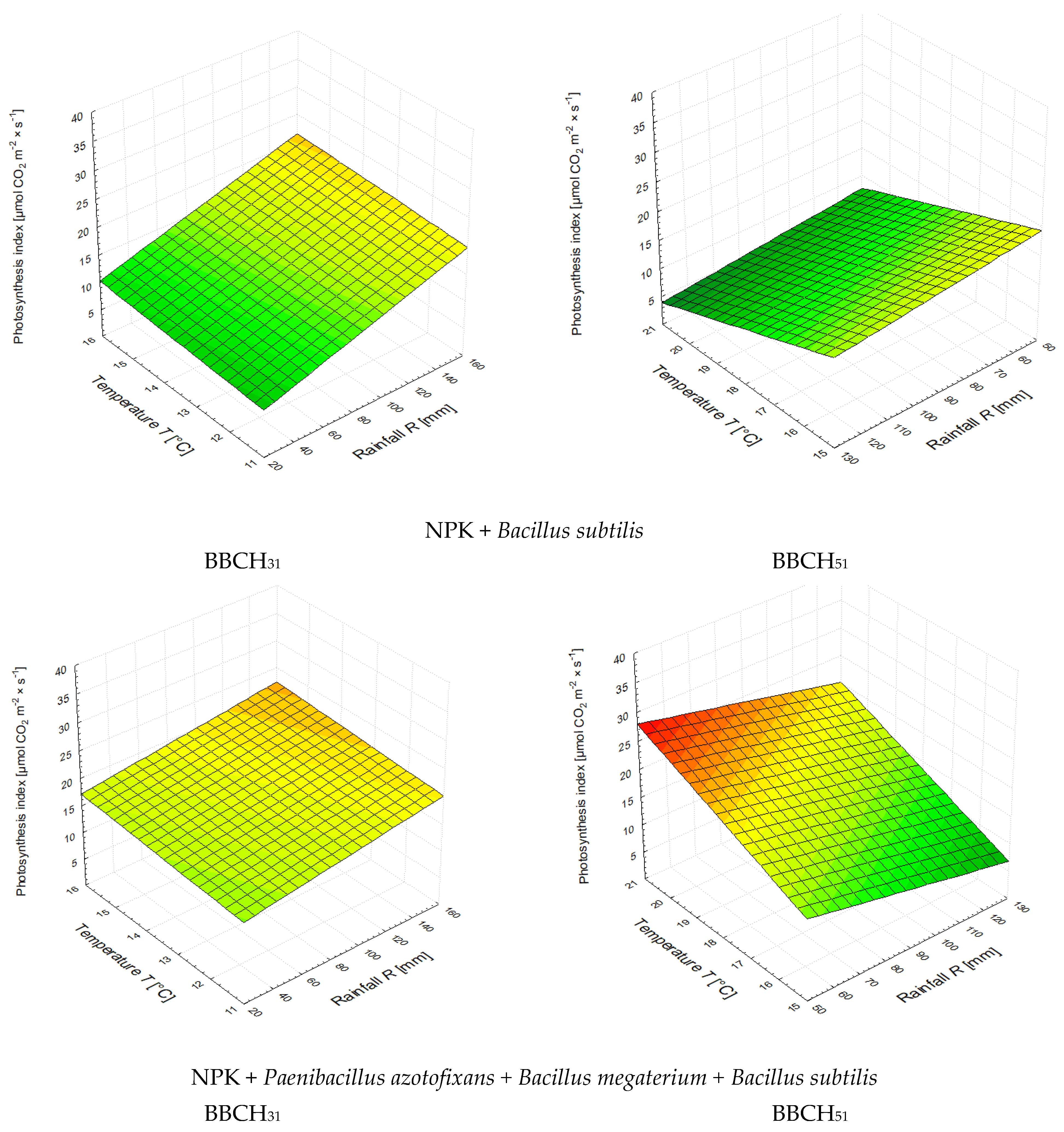
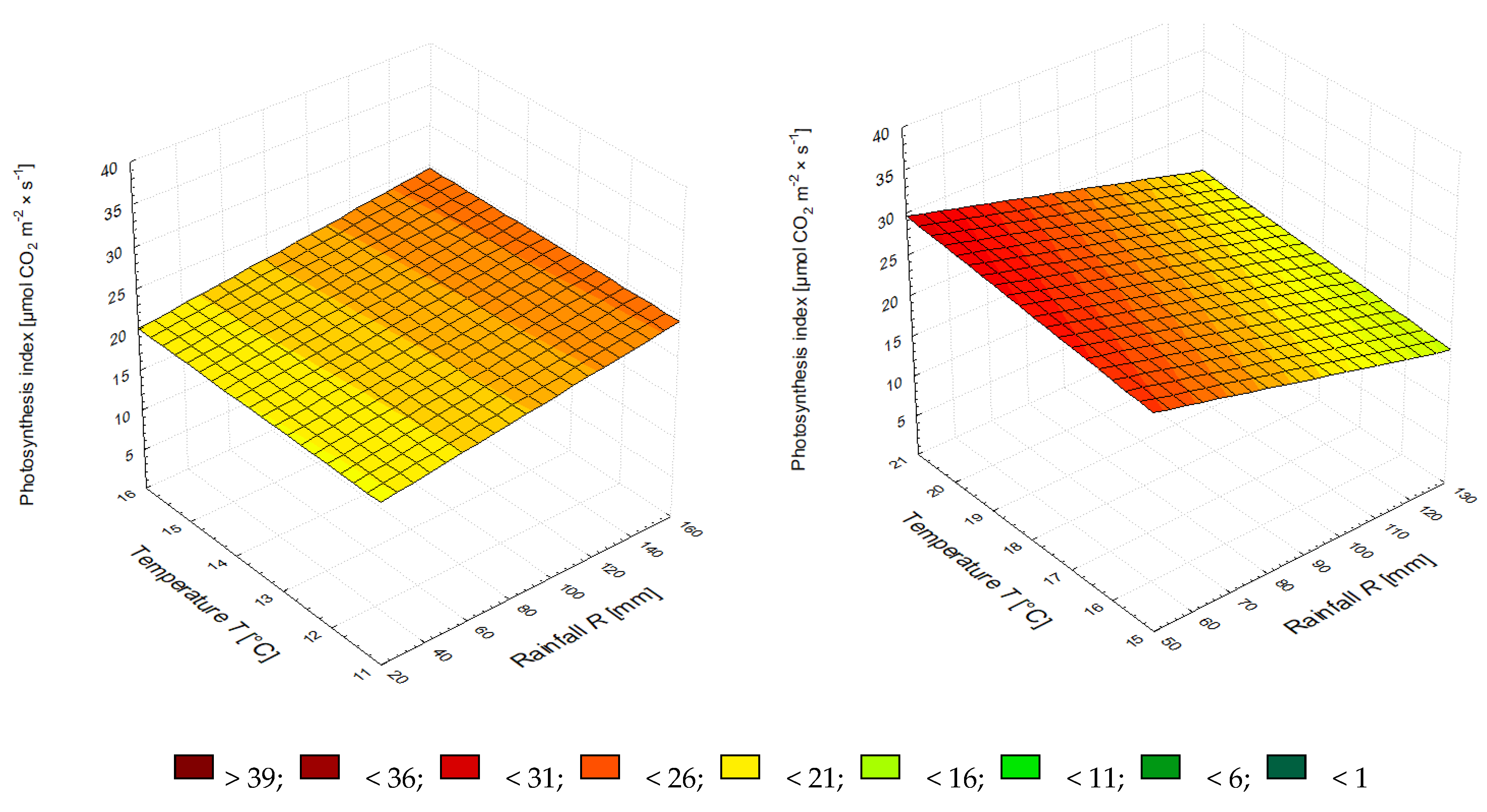
3.2. The Leaf Greenness Index (SPAD) and the Photosynthetic Rate
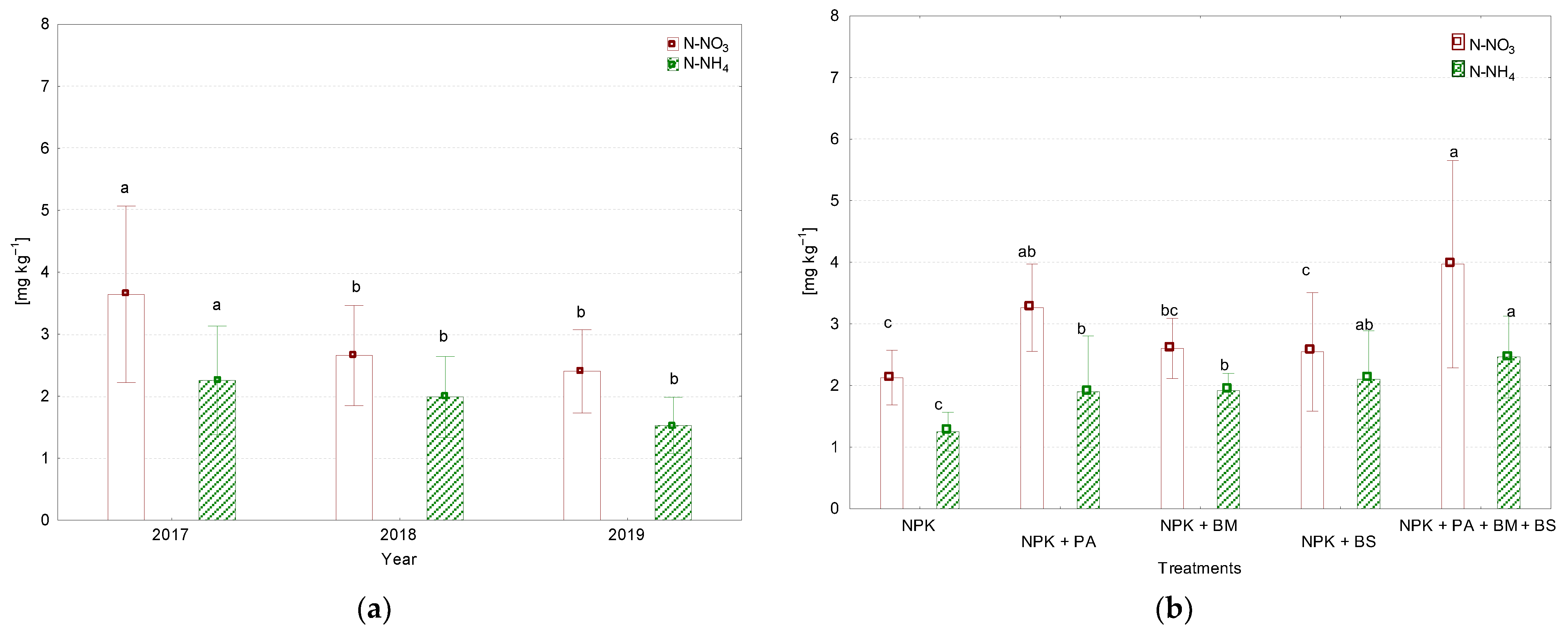
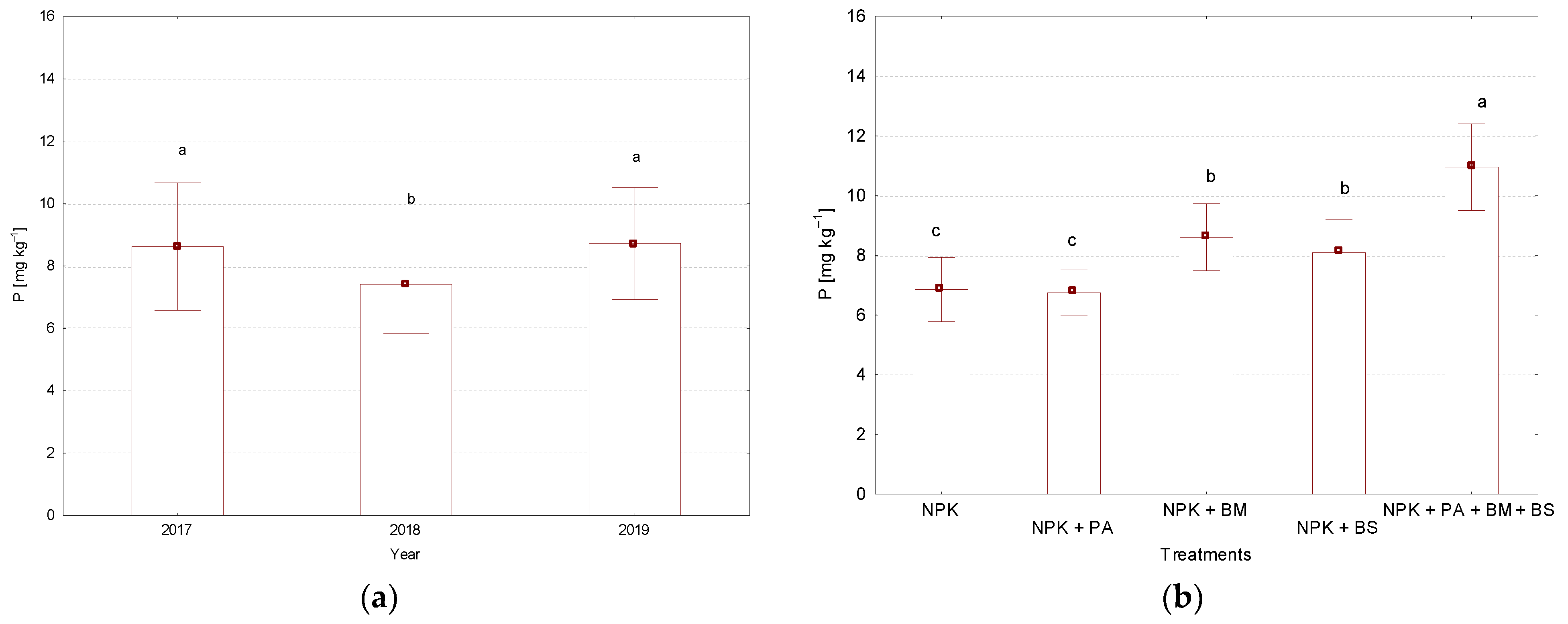
3.3. N-NO3,N-NH4, and P Contents in the Soil
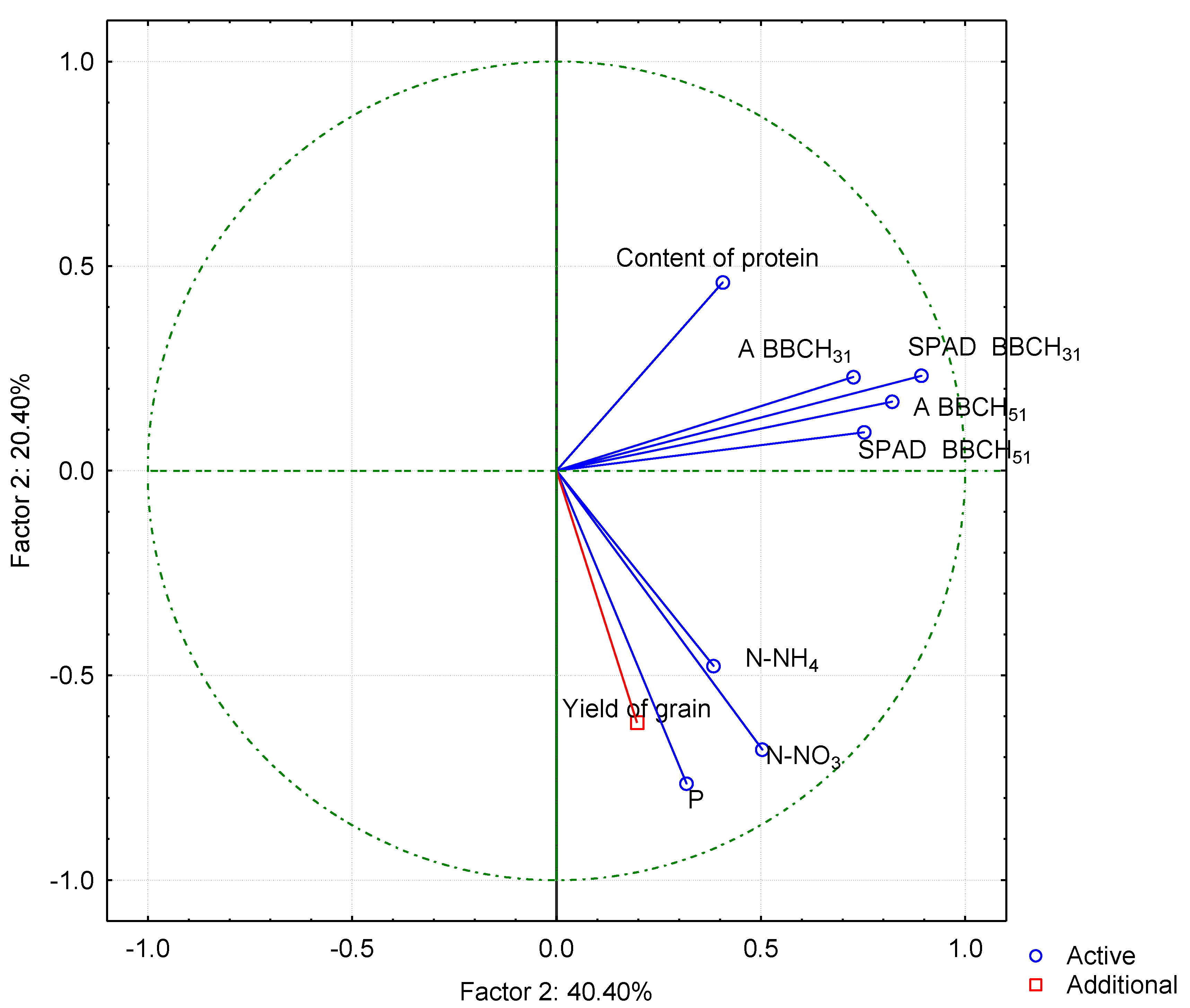
3.4. Principal Component Analysis–PCA
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Jaskulska, I.; Jaskulski, D.; Gałęzewski, L.; Knapowski, T.; Kozera, W.; Wacławowicz, R. Mineral Composition and Baking Value of the Winter Wheat Grain under Varied Environmental and Agronomic Conditions. J. Chem. 2018, 2018, 5013825. [Google Scholar] [CrossRef] [Green Version]
- Woźniak, A.; Rachoń, L. Effect of Tillage Systems on the Yield and Quality of Winter Wheat Grain and Soil Properties. Agriculture 2020, 10, 405. [Google Scholar] [CrossRef]
- Buczek, J.; Migut, D.; Jańczak-Pieniążek, M. Effect of Soil Tillage Practice on Photosynthesis, Grain Yield and Quality of Hybrid Winter Wheat. Agriculture 2021, 11, 479. [Google Scholar] [CrossRef]
- Ju, X.-T.; Xing, G.-X.; Chen, X.-P.; Zhang, S.-L.; Zhang, L.-J.; Liu, X.-J.; Cui, Z.-L.; Yin, B.; Christie, P.; Zhu, Z.-L.; et al. Reducing environmental risk by improving N management in intensive Chinese agricultural systems. Proc. Natl. Acad. Sci. USA 2009, 106, 3041–3046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kosicka, D.; Wolna-Maruwka, A.; Trzeciak, M. The influence of microbial inoculates on morphological traits in plants. Kosmos 2015, 64, 327–335. (In Polish) [Google Scholar]
- Bonanomi, G.; De Filippis, F.; Zotti, M.; Idbella, M.; Cesarano, G.; Al-Rowaily, S.; Abd-ElGawad, A. Repeated applications of organic amendments promote beneficial microbiota, improve soil fertility and increase crop yield. Appl. Soil Ecol. 2020, 156, 103714. [Google Scholar] [CrossRef]
- Itelima, J.U.; Bang, W.J.; Onyimba, I.A.; Sila, M.D.; Egbere, O.J. Bio-fertilizers as key player in enhancing soil fertility and crop productivity. Direct Res. J. Agric. Food Sci. 2018, 6, 73–83. [Google Scholar] [CrossRef]
- Mącik, M.; Gryta, A.; Frąc, M. Biofertilizers in agriculture: An overview on concepts, strategies and effects on soil microorganisms. Adv. Agron. 2020, 162, 31–87. [Google Scholar] [CrossRef]
- Basu, A.; Prasad, P.; Das, S.N.; Kalam, S.; Sayyed, R.Z.; Reddy, M.S.; El Enshasy, H. Plant Growth Promoting Rhizobacteria (PGPR) as Green Bioinoculants: Recent Developments, Constraints, and Prospects. Sustainability 2021, 13, 1140. [Google Scholar] [CrossRef]
- Mahmud, A.A.; Upadhyay, S.K.; Srivastava, A.K.; Bhojiya, A.A. Biofertilizers: A Nexus between soil fertility and crop productivity under abiotic stress. Curr. Res. Environ. Sustain. 2021, 3, 100063. [Google Scholar] [CrossRef]
- Sagar, A.; Yadav, S.S.; Sayyed, R.Z.; Sharma, S.; Ramteke, P.W. Bacillus subtilis: A Multifarious Plant Growth Promoter, Biocontrol Agent, and Bioalleviator of Abiotic Stress. In Bacilli in Agrobiotechnology; Springer: Cham, Switzerland, 2022; pp. 561–580. [Google Scholar] [CrossRef]
- Hurek, T.; Reinhold-Hurek, B. Azoarcus sp. strain BH72 as a model for nitrogen-fixing grass endophytes. J. Biotechnol. 2003, 106, 169–178. [Google Scholar] [CrossRef]
- Ahemad, M.; Kibret, M. Mechanisms and applications of plant growth promoting rhizobacteria: Current perspective. J. King Saud Univ. Sci. 2014, 26, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Pii, Y.; Mimmo, T.; Tomasi, N.; Terzano, R.; Cesco, S.; Crecchio, C. Microbial interactions in the rhizosphere: Beneficial influences of plant growth-promoting rhizobacteria on nutrient acquisition process. A review. Biol. Fertil. Soils 2015, 51, 403–415. [Google Scholar] [CrossRef]
