
Digital Droplet-PCR for Quantification of Viable Campylobacter jejuni and Campylobacter coli in Chicken Meat Rinses

Abstract
:Featured Application
Abstract
1. Introduction
2. Materials and Methods
2.1. Strains and Growth Conditions
2.2. Live and Dead Cell Standards
2.3. Raw Meat Samples and Rinses
- (A)
- Campylobacter-free rinses were spiked with live and dead cell standards, and they were used for method comparison study (accuracy and relative trueness), as well as technical measurement uncertainty study. The absence of live and dead Campylobacter cells was confirmed previously in-house based on the three methods described in Section 2.7, Section 2.8 and Section 2.10. Each of the spiked meat rinses was divided into three aliquots, 1 mL for ddPCR with PMA, 1 mL for ddPCR without PMA, and 1 mL for the microbial reference method. The v-ddPCR results were compared to the results of the reference method [7];
- (B)
- Rinses prepared with naturally contaminated routine samples from LGL (2020) and retail samples (2019 to 2021) were used to investigate the applicability of v-ddPCR for quantifying Campylobacter in chicken rinses. The LGL routine samples consist of 19 chicken neck skin (NS) from Bavarian slaughterhouses and 13 chicken breast meat (BM, within the scope of zoonosis monitoring) from Bavarian retail. Additionally, 25 chicken meat retail samples (RS) like chicken neck skin, chicken breast meat, chicken thighs, chicken wings, and chicken drumstick were bought in supermarkets between October 2019 and March 2021 in the region of Munich. Five 1 mL aliquots were needed for each rinse: 1 mL was used for the quantification of Campylobacter with the microbiological reference assay, two times 1 mL for qPCR (with and without PMA treatment), and two times 1 mL for ddPCR (with and without PMA treatment). The v-ddPCR results were compared to the results of the reference method [7] and the v-qPCR [18].
2.4. DNA Extraction
2.5. DNA Quantification
2.6. Oligonucleotides for ddPCR and qPCR
2.7. Optimization of ddPCR
2.8. qPCR
2.9. Live/Dead Differentiation of Campylobacter in Meat Rinse Samples
2.10. Microbiological Reference Method
2.11. Determination of LOD95%
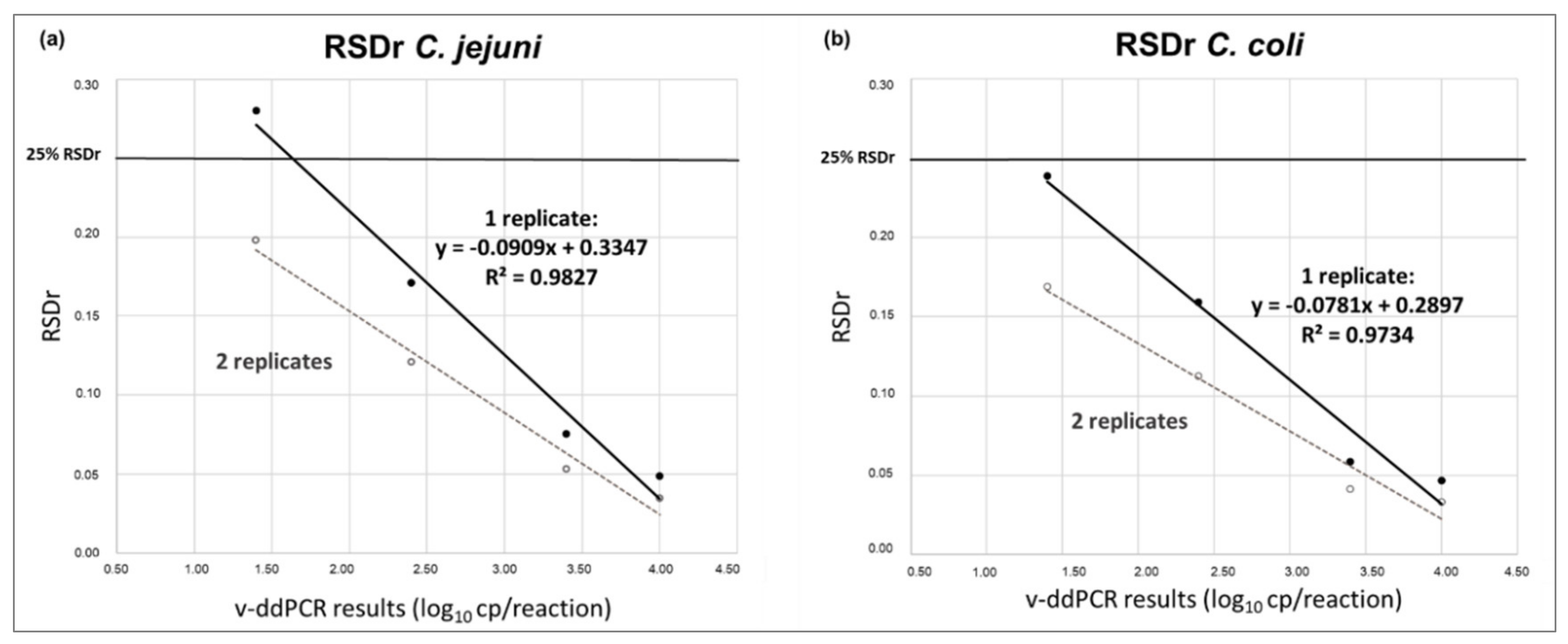
2.12. Precision—Relative Repeatability Standard Deviation (RSDr)
2.13. Determination of LOQ
2.14. Selectivity
2.15. Method Comparison Study According to ISO 16140-2:2016