- Ramakrishna, W.; Yadav, R.; Li, K. Plant growth promoting bacteria in agriculture: Two sides of a coin. Appl. Soil Ecol. 2019, 138, 10–18. [Google Scholar] [CrossRef]
- Kumar, S.; Diksha; Sindhu, S.S.; Kumar, R. Biofertilizers: An ecofriendly technology for nutrient recycling and environmental sustainability. Curr. Res. Microb. Sci. 2022, 3, 100094. [Google Scholar] [CrossRef] [PubMed]
- Schallmey, M.; Singh, A.; Ward, O.P. Developments in the use of Bacillus species for industrial production. Can. J. Microbiol. 2004, 50, 1–17. [Google Scholar] [CrossRef]
- Alabouvette, C.; Olivain, C.; Steinberg, C. Biological Control of Plant Diseases: The European Situation. Eur. J. Plant Pathol. 2006, 114, 329–341. [Google Scholar] [CrossRef]
- Borriss, R. Use of plant-associated Bacillus strains as biofertilizers and biocontrol agents in agriculture. In Bacteria in Agrobiology: Plant Growth Responses; Springer: Berlin/Heidelberg, Germany, 2011; pp. 41–76. [Google Scholar] [CrossRef]
- Hashem, A.; Tabassum, B.; Fathi Abd Allah, E. Bacillus subtilis: A plant-growth promoting rhizobacterium that also impacts biotic stress. Saudi J. Biol. Sci. 2019, 26, 1291–1297. [Google Scholar] [CrossRef]
- Goswami, D.; Thakker, J.N.; Dhandhukia, P.C. Portraying mechanics of plant growth promoting rhizobacteria (PGPR): A review. Cogent Food Agric. 2016, 2, 1127500. [Google Scholar] [CrossRef]
- Di Benedetto, N.A.; Corbo, M.R.; Campaniello, D.; Cataldi, M.P.; Bevilacqua, A.; Sinigaglia, M.; Flagella, Z. The role of Plant Growth Promoting Bacteria in improving nitrogen use efficiency for sustainable crop production: A focus on wheat. AIMS Microbiol. 2017, 3, 413–434. [Google Scholar] [CrossRef]
- Miljaković, D.; Marinković, J.; Balešević-Tubić, S. The Significance of Bacillus spp. in Disease Suppression and Growth Promotion of Field and Vegetable Crops. Microorganisms 2020, 8, 1037. [Google Scholar] [CrossRef]
- Fujita, Y.; Yasuda, Y.; Tochikubo, K. Permeability of Gentamicin and Polymyxin B into the Inside of Bacillus subtilis Spores. Microbiol. Immunol. 1990, 34, 1013–1023. [Google Scholar] [CrossRef] [PubMed]
- Nurmatov, S.; Akhmedova, Z.R.; Rakhimov, D.A. Isolation, Purification, and Properties of α-Amylase from Bacillus subtilis-7A. Chem. Nat. Compd. 2001, 37, 364–368. [Google Scholar] [CrossRef]
- Rey, M.W.; Ramaiya, P.; Nelson, B.A.; Brody-Karpin, S.D.; Zaretsky, E.J.; Tang, M.; De Leon, A.L.; Xiang, H.; Gusti, V.; Clausen, I.G.; et al. Complete genome sequence of the industrial bacterium Bacillus licheniformis and comparisons with closely related Bacillus species. Genome Biol. 2004, 5, r77. [Google Scholar] [CrossRef] [Green Version]
- Stein, T. Bacillus subtilis antibiotics: Structures, syntheses and specific functions. Mol. Microbiol. 2005, 56, 845–857. [Google Scholar] [CrossRef]
- Govindasamy, V.; Senthilkumar, M.; Magheshwaran, V.; Kumar, U.; Bose, P.; Sharma, V.; Annapurna, K. Bacillus and Paenibacillus spp.: Potential PGPR for sustainable agriculture. In Plant Growth and Health Promoting Bacteria; Springer: Berlin/Heidelberg, Germany, 2010; pp. 333–364. [Google Scholar]
- Vlamakis, H.; Chai, Y.; Beauregard, P.; Losick, R.; Kolter, R. Sticking together: Building a biofilm the Bacillus subtilis way. Nat. Rev. Microbiol. 2013, 11, 157–168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hobley, L.; Ostrowski, A.; Rao, F.V.; Bromley, K.M.; Porter, M.; Prescott, A.R.; MacPhee, C.E.; van Aalten, D.M.F.; Stanley-Wall, N.R. BslA is a self-assembling bacterial hydrophobin that coats the Bacillus subtilis biofilm. Proc. Natl. Acad. Sci. USA 2013, 110, 13600–13605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kobayashi, K.; Iwano, M. BslA(YuaB) forms a hydrophobic layer on the surface of Bacillus subtilis biofilms. Mol. Microbiol. 2012, 85, 51–66. [Google Scholar] [CrossRef]
- Hong, Z.-N.; Yan, J.; Lu, H.-L.; Jiang, J.; Li, J.-Y.; Xu, R.-K. Inhibition of phosphate sorptions on four soil colloids by two bacteria. Environ. Pollut. 2021, 290, 118001. [Google Scholar] [CrossRef]
- Bardin, M.; Ajouz, S.; Comby, M.; Lopez-Ferber, M.; Graillot, B.; Siegwart, M.; Nicot, P.C. Is the efficacy of biological control against plant diseases likely to be more durable than that of chemical pesticides? Front. Plant Sci. 2015, 6, 566. [Google Scholar] [CrossRef]
- Blake, C.; Christensen, M.N.; Kovács, Á.T. Molecular Aspects of Plant Growth Promotion and Protection by Bacillus subtilis. Mol. Plant-Microbe Interact. 2021, 34, 15–25. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharyya, P.N.; Jha, D.K. Plant growth-promoting rhizobacteria (PGPR): Emergence in agriculture. World J. Microbiol. Biotechnol. 2011, 28, 1327–1350. [Google Scholar] [CrossRef] [PubMed]
- Labuda, M.; Saeid, A.; Chojnacka, K.; Górecki, H. Use of Bacillus megaterium in solubilization of phosphorus. Przem. Chem. 2012, 91, 837–840. (In Polish) [Google Scholar]
- Saeid, A.; Prochownik, E.; Dobrowolska-Iwanek, J. Phosphorus Solubilization by Bacillus Species. Molecules 2018, 23, 2897. [Google Scholar] [CrossRef] [Green Version]
- Heyndrickx, M.; Vandemeulebroecke, K.; Hoste, B.; Janssen, P.; Kersters, K.; DE Vos, P.; Logan, N.A.; Ali, N.; Berkeley, R.C.W. Reclassification of Paenibacillus (formerly Bacillus) pulvifaciens (Nakamura 1984) Ash et al. 1994, a Later Subjective Synonym of Paenibacillus (formerly Bacillus) larvae (White 1906) Ash et al. 1994, as a Subspecies of P. larvae, with Emended Descriptions of P. larvae as P. larvae subsp. larvae and P. larvae subsp. pulvifaciens. Int. J. Syst. Evol. Microbiol. 1996, 46, 270–279. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Li, Q.; Li, Y.; Guan, G.; Chen, S. Paenibacillus strains with nitrogen fixation and multiple beneficial properties for promoting plant growth. PeerJ 2019, 7, e7445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hao, T.; Chen, S. Colonization of Wheat, Maize and Cucumber by Paenibacillus polymyxa WLY78. PLoS ONE 2017, 12, e0169980. [Google Scholar] [CrossRef] [PubMed]
- Hussain, A.; Ahmad, M.; Nafees, M.; Iqbal, Z.; Luqman, M.; Jamil, M.; Maqsood, A.; Mora-Poblete, F.; Ahmar, S.; Chen, J.-T.; et al. Plant-growth-promoting Bacillus and Paenibacillus species improve the nutritional status of Triticum aestivum L. PLoS ONE 2020, 15, e0241130. [Google Scholar] [CrossRef]
- Aquino, J.P.A.; Antunes, J.E.L.; Bonifácio, A.; Rocha, S.M.B.; Amorim, M.R.; Neto, F.A.; Araujo, A.S.F. Plant growth-promoting bacteria improve growth and nitrogen metabolism in maize and sorghum. Theor. Exp. Plant Physiol. 2021, 33, 249–260. [Google Scholar] [CrossRef]
- Kaziūnienė, J.; Mažylytė, R.; Krasauskas, A.; Toleikienė, M.; Gegeckas, A. Optimizing the Growth Conditions of the Selected Plant-Growth-Promoting Rhizobacteria Paenibacillus sp. MVY-024 for Industrial Scale Production. Biology 2022, 11, 745. [Google Scholar] [CrossRef]
- Dos Santos, R.M.; Diaz, P.A.E.; Lobo, L.L.B.; Rigobelo, E.C. Use of Plant Growth-Promoting Rhizobacteria in Maize and Sugarcane: Characteristics and Applications. Front. Sustain. Food Syst. 2020, 4, 136. [Google Scholar] [CrossRef]