2.15.1. Accuracy
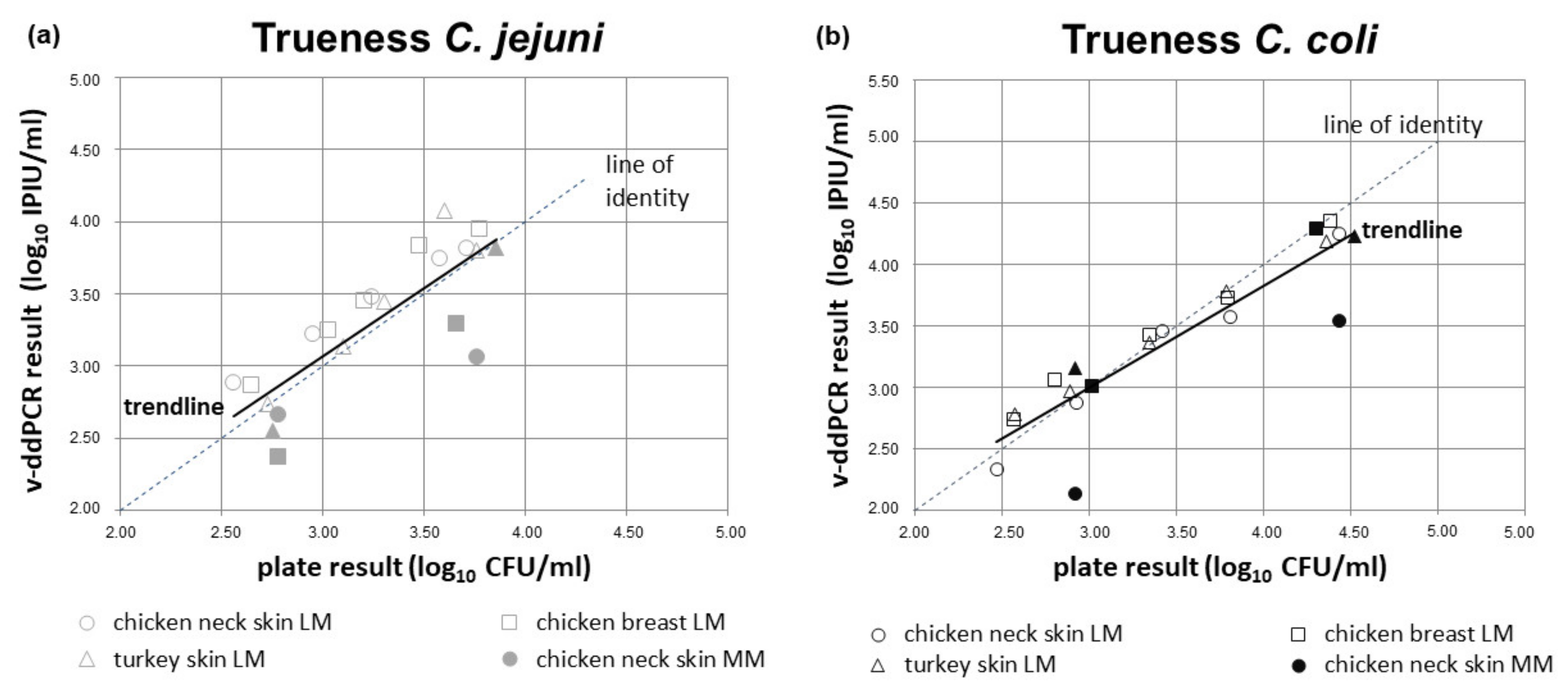
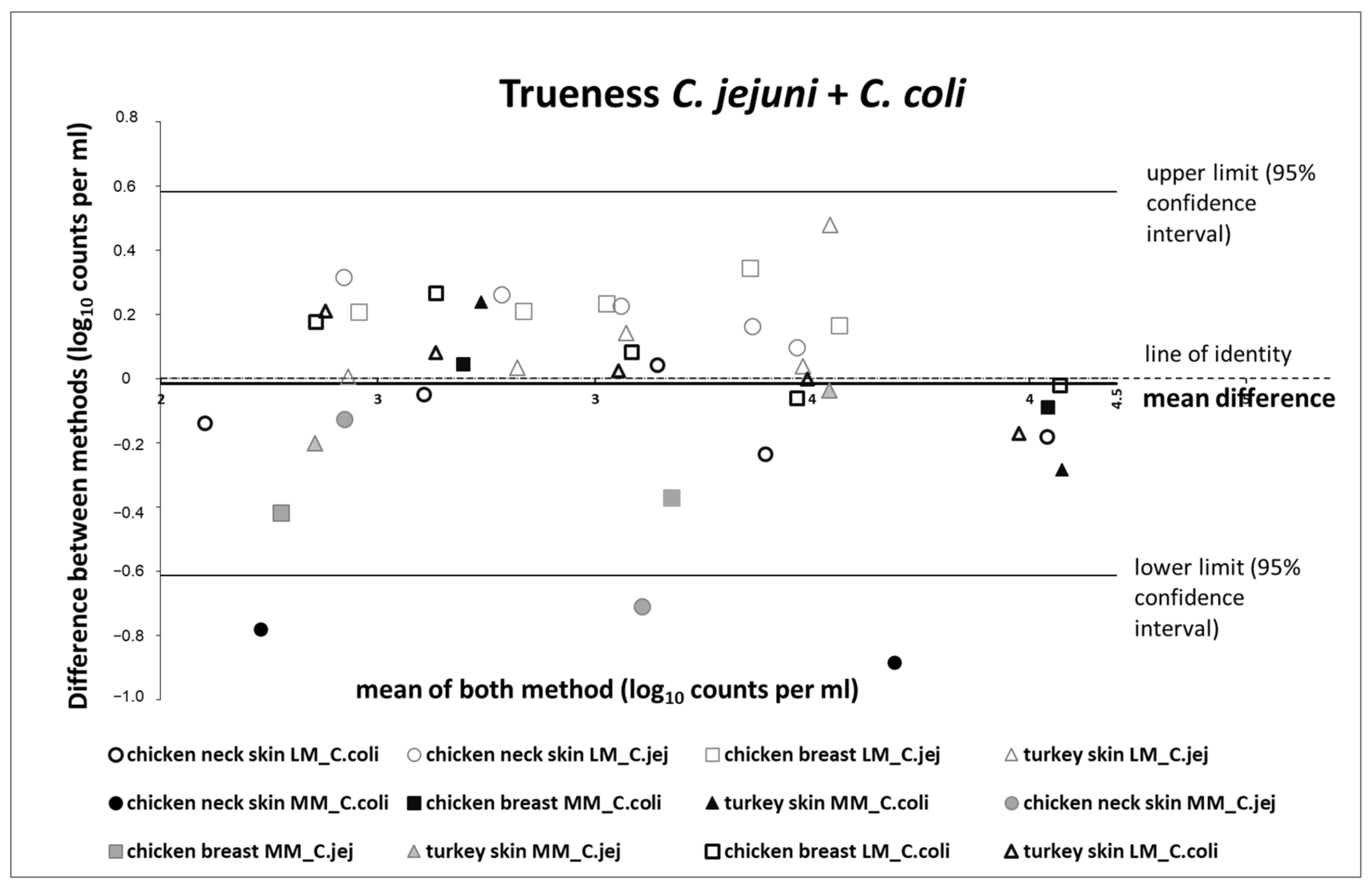
2.15.2. Relative Trueness
2.16. Technical Measurement Uncertainty
3. Results
3.1. Determination of LOD95%
3.2. Precision and RSDr
3.3. Determination of LOQ
3.4. Inclusivity and Exclusivity Study
3.5. Method Comparison Study According to ISO 16140-2:2016
3.5.1. Accuracy Profile Study
3.5.2. Relative Trueness Study
3.6. Technical Measurement Uncertainty
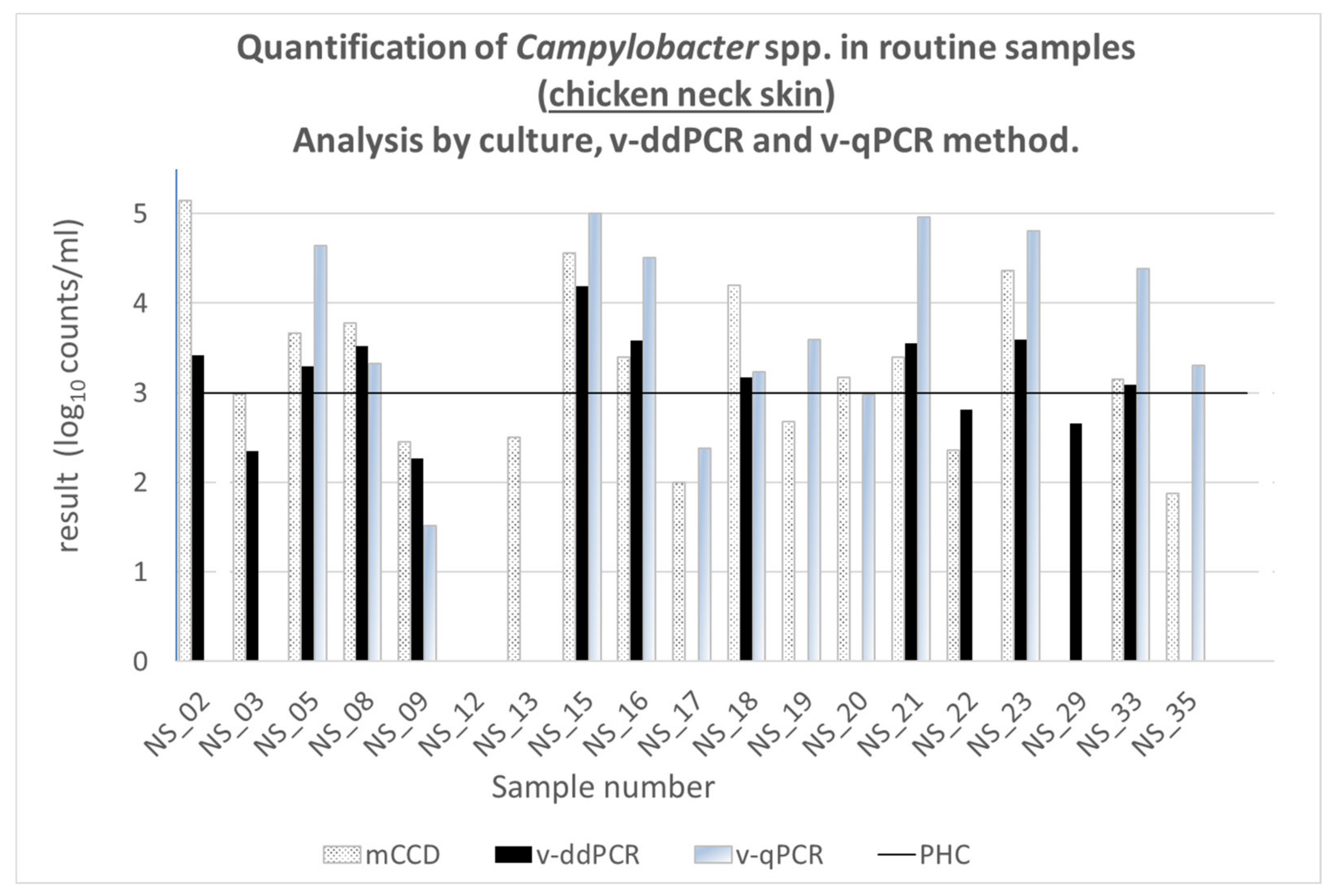
3.7. Performance of v-ddPCR on Routine and Retail Samples
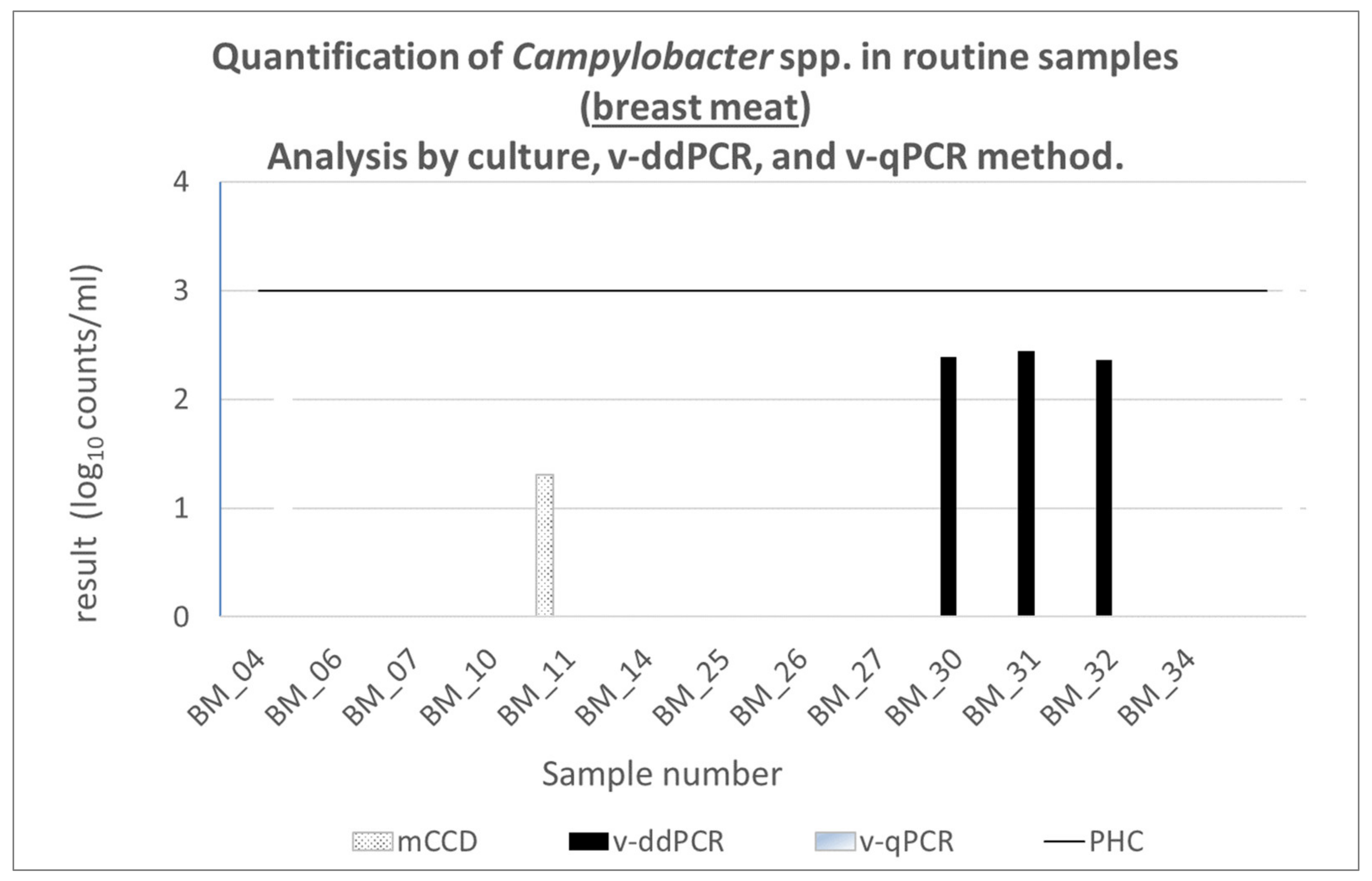
3.7.1. Routine Samples
3.7.2. Retail Samples
4. Discussion
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- EFSA BIOHAZ Panel (EFSA Panel on Biological Hazards). Scientific Opinion on quantification of the risk posed by broiler meat to human Campylobacteriosis in the EU. EFSA J. 2010, 8, 1437. [Google Scholar] [CrossRef]
- EFSA and ECDC (European Food Safety Authority and European Centre for Disease Prevention and Control). The European Union One Health 2018 Zoonoses Report. EFSA J. 2019, 17, 5926. [Google Scholar] [CrossRef] [Green Version]