- Seldin, L.; van Elsas, J.D.; Penido, E.G.C. Bacillus polymyxa bacteriophages from Brazilian soils. Antonie Leeuwenhoek 1984, 50, 39–51. [Google Scholar] [CrossRef] [PubMed]
- De Lima, B.D.; Penido, E.G.C. Inoculation in vitro of wheat seedlings and wheat straw cultures with Bacillus azotofixans. Plant Soil 1989, 113, 133–136. [Google Scholar] [CrossRef]
- Rosado, A.S.; Duarte, G.F.; Seldin, L.; Van Elsas, J.D. Genetic Diversity of nifH Gene Sequences in Paenibacillus azotofixans Strains and Soil Samples Analyzed by Denaturing Gradient Gel Electrophoresis of PCR-Amplified Gene Fragments. Appl. Environ. Microbiol. 1998, 64, 2770–2779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Padda, K.P.; Puri, A.; Chanway, C.P. Paenibacillus polymyxa: A prominent biofertilizer in the whole organism and biocontrol agent for sustainable agriculture. In Agriculturally Important Microbes for Sustainable Agriculture; Springer: Singapore, 2017; pp. 165–191. [Google Scholar]
- Seldin, L. Paenibacillus, nitrogen fixation and soil fertility. In Endospore-Forming Soil Bacteria; Springer: Berlin/Heidelberg, Germany, 2011; pp. 287–307. [Google Scholar]
- Stanisz, A. Accessible Course in Statistics Based on the STATISTICA PL Software on Examples from Medicine. In Basic Statistics; StatSoft Polska: Kraków, Poland, 2007; Volume 1. (In Polish) [Google Scholar]
- Rabiej, M. Statistics with the Program Statistica; Wydawnictwo Helion: Gliwice, Poland, 2012. (In Polish) [Google Scholar]
- Wątroba, J. Statystyka I Data Mining W Badaniach Naukowych; Wydawnictwo StatSoft Polska: Kraków, Poland, 2006. (In Polish) [Google Scholar]
- Starck, A. Integration of biomass partitioning includes total export from photosynthate donors and partitioning of assimilates between various sinks. Zesz. Probl. Post. Nauk Rol. 2002, 481, 113–123. [Google Scholar]
- Anjum, S.A.; Xie, X.Y.; Wang, L.C.; Saleem, M.F.; Man, C.; Lei, W. Morphological, physiological and biochemical responses of plants to drought stress. Afr. J. Agric. Res. 2011, 6, 2026–2032. [Google Scholar] [CrossRef]
- Sehgal, A.; Sita, K.; Siddique, K.H.M.; Kumar, R.; Bhogireddy, S.; Varshney, R.K.; HanumanthaRao, B.; Nair, R.M.; Prasad, P.V.V.; Nayyar, H. Drought or/and Heat-Stress Effects on Seed Filling in Food Crops: Impacts on Functional Biochemistry, Seed Yields, and Nutritional Quality. Front. Plant Sci. 2018, 9, 1705. [Google Scholar] [CrossRef] [Green Version]
- Hlaváčová, M.; Klem, K.; Rapantová, B.; Novotná, K.; Urban, O.; Hlavinka, P.; Smutná, P.; Horáková, V.; Škarpa, P.; Pohanková, E.; et al. Interactive effects of high temperature and drought stress during stem elongation, anthesis and early grain filling on the yield formation and photosynthesis of winter wheat. Field Crop. Res. 2018, 221, 182–195. [Google Scholar] [CrossRef]
- Sharma, A.; Kumar, V.; Shahzad, B.; Ramakrishnan, M.; Sidhu, G.P.S.; Bali, A.S.; Handa, N.; Kapoor, D.; Yadav, P.; Khanna, K.; et al. Photosynthetic Response of Plants Under Different Abiotic Stresses: A Review. J. Plant Growth Regul. 2020, 39, 509–531. [Google Scholar] [CrossRef]
- Jeppesen, E.; Kronvang, B.; Olesen, J.E.; Audet, J.; Søndergaard, M.; Hoffmann, C.C.; Andersen, H.E.; Lauridsen, T.L.; Liboriussen, L.; Larsen, S.E. Climate Change effect on nitrogen loading from cultivated catchments in Europe: Implications for nitrogen retention, ecological stat of lakes and adaptation. Hydrobiologia 2011, 663, 1–21. [Google Scholar] [CrossRef]
- Skowera, B.; Kopcinska, J.; Kolodziejczyk, M.; Kopec, B. Precipitation deficiencies and excesses during the growing season of winter rape and winter wheat in Poland (1971–2010). Acta Agrophys. 2015, 22, 193–207. [Google Scholar]
- Turan, M.; Gulluce, M.; von Wirén, N.; Sahin, F. Yield promotion and phosphorus solubilization by plant growth–promoting rhizobacteria in extensive wheat production in Turkey. J. Plant Nutr. Soil Sci. 2012, 175, 818–826. [Google Scholar] [CrossRef]
- Schütz, L.; Gattinger, A.; Meier, M.; Müller, A.; Boller, T.; Mäder, P.; Mathimaran, N. Improving Crop Yield and Nutrient Use Efficiency via Biofertilization—A Global Meta-analysis. Front. Plant Sci. 2018, 8, 2204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mayer, J.; Scheid, S.; Widmer, F.; Fliessbach, A.; Oberholzer, H.-R. How effective are ‘Effective microorganisms® (EM)’? Results from a field study in temperate climate. Appl. Soil Ecol. 2010, 46, 230–239. [Google Scholar] [CrossRef]
- Santoyo, G.; Moreno-Hagelsieb, G.; del Carmen Orozco-Mosqueda, M.; Glick, B.R. Plant growth-promoting bacterial endophytes. Microbiol. Res. 2016, 183, 92–99. [Google Scholar] [CrossRef]
- Shahzad, R.; Khan, A.L.; Bilal, S.; Waqas, M.; Kang, S.-M.; Lee, I.-J. Inoculation of abscisic acid-producing endophytic bacteria enhances salinity stress tolerance in Oryza sativa. Environ. Exp. Bot. 2017, 136, 68–77. [Google Scholar] [CrossRef]
- Lastochkina, O.; Pusenkova, L.; Yuldashev, R.; Babaev, M.; Garipova, S.; Blagova, D.; Khairullin, R.; Aliniaeifard, S. Effects of Bacillus subtilis on some physiological and biochemical parameters of Triticum aestivum L. (wheat) under salinity. Plant Physiol. Biochem. 2017, 121, 80–88. [Google Scholar] [CrossRef]
- de Lima, B.C.; Moro, A.L.; Santos, A.C.P.; Bonifacio, A.; Araujo, A.S.F.; de Araujo, F.F. Bacillus subtilis ameliorates water stress tolerance in maize and common bean. J. Plant Interact. 2019, 14, 432–439. [Google Scholar] [CrossRef] [Green Version]
- Ali, S.; Khan, N. Delineation of mechanistic approaches employed by plant growth promoting microorganisms for improving drought stress tolerance in plants. Microbiol. Res. 2021, 249, 126771. [Google Scholar] [CrossRef]
- Maslennikova, D.; Lastochkina, O. Contribution of Ascorbate and Glutathione in Endobacteria Bacillus subtilis-Mediated Drought Tolerance in Two Triticum aestivum L. Genotypes Contrasting in Drought Sensitivity. Plants 2021, 10, 2557. [Google Scholar] [CrossRef]
- Khan, N.; Mehmood, A.; Ali, S.; Shahid, M.A. Plant growth-promoting rhizobacteria and their role as bio-inoculants for sustainable agriculture under stressful environments. In Biofertilizers; Elsevier: Amsterdam, The Netherlands, 2021; pp. 313–321. [Google Scholar] [CrossRef]
- Masclaux-Daubresse, C.; Daniel-Vedele, F.; Dechorgnat, J.; Chardon, F.; Gaufichon, L.; Suzuki, A. Nitrogen uptake, assimilation and remobilization in plants: Challenges for sustainable and productive agriculture. Ann. Bot. 2010, 105, 1141–1157. [Google Scholar] [CrossRef]
- Kuan, K.B.; Othman, R.; Rahim, K.A.; Shamsuddin, Z.H. Plant Growth-Promoting Rhizobacteria Inoculation to Enhance Vegetative Growth, Nitrogen Fixation and Nitrogen Remobilisation of Maize under Greenhouse Conditions. PLoS ONE 2016, 11, e0152478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, K.; Dubey, N.K.; Singh, S.P.; Kheni, J.K.; Gupta, S.; Varshney, A. Plant Growth-Promoting Rhizobacteria (PGPR): Current and Future Prospects for Crop Improvement. In Current Trends in Microbial Biotechnology for Sustainable Agriculture; Springer: Singapore, 2021; pp. 203–226. [Google Scholar] [CrossRef]
- Pan, J.; Zhu, Y.; Cao, W.; Dai, T.; Jiang, D. Predicting the Protein Content of Grain in Winter Wheat with Meteorological and Genotypic Factors. Plant Prod. Sci. 2006, 9, 323–333. [Google Scholar] [CrossRef]