- EFSA and ECDC (European Food Safety Authority and European Centre for Disease Prevention and Control). The European Union One Health 2019 Zoonoses Report. EFSA J. 2021, 19, 6406. [Google Scholar] [CrossRef]
- EFSA and ECDC (European Food Safety Authority and European Centre for Disease Prevention and Control). The European Union summary report on trends and sources of zoonoses, zoonotic agents and food-borne outbreaks in 2017. EFSA J. 2018, 16, e05500. [Google Scholar] [CrossRef]
- Nastasijevic, I.; Proscia, F.; Boskovic, M.; Glisic, M.; Blagojevic, B.; Sorgentone, S.; Kirbis, A.; Ferri, M. The European Union control strategy for Campylobacter spp. in the broiler meat chain. J. Food Saf. 2020, 40, e12819. [Google Scholar] [CrossRef]
- Commission Regulation (EU) 2017/1495 of 23 August 2017, Amending Regulation (EC) No 2073/2005 as Regards Campylobacter in Broiler Carcases. Available online: http://eurlex.europa.eu/eli/reg/2017/1495/oj (accessed on 18 April 2022).
- ISO 10272-2:2017; Microbiology of the Food Chain—Horizontal Method for Detection and Enumeration of Campylobacter spp.—Part 2: Colony-Count Technique. International Organization for Standardization: Geneva, Switzerland, 2017.