- Rollins, J.A.; Habte, E.; Templer, S.E.; Colby, T.; Schmidt, J.; von Korff, M. Leaf proteome alterations in the context of physiological and morphological responses to drought and heat stress in barley (Hordeum vulgare L.). J. Exp. Bot. 2013, 64, 3201–3212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.; Zhang, J.; Wang, Z.; Zhu, Q.; Liu, L. Abscisic acid and cytokinins in the root exudates and leaves and their relationship to senescence and remobilization of carbon reserves in rice subjected to water stress during grain filling. Planta 2002, 215, 645–652. [Google Scholar] [CrossRef] [PubMed]
- Liu, E.; Mei, X.; Yan, C.; Gong, D.; Zhang, Y. Effects of water stress on photosynthetic characteristics, dry matter translocation and WUE in two winter wheat genotypes. Agric. Water Manag. 2016, 167, 75–85. [Google Scholar] [CrossRef]
- Buczek, J.; Jańczak-Pieniążek, M. Hybrid wheat response to high nitrogen application rates and foliar fertilization. Biul. Inst. Hod. I Aklim. Roślin 2021, 296, 17–24. [Google Scholar] [CrossRef]
- Islam, M.R.; Haque, K.S.; Akter, N.; Karim, M.A. Leaf chlorophyll dynamics in wheat based on SPAD meter reading and its relationship with grain yield. Sci. Agric. 2014, 4, 13–18. [Google Scholar] [CrossRef]
- Monostori, I.; Árendás, T.; Hoffman, B.; Galiba, G.; Gierczik, K.; Szira, F.; Vágújfalvi, A. Relationship between SPAD value and grain yield can be affected by cultivar, environment and soil nitrogen content in wheat. Euphytica 2016, 211, 103–112. [Google Scholar] [CrossRef] [Green Version]
- Wojcieska, U. Fizjologiczna rola azotu w ksztaltowaniu plonu roslin. Czesc II. Zywienie roslin azotem a fotosynteza, fotorespiracja i oddychanie ciemniowe. Postępy Nauk Rol. 1994, 41, 127–143. (In Polish) [Google Scholar]
- Dong, Z.; Zhang, X.; Li, J.; Zhang, C.; Wei, T.; Yang, Z.; Cai, T.; Zhang, P.; Ding, R.; Jia, Z. Photosynthetic characteristics and grain yield of winter wheat (Triticum aestivum L.) in response to fertilizer, precipitation, and soil water storage before sowing under the ridge and furrow system: A path analysis. Agric. For. Meteorol. 2019, 272–273, 12–19. [Google Scholar] [CrossRef]
- Olszewski, J.; Makowska, M.; Pszczółkowska, A.; Okorski, A.; Bieniaszewski, T. The effect of nitrogen fertilization on flag leaf and ear photosynthesis and grain yield of spring wheat. Plant Soil Environ. 2014, 60, 531–536. [Google Scholar] [CrossRef] [Green Version]
- Carmo-Silva, E.; Andralojc, P.J.; Scales, J.C.; Driever, S.; Mead, A.; Lawson, T.; A Raines, C.; Parry, M. Phenotyping of field-grown wheat in the UK highlights contribution of light response of photosynthesis and flag leaf longevity to grain yield. J. Exp. Bot. 2017, 68, 3473–3486. [Google Scholar] [CrossRef] [PubMed]
- Murchie, E.H.; Kefauver, S.; Araus, J.L.; Muller, O.; Rascher, U.; Flood, P.J.; Lawson, T. Measuring the dynamic photosynthome. Ann. Bot. 2018, 122, 207–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanchez-Bragado, R.; Vicente, R.; Molero, G.; Serret, M.D.; Maydup, M.L.; Araus, J.L. New avenues for increasing yield and stability in C3 cereals: Exploring ear photosynthesis. Curr. Opin. Plant Biol. 2020, 56, 223–234. [Google Scholar] [CrossRef] [PubMed]
- Ashraf, M. Effect of salicylic acid applied through rooting medium on drought tolerance of wheat. Pak. J. Bot. 2006, 38, 1127–1136. [Google Scholar]
- Kołodziejczyk, M. Effect of nitrogen fertilization and application of soil properties improving microbial preparations on the content of mineral nitrogen in soil after spring wheat harvesting. J. Cent. Eur. Agric. 2013, 14, 306–318. [Google Scholar] [CrossRef]