- Kim, J.C.; Oh, E.; Kim, J.; Jeon, B. Regulation of oxidative stress resistance in Campylobacter jejuni; a microaerophilic foodborne pathogen. Front. Microbiol. 2015, 6, 751. [Google Scholar] [CrossRef] [Green Version]
- Lv, R.; Wang, K.; Feng, J.; Heeney, D.D.; Liu, D.; Lu, X. Detection and Quantification of Viable but Non-culturable Campylobacter jejuni. Front. Microbiol. 2020, 10, 2920. [Google Scholar] [CrossRef]
- Goode, O.; Smith, A.; Zarkan, A.; Cama, J.; Invergo, B.M.; Belgami, D.; Caño-Muñiz, S.; Metz, J.; O’Neill, P.; Jeffries, A.; et al. Persister Escherichia coli Cells Have a Lower Intracellular pH than Susceptible Cells but Maintain Their pH in Response to Antibiotic Treatment. mBio 2021, 12, e0090921. [Google Scholar] [CrossRef]
- Bamford, R.; Smith, A.; Metz, J.; Glover, G.; Titball, R.W.; Pagliara, R. Investigating the physiology of viable but non-culturable bacteria by microfluidics and time-lapse microscopy. BMC Biol. 2017, 15, 121. [Google Scholar] [CrossRef]
- Josefsen, M.H.; Löfström, C.; Hansen, T.B.; Christensen, L.S.; Olsen, J.E.; Hoorfar, J. Rapid quantification of viable Campylobacter bacteria on chicken carcasses, using real-time PCR and propidium monoazide treatment, as a tool for quantitative risk assessment. Appl. Environ. Microbiol. 2010, 76, 5097–5104. [Google Scholar] [CrossRef] [Green Version]
- Wagley, S.; Morcrette, H.; Kovacs-Simon, A.; Yang, Z.R.; Power, A.; Tennant, R.K.; Love, J.; Murray, N.; Titball, R.W.; Butler, C.S. Bacterial dormancy: A subpopulation of viable but non-culturable cells demonstrates better fitness for revival. PLoS Pathog. 2021, 17, e1009194. [Google Scholar] [CrossRef] [PubMed]
- Baffone, W.; Casaroli, A.; Citterio, B.; Pierfelici, L.; Campana, R.; Vittoria, E.; Guaglianone, E.; Donelli, G. Campylobacter jejuni loss of culturability in aqueous microcosms and ability to resuscitate in a mouse model. Int. J. Food Microbiol. 2006, 107, 83–91. [Google Scholar] [CrossRef] [PubMed]
- Wideman, N.E.; Oliver, J.D.; Crandall, P.G.; Jarvis, N.A. Detection and Potential Virulence of Viable but Non-Culturable (VBNC) Listeria monocytogenes: A Review. Microorganisms 2021, 9, 194. [Google Scholar] [CrossRef] [PubMed]
- Gin, K.Y.; Goh, S.G. Modeling the effect of light and salinity on viable but non-culturable (VBNC) Enterococcus. Water Res. 2013, 47, 3315–3328. [Google Scholar] [CrossRef]
- Castro, A.G.S.A.; Dorneles, E.M.S.; Santos, E.L.S.; Alves, T.M.; Silva, G.R.; Figueiredo, T.C.; Assis, D.C.S.; Lage, A.P.; Cançado, S.V. Viability of Campylobacter spp. in frozen and chilled broiler carcasses according to real-time PCR with propidium monoazide pretreatment. Poult. Sci. 2018, 97, 1706–1711. [Google Scholar] [CrossRef] [PubMed]
- Stingl, K.; Heise, J.; Thieck, M.; Wulsten, I.F.; Pacholewicz, E.; Iwobi, A.N.; Govindaswamy, J.; Zeller-Péronnet, V.; Scheuring, S.; Luu, H.Q.; et al. Challenging the “gold standard” of colony-forming units—Validation of a multiplex real-time PCR for quantification of viable Campylobacter spp. in meat rinses. Int. J. Food Microbiol. 2021, 359, 109417. [Google Scholar] [CrossRef]
- Pacholewicz, E.; Buhler, C.; Wulsten, I.F.; Kraushaar, B.; Luu, H.Q.; Iwobi, A.N.; Huber, I.; Stingl, K. Internal sample process control improves cultivation-independent quantification of thermotolerant Campylobacter. Food Microbiol. 2019, 78, 53–61. [Google Scholar] [CrossRef]
- Pacholewicz, E.; Swart, A.; Lipman, L.J.; Wagenaar, J.A.; Havelaar, A.H.; Duim, B. Propidium monoazide does not fully inhibit the detection of dead Campylobacter on broiler chicken carcasses by qPCR. J. Microbiol. Methods 2013, 95, 32–38. [Google Scholar] [CrossRef]
- Lazou, T.P.; Gelasakis, A.I.; Chaintoutis, S.C.; Iossifidou, E.G.; Dovas, C.I. Method-Dependent Implications in Foodborne Pathogen Quantification: The Case of Campylobacter coli Survival on Meat as Comparatively Assessed by Colony Count and Viability PCR. Front. Microbiol. 2021, 12, 604933. [Google Scholar] [CrossRef]
- Strain, M.C.; Lada, S.M.; Luong, T.; Rought, S.E.; Gianella, S.; Terry, V.H.; Spina, C.A.; Woelk, C.H.; Richman, D.D. Highly precise measurement of HIV DNA by droplet digital PCR. PLoS ONE 2013, 8, e55943. [Google Scholar] [CrossRef]
- Taylor, S.C.; Carbonneau, J.; Shelton, D.N.; Boivin, G. Optimization of droplet digital PCR from RNA and DNA extracts with direct comparison to RT-qPCR: Clinical implications for quantification of oseltamivir-resistant subpopulations. J. Virol. Methods 2015, 224, 58–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, R.; Paparini, A.; Monis, P.; Ryan, U. Comparison of next-generation droplet digital PCR (ddPCR) with quantitative PCR (qPCR) for enumeration of Cryptosporidium oocysts in faecal samples. Int. J. Parasitol. 2014, 44, 1105–1113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Demeke, T.; Gräfenhan, T.; Holigroski, M.; Fernando, U.; Bamforth, J.; Lee, S.J. Assessment of droplet digital PCR for absolute quantification of genetically engineered OXY235 canola and DP305423 soybean samples. Food Control 2014, 46, 470–474. [Google Scholar] [CrossRef]
- Pecoraro, S. Chapter 5: Digital Polymerase Chain Reaction (dPCR)—General Aspects and Applications. In DNA Techniques to Verify Food Authenticity: Applications in Food Fraud; Burns, M., Foster, L., Walker, M., Eds.; The Royal Society of Chemistry: London, UK, 2020; pp. 63–69. ISBN 978-1-78801-178-5. [Google Scholar]
- Quan, P.L.; Sauzade, M.; Brouzes, E. dPCR: A Technology Review. Sensors 2018, 18, 1271. [Google Scholar] [CrossRef] [Green Version]
- Singh, G.; Sithebe, A.; Enitan, A.M.; Kumari, S.; Bux, F.; Stenström, T.A. Comparison of droplet digital PCR and quantitative PCR for the detection of Salmonella and its application for river sediments. J. Water Health 2017, 15, 505–508. [Google Scholar] [CrossRef]
- Mauvisseau, Q.; Davy-Bowker, J.; Bulling, M.; Brys, R.; Neyrinck, S.; Troth, C.; Sweet, M. Combining ddPCR and environmental DNA to improve detection capabilities of a critically endangered freshwater invertebrate. Sci. Rep. 2019, 9, 14064. [Google Scholar] [CrossRef] [Green Version]
- ISO 16140-2:2016(E); Microbiology of the Food Chain—Method Validation—Part 2: Protocol for the Validation of Alternative (Proprietary) Methods against a Reference Method. International Organization for Standardization: Geneva, Switzerland, 2016.
- ISO 19036:2020; Microbiology of the Food Chain—Estimation of Measurement Uncertainty for Quantitative Determinations. International Organization for Standardization: Geneva, Switzerland, 2020.
- Huber, I.; Pavlovic, M.; Maggipinto, M.; Konrad, R.; Busch, U. Interlaboratory proficiency test using MALDI-TOF MS for identification of food-associated bacteria. Food Anal. Methods 2018, 11, 1068–1075. [Google Scholar] [CrossRef]
- German Federal Institute for Risk Assessment (BfR). Laboratory Protocol of the National Reference Laboratory for Campylobacter. 2021. Available online: https://www.bfr.bund.de/cm/349/preparation-of-quantitative-thermophilic-campylobacter-spp-reference-material.pdf (accessed on 18 April 2022).
- He, Y.; Yao, X.; Gunther, N.W.; Xie, Y.; Tu, S.I.; Shi, X. Simultaneous Detection and Differentiation of Campylobacter jejuni, C. coli, and C. lari in Chickens Using a Multiplex Real-Time PCR Assay. Food Anal. Methods 2010, 3, 321–329. [Google Scholar] [CrossRef]
- LaGier, M.J.; Joseph, L.A.; Passaretti, T.V.; Musser, K.A.; Cirino, N.M. A real-time multiplexed PCR assay for rapid detection and differentiation of Campylobacter jejuni and Campylobacter coli. Mol. Cell. Probes 2004, 18, 275–282. [Google Scholar] [CrossRef]
- Anderson, A.; Pietsch, K.; Zucker, R.; Mayr, A.; Müller-Hohe, E.; Messelhäusser, U.; Sing, A.; Busch, U.; Huber, I. Validation of a Duplex Real-Time PCR for the Detection of Salmonella spp. in Different Food Products. Food Anal. Methods 2011, 4, 259–267. [Google Scholar] [CrossRef]
- dMIQE Group; Huggett, J.F. The Digital MIQE Guidelines Update: Minimum Information for Publication of Quantitative Digital PCR Experiments for 2020. Clin. Chem. 2020, 66, 1012–1029. [Google Scholar] [CrossRef] [PubMed]
- Uhlig, S.; Frost, K.; Colson, B.; Simon, K.; Mäde, D.; Reiting, R.; Gowik, P.; Grohmann, L. QuoData Web Service ‘Validation of Qualitative PCR Methods within a Single Laboratory’. 2015. Available online: https://quodata.de/content/validation-qualitative-pcr-methods-single-laboratory (accessed on 18 April 2022).
- Bundesamts für Verbraucherschutz und Lebensmittelsicherheit (BVL). Guidelines for the Single-Laboratory Validation of Qualitative Real-Time PCR Methods; Bundesamt für Verbraucherschutz und Lebensmittelsicherheit, Federal Office of Consumer Protection and Food Safety: Braunschweig, Germany, 2016. Available online: https://www.bvl.bund.de/SharedDocs/Downloads/07_Untersuchungen/Guidelines%20for%20the%20single%20laboratory.pdf?__blob=publicationFile&v=3 (accessed on 18 April 2022).
- Pecoraro, S.; Berben, G.; Burns, M.; Corbisier, P.; De Giacomo, M.; De Loose, M.; Dagand, E.; Dobnik, D.; Eriksson, R.; Holst-Jensen, A.; et al. Overview and Recommendations for the Application of Digital PCR; JRC Technical Report, European Network of GMO Laboratories (ENGL); EUR 29673 EN; Publications Office of the European Union: Luxembourg, 2019; Available online: https://data.europa.eu/doi/10.2760/31034 (accessed on 18 April 2022)ISBN 978-92-76-00180-5.
- Bland, J.M.; Altman, D.G. Measuring agreement in method comparison studies. Stat. Methods Med. Res. 1999, 8, 135–160. [Google Scholar] [CrossRef] [PubMed]
- Uhlig, S.; Weidner, C.; Colson, B. Statistical Modelling and Experimental Design for the Validation of Droplet Digital PCR Methods. Preprints 2022, 2022040104. [Google Scholar] [CrossRef]
- Huggett, J.F.; Cowen, S.; Foy, C.A. Considerations for digital PCR as an accurate molecular diagnostic tool. Clin. Chem. 2015, 61, 79–88. [Google Scholar] [CrossRef] [Green Version]
- Huggett, J.F.; Foy, C.A.; Benes, V.; Emslie, K.; Garson, J.A.; Haynes, R.; Hellemans, J.; Kubista, M.; Mueller, R.D.; Nolan, T.; et al. The digital MIQE guidelines: Minimum Information for Publication of Quantitative Digital PCR Experiments. Clin. Chem. 2013, 59, 892–902. [Google Scholar] [CrossRef]
- Papić, B.; Pate, M.; Henigman, U.; Zajc, U.; Gruntar, I.; Biasizzo, M.; Ocepek, M.; Kušar, D. New Approaches on Quantification of Campylobacter jejuni in Poultry Samples: The Use of Digital PCR and Real-time PCR against the ISO Standard Plate Count Method. Front. Microbiol. 2017, 8, 331. [Google Scholar] [CrossRef] [Green Version]
- Basu, A.S. Digital Assays Part I: Partitioning Statistics and Digital PCR. SLAS TECHNOL. Transl. Life Sci. Innov. 2017, 22, 369–386. [Google Scholar] [CrossRef] [Green Version]