- Kocoń, A.; Jadczyszyn, T. The effect of microbiological preparations, method of their application and doses of nitrogen fertilization on the content of available phosphorus in the soil, and other selected chemical indicators of soil fertility. Pol. J. Agron. 2015, 21, 11–18. (In Polish) [Google Scholar]
- Saxena, A.; Kumar, M.; Chakdar, H.; Anuroopa, N.; Bagyaraj, D.; Murugan, K. Bacillus species in soil as a natural resource for plant health and nutrition. J. Appl. Microbiol. 2020, 128, 1583–1594. [Google Scholar] [CrossRef] [Green Version]
- Tahir, M.; Khalid, U.; Ijaz, M.; Shah, G.M.; Naeem, M.A.; Shahid, M.; Mahmood, K.; Ahmad, N.; Kareem, F. Combined application of bio-organic phosphate and phosphorus solubilizing bacteria (Bacillus strain MWT 14) improve the performance of bread wheat with low fertilizer input under an arid climate. Braz. J. Microbiol. 2018, 49, 15–24. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Growing Season | Mean Temperature (°C) | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| IX * | X | XI-III | IV | V | VI | VII | VIII | Av. | |
| 2016/2017 | 13.6 | 6.1 | 2.4 | 5.7 | 12.1 | 15.7 | 16.8 | 17.4 | 7.5 |
| 2017/2018 | 12.8 | 8.7 | 3.9 | 10.8 | 15.7 | 17.2 | 19.7 | 19.2 | 8.6 |
| 2018/2019 | 14.5 | 8.7 | 3.3 | 8.0 | 11.6 | 20.2 | 17.1 | 18.5 | 8.8 |
| 1981–2010 | 12.8 | 8.0 | 2.9 | 7.7 | 13.5 | 16.1 | 18.7 | 17.9 | 7.9 |
| Rainfall (mm) | Sum | ||||||||
| 2016/2017 | 21.1 | 104.3 | 84.8 | 59.1 | 25.1 | 74.5 | 107.6 | 63.1 | 693.8 |
| 2017/2018 | 168.1 | 114.9 | 42.4 | 33.5 | 25.0 | 53.7 | 141.0 | 44.6 | 713.4 |
| 2018/2019 | 20.3 | 84.7 | 16.0 | 0.0 | 142.8 | 120.6 | 56.3 | 55.9 | 677.6 |
| 1981–2010 | 56.9 | 42.6 | 44.8 | 33.3 | 58.5 | 80.4 | 74.2 | 59.4 | 581.8 |
| Treatment (Shortcut) | Component | Application Date/Dose |
|---|---|---|
| NPK (NPK) | N (ammonium sulphate 34%; N (ammonium sulphate 34%; P (triple superphosphate 20%); K (potash salt, 49.8%) | BBCH23–24/90.0 kg ha−1 BBCH31–32/60.0 kg ha−1 Pre-sowing/70.0 kg ha−1 Pre-sowing/100.0 kg ha−1 |
| NPK * + Paenibacillus azotofixans (NPK + PA) | Paenibacillus azotofixans 1 × 109 CFU ** in 1 g of the product (maltodextrin) | BBCH23–24/1.0 L ha−1 BBCH31–32/1.0 L ha−1 |
| NPK * + Bacillus megaterium (NPK + BM) | Bacillus megaterium 1 × 109 CFU in 1 g of the product (maltodextrin) | BBCH23–24/1.0 L ha−1 BBCH31–32/1.0 L ha−1 |
| NPK * + Bacillus subtilis (NPK + BS) | Bacillus subtilis 5 × 109 CFU in 1 g of the product (maltodextrin) | BBCH23–24/1.0 L ha−1 BBCH31–32/1.0 L ha−1 |
| NPK * + Paenibacillus azotofixans + Bacillus megaterium + Bacillus subtilis (NPK + PA + BM + BS) | Paenibacillus azotofixans Bacillus megaterium Bacillus subtilis | BBCH23–24/1.0 L ha−1 BBCH31–32/1.0 L ha−1 BBCH23–24/1.0 L ha−1 BBCH31–32/1.0 L ha−1 BBCH23–24/1.0 L ha−1 BBCH31–32/1.0 L ha−1 |
| Year | NPK | NPK + PA * | NPK + BM ** | NPK + BS *** | NPK + PA + BM + BS |
|---|---|---|---|---|---|
| 2017 | 8.72 ± 0.56 ab | 9.81 ± 0.23 a | 8.38 ± 0.69 abc | 8.60 ± 0.58 abc | 9.76 ± 0.89 a |
| 2018 | 6.46 ± 0.32 e | 6.41 ± 0.84 e | 5.81 ± 0.29 e | 6.45 ± 0.17 e | 7.13 ± 0.07 cd |
| 2019 | 6.51 ± 0.38 de | 9.10 ± 0.60 ab | 7.98 ± 0.45 bcd | 8.57 ± 0.91 abc | 8.71 ± 0.75 ab |
| Year | NPK | NPK + PA * | NPK + BM ** | NPK + BS *** | NPK + PA + BM + BS |
|---|---|---|---|---|---|
| 2017 | 103 ± 2.36 b | 108 ± 4.43 b | 108 ± 2.44 b | 109 ± 5.57 b | 109 ± 3.30 b |
| 2018 | 121 ± 4.78 a | 121 ± 2.21 a | 121 ± 3.30 a | 121 ± 6.18 a | 124 ± 6.39 a |
| 2019 | 109 ± 2.50 b | 108 ± 3.56 b | 106 ± 3.77 b | 108 ± 1.50 b | 110 ± 2.89 b |