- Košir, A.B.; Divieto, C.; Pavšič, J.; Pavarelli, S.; Dobnik, D.; Dreo, T.; Bellotti, R.; Sassi, M.P.; Žel, J. Droplet volume variability as a critical factor for accuracy of absolute quantification using droplet digital PCR. Anal. Bioanal. Chem. 2017, 409, 6689–6697. [Google Scholar] [CrossRef] [Green Version]
- Stingl, K.; Knüver, M.T.; Vogt, P.; Buhler, C.; Krüger, N.J.; Alt, K.; Tenhagen, B.A.; Hartung, M.; Schroeter, A.; Ellerbroek, L.; et al. Quo vadis?—Monitoring Campylobacter in Germany. Eur. J. Microbiol. Immunol. 2012, 2, 88–96. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Assay | Primer/Probe | Sequence 5′→3′ | Amplicon Size [bp] | Final Concentration in ddPCR [nM] | Reference |
|---|---|---|---|---|---|
| C. jejuni (target gene hipO 1) | hipO-fw | TGCACCAGTGACTATGAATAACGA | 124 (according to Acc. No. NC_002163.1) | 350 | He et al. [34] |
| hipO-re | TCCAAAATCCTCACTTGCCATT | 350 | |||
| hipO-p | FAM 4—TTGCAACCT*CACTAGCAAAATCCACAGCT—IABkFQ 6,7 | 200 | |||
| C. coli (target gene glyA 2) | glyA-fw | CATATTGTAAAACCAAAGCTTATCGTG | 133 (according to Acc. No. AF136494.1) | 350 | LaGier et al. [35] |
| glyA-re | AGTCCAGCAATGTGTGCAATG | 350 | |||
| glyA-p | FAM 4—TAAGCTCCA*ACTTCATCCGCAATCTCTCTAAATTT—IABkFQ 6,7 | 200 | |||
| Internal PCR control (target gene ntb2 3) | IPC-ntb2-fw | ACCACAATGCCAGAGTGACAAC | 125 | 350 | Anderson et al. [36] |
| IPC-ntb2-re | TACCTGGTCTCCAGCTTTCAGTT | 350 | |||
| IPC-ntb2 probe | HEX 5—CACGCGCAT*GAAGTTAGGGGACCA—IABkFQ 6,7 | 200 |
| Sample | Level and Organic Matter Content of Meat Rinse/mL | Bacterial Contamination Level | C. jejuni/C. coli Live CFU/mL | C. jejuni/C. coli Dead Cells/mL | Biological Replicates |
|---|---|---|---|---|---|
| A1 | Low/12 mg | low | 200 | 2000 | 5 |
| A2 | Low/12 mg | low | 500 | 5000 | 5 |
| A3 | Low/12 mg | medium | 1000 | 10,000 | 5 |
| A4 | Low/12 mg | medium | 2000 | 20,000 | 5 |
| A5 | Low/12 mg | high | 5000 | 50,000 | 5 |
| A6 | Low/12 mg | high | 10,000 | 100,000 | 5 |
| Sample | Matrices | Level and Organic Matter Content of Meat Rinse/mL | Bacterial Contamination Level | C. jejuni/C. coli Live CFU/mL | C. jejuni/C. coli Dead Cells/mL | Biological Replicate |
|---|---|---|---|---|---|---|
| T1-LM | Chicken neck skin/Chicken breast/Turkey skin | Low/8 mg | low | 500 | 5000 | 1 |
| T2-LM | Low/8 mg | low | 1000 | 10,000 | 1 | |
| T2-MM | medium/37–48 mg | low | 1000 | 10,000 | 1 | |
| T3-LM | Low/8 mg | medium | 2000 | 20,000 | 1 | |
| T4-LM | Low/8 mg | medium | 5000 | 50,000 | 1 | |
| T5-LM | Low/8 mg | high | 20,000 | 200,000 | 1 | |
| T5-MM | medium/37–48 mg | high | 20,000 | 200,000 | 1 |
| Factor 1 | Factor 2 | Factor 3 | Factor 4 | Factor 5 | Factor 6 | Factor 7 | |
|---|---|---|---|---|---|---|---|
| Day | Scientist | Mastermix | Time interval: DNA Extraction to generation of droplets | Time interval: Generation of droplets to PCR reaction | Time interval: PCR reaction to Droplet reading | Cartridge for generation of droplets | |
| day 1 vs. day 2 | 2 different Scientists | with vs. without dUTP | immediately vs. 3 nights freezing | immediately vs. after 30 min | immediately vs. overnight 4 °C | batch 1 (Lot. C000112961) vs. batch 2 (Lot. C000114859) | |
| Run 1 | day 1 | 1 | with | immediately | immediately | immediately | batch 1 |
| Run 2 | day 1 | 1 | with | 3 nights | 30 min | overnight | batch 2 |
| Run 3 | day 1 | 2 | without | 3 nights | immediately | immediately | batch 2 |
| Run 4 | day 1 | 2 | without | immediately | 30 min | overnight | batch 1 |
| Run 5 | day 2 | 2 | with | 3 nights | immediately | overnight | batch 1 |
| Run 6 | day 2 | 2 | with | immediately | 30 min | immediately | batch 2 |
| Run 7 | day 2 | 1 | without | immediately | immediately | overnight | batch 2 |
| Run 8 | day 2 | 1 | without | 3 nights | 30 min | immediately | batch 1 |
| Sample | Bacterial Contamination Level | C. jejuni Live CFU/mL | C. jejuni Dead Cells/mL | C. coli Live CFU/mL | C. coli Dead Cells/mL | Total Live Cells/mL | Total Dead Cells/mL | Total Number of Spike for Eight Runs |
|---|---|---|---|---|---|---|---|---|
| TU1 | low | 150 | 750 | 50 | 250 | 200 | 1000 | 26 |
| TU2 | low | 125 | 625 | 375 | 1875 | 500 | 2500 | 26 |
| TU3 | Medium | 250 | 1250 | 750 | 3750 | 1000 | 5000 | 18 |
| TU4 | High | 3750 | 18,750 | 1250 | 6250 | 5000 | 25,000 | 18 |
| TU5 | High | 7500 | 37,500 | 2500 | 12,500 | 10,000 | 50,000 | 18 |
| Sample | Central Value Ref. Method | Central Value Alt. Method | Absolute Bias | Upper β-ETI | Lower β-ETI | Upper AL | Lower AL |
|---|---|---|---|---|---|---|---|
| A1 | 2.25 | 2.34 | 0.09 | 0.21 | −0.03 | +0.50 | −0.50 |
| A2 | 2.55 | 2.75 | 0.20 | 0.32 | 0.08 | ||
| A3 | 2.83 | 3.09 | 0.27 | 0.39 | 0.15 | ||
| A4 | 3.18 | 3.30 | 0.12 | 0.24 | 0.00 | ||
| A5 | 3.66 | 3.71 | 0.05 | 0.17 | −0.07 | ||
| A6 | 3.94 | 4.03 | 0.09 | 0.21 | −0.03 |
| Sample | Central Value Ref. Method | Central Value Alt. Method | Absolute Bias | Upper β-ETI | Lower β-ETI | Upper AL | Lower AL |
|---|---|---|---|---|---|---|---|
| A1 | 2.23 | 2.30 | 0.07 | 0.23 | −0.09 | +0.50 | −0.50 |
| A2 | 2.70 | 2.78 | 0.08 | 0.24 | −0.08 | ||
| A3 | 3.03 | 3.04 | 0.02 | 0.18 | −0.14 | ||
| A4 | 3.36 | 3.46 | 0.10 | 0.26 | −0.06 | ||
| A5 | 3.76 | 3.77 | 0.00 | 0.16 | −0.16 | ||
| A6 | 3.92 | 4.02 | 0.09 | 0.25 | −0.07 |
| Type | Effect | Estimate | |
|---|---|---|---|
| Constant Term | Proportional Term | ||
| Random | Repeatability | 1.039 × 10−1 | 5.699 × 10−2 |
| Run | 4.560 × 10−5 | 2.301 × 10−5 | |
| Factorial | Day | 0.005 | 0.113 |
| Technician | 0.237 | 0.054 | |
| Supermix | 0.270 | 0.055 | |
| Cartridge | 0.051 | 0.025 | |
| Time interval: extraction-droplet | 0.067 | 0.086 | |
| Time interval: droplet-PCR | 0.037 | 0.008 | |
| Time interval: PCR-reading | 0.160 | 0.087 | |
| Living Cells/mL Nominal Value | Day | Cartridge | Time Interval: Extraction to Droplet Generation | Factorial Effects | Recovery Correction | Repeat-Ability | Run | Binomial (Poisson) | Total | Total Minus Binomial |
|---|---|---|---|---|---|---|---|---|---|---|
| 200 | 16.4% | 5.7% | 37.0% | 42.2% | 16.5% | 28.0% | 0.0% | 58.0% | 78.7% | 53.3% |
| 500 | 14.6% | 5.2% | 27.6% | 32.4% | 11.5% | 22.7% | 0.0% | 36.7% | 55.2% | 41.2% |
| 1000 | 13.4% | 4.9% | 20.9% | 25.8% | 8.3% | 19.1% | 0.0% | 25.7% | 42.0% | 33.2% |
| 5000 | 11.8% | 4.5% | 7.6% | 14.8% | 7.7% | 13.5% | 0.0% | 12.1% | 24.7% | 21.5% |
| 10,000 | 11.6% | 4.5% | 4.6% | 13.3% | 10.5% | 12.9% | 0.0% | 9.4% | 23.3% | 21.3% |
| Living Cells/mL Nominal Value | Day | Cartridge | Time Interval: Extraction to Droplet Generation | Factorial Effects | Recovery Correction | Repeat-Ability | Run | Binomial (Poisson) | Total | Total Minus Binomial |
|---|---|---|---|---|---|---|---|---|---|---|
| 200 | 38.2% | 6.9% | 11.4% | 41.2% | 17.8% | 25.2% | 0.0% | 55.1% | 75.4% | 51.5% |
| 500 | 29.5% | 5.3% | 11.4% | 32.6% | 12.6% | 19.7% | 0.0% | 34.8% | 53.1% | 40.1% |
| 1000 | 23.6% | 4.1% | 11.4% | 26.9% | 9.0% | 15.8% | 0.0% | 24.5% | 40.7% | 32.5% |
| 5000 | 13.7% | 1.5% | 11.4% | 18.1% | 5.5% | 8.9% | 0.0% | 11.4% | 23.8% | 20.9% |
| 10,000 | 12.4% | 1.0% | 11.4% | 17.0% | 7.8% | 7.9% | 0.0% | 8.6% | 22.1% | 20.3% |
| Living Cells/mL Nominal Value | Technical Uncertainty (One Replicate) in %RSD | Technical Uncertainty (One Replicate) in log10 | Technical Uncertainty (Two Replicates) in %RSD | Technical Uncertainty (Two Replicates) in log10 |
|---|---|---|---|---|
| 200 | 75.4 | 0.32 | 60.8 | 0.26 |
| 500 | 53.1 | 0.23 | 44.1 | 0.19 |
| 1000 | 40.7 | 0.17 | 34.5 | 0.15 |
| 5000 | 23.8 | 0.10 | 21.1 | 0.09 |
| 10,000 | 22.1 | 0.09 | 19.7 | 0.08 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Govindaswamy, J.; Zeller-Péronnet, V.; Pavlovic, M.; Wirtz, D.; Murr, L.; Thärigen, D.; Colson, B.; Uhlig, S.; Busch, U.; Huber, I. Digital Droplet-PCR for Quantification of Viable Campylobacter jejuni and Campylobacter coli in Chicken Meat Rinses. Appl. Sci. 2022, 12, 5315. https://doi.org/10.3390/app12115315
Govindaswamy J, Zeller-Péronnet V, Pavlovic M, Wirtz D, Murr L, Thärigen D, Colson B, Uhlig S, Busch U, Huber I. Digital Droplet-PCR for Quantification of Viable Campylobacter jejuni and Campylobacter coli in Chicken Meat Rinses. Applied Sciences. 2022; 12(11):5315. https://doi.org/10.3390/app12115315
Chicago/Turabian StyleGovindaswamy, Janani, Véronique Zeller-Péronnet, Melanie Pavlovic, Daniela Wirtz, Larissa Murr, Diana Thärigen, Bertrand Colson, Steffen Uhlig, Ulrich Busch, and Ingrid Huber. 2022. "Digital Droplet-PCR for Quantification of Viable Campylobacter jejuni and Campylobacter coli in Chicken Meat Rinses" Applied Sciences 12, no. 11: 5315. https://doi.org/10.3390/app12115315