| Year | NPK | NPK + PA * | NPK + BM ** | NPK + BS *** | NPK + PA + BM + BS |
|---|---|---|---|---|---|
| BBCH31 | |||||
| 2017 | 8.18 ± 3.04 e | 13.14 ± 0.46 cde | 9.33 ± 5.35 e | 15.58 ± 1.14 bcd | 20.02 ± 3.03 ab |
| 2018 | 17.23 ± 1.00 bc | 20.99 ± 0.29 ab | 10.57 ± 4.03 de | 17.17 ± 0.38 bc | 20.25 ± 2.11 ab |
| 2019 | 17.75 ± 0.07 bc | 23.77 ± 0.06 a | 18.58 ± 0.33 abc | 19.76 ± 0.16 ab | 23.77 ± 0.06 a |
| BBCH51 | |||||
| 2017 | 10.22 ± 0.91 de | 27.07 ± 5.15 a | 16.85 ± 4.10 bcd | 13.75 ± 0.65 cde | 23.74 ± 3.81 ab |
| 2018 | 19.53 ± 7.13 abc | 25.80 ± 0.52 a | 14.93 ± 6.07 cd | 19.12 ± 1.92 a–d | 27.08 ± 0.21 a |
| 2019 | 19.12 ± 0.71 a–d | 21.55a ± 3.75 bc | 5.89 ± 0.46 e | 19.33 ± 0.12 abc | 20.67 ± 0.06 abc |
| Treatments | BBCH31 | BBCH51 | ||
|---|---|---|---|---|
| Regression Equation | Coefficient of Determination R2 | Regression Equation | Coefficient of Determination R2 | |
| NPK | A = 0.0921·R + 2.5164·T − 24.5848 | 0.883 | A = −0.1753·R + 3.7715·T − 35.9334 | 0.585 |
| NPK + PA * | A = 0.0996·R + 2.1819·T − 15.7609 | 0.996 | A = −0.0160·R − 1.0631·T + 44.9450 | 0.354 |
| NPK + BM ** | A = 0.0801·R + 0.3463·T + 3.1292 | 0.599 | A = −0.0479·R − 1.9456·T + 50.9656 | 0.629 |
| NPK + BS *** | A = 0.0374·R + 0.4448·T + 9.2531 | 0.889 | A = −0.0972·R + 2.2360·T − 14.1208 | 0.867 |
| NPK + PA + BM + BS | A = 0.0321·R + 0.0641·T + 18.4380 | 0.464 | A = −0.1206·R + 0.5517·T + 24.0601 | 0.653 |
| Year | NPK | NPK + PA * | NPK + BM ** | NPK + BS *** | NPK + PA + BM + BS |
|---|---|---|---|---|---|
| N-NO3 | |||||
| 2017 | 2.32 ± 0.52 bc | 3.79 ±0.45 b | 2.77 ± 0.43 bc | 3.30 ± 0.57 bc | 6.05 ± 0.73 a |
| 2018 | 1.82 ± 0.48 c | 3.25 ± 0.81 bc | 2.82 ± 0.45 bc | 2.60 ± 1.17 bc | 2.80 ± 0.45 bc |
| 2019 | 2.25 ± 0.18 bc | 2.75 ± 0.51 bc | 2.22 ± 0.42 c | 1.75 ± 0.33 c | 3.05 ± 0.96 bc |
| N-NH4 | |||||
| 2017 | 1.12 ± 0.11 d | 2.97 ± 0.81 a | 2.04 ± 0.14 a–d | 3.03 ± 0.52 a | 2.13 ± 0.78 a–d |
| 2018 | 1.48b ± 0.50 cd | 1.46 ± 0.07 bcd | 2.15 ± 0.01 abc | 1.88 ± 0.02 bcd | 2.99 ± 0.58 a |
| 2019 | 1.15 ± 0.07 cd | 1.29 ± 0.21 bcd | 1.56 ± 0.08 bcd | 1.38 ± 0.33 bcd | 2.27 ± 0.32 ab |
| Year | NPK | NPK + PA * | NPK + BM ** | NPK + BS *** | NPK + PA + BM + BS |
|---|---|---|---|---|---|
| 2017 | 7.78 ± 1.34 b–e | 6.74 ± 0.84 de | 8.73 ± 1.24 bcd | 7.99 ± 1.52 b–e | 11.84 ± 0.24 a |
| 2018 | 5.91 ± 0.18 e | 6.14 ± 0.35 e | 8.54 ± 1.62 bcd | 7.26 ± 0.18 b–e | 9.24 ± 1.28 b |
| 2019 | 6.89 ± 0.09 cde | 7.39 ± 0.46 b–e | 8.56 ± 0.65 bcd | 9.03 ± 0.38 bc | 11.77 ± 0.48 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stępień, A.; Wojtkowiak, K.; Kolankowska, E. Effect of Commercial Microbial Preparations Containing Paenibacillus azotofixans, Bacillus megaterium and Bacillus subtilis on the Yield and Photosynthesis of Winter Wheat and the Nitrogen and Phosphorus Content in the Soil. Appl. Sci. 2022, 12, 12541. https://doi.org/10.3390/app122412541
Stępień A, Wojtkowiak K, Kolankowska E. Effect of Commercial Microbial Preparations Containing Paenibacillus azotofixans, Bacillus megaterium and Bacillus subtilis on the Yield and Photosynthesis of Winter Wheat and the Nitrogen and Phosphorus Content in the Soil. Applied Sciences. 2022; 12(24):12541. https://doi.org/10.3390/app122412541
Chicago/Turabian StyleStępień, Arkadiusz, Katarzyna Wojtkowiak, and Ewelina Kolankowska. 2022. "Effect of Commercial Microbial Preparations Containing Paenibacillus azotofixans, Bacillus megaterium and Bacillus subtilis on the Yield and Photosynthesis of Winter Wheat and the Nitrogen and Phosphorus Content in the Soil" Applied Sciences 12, no. 24: 12541. https://doi.org/10.3390/app122412541