
Comparative Tissue Responses of Marine Mollusks on Seasonal Changes in the Northern Adriatic Sea

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Study Site
2.2. Chemicals
2.3. Analytical Methods
2.4. Statistical Analysis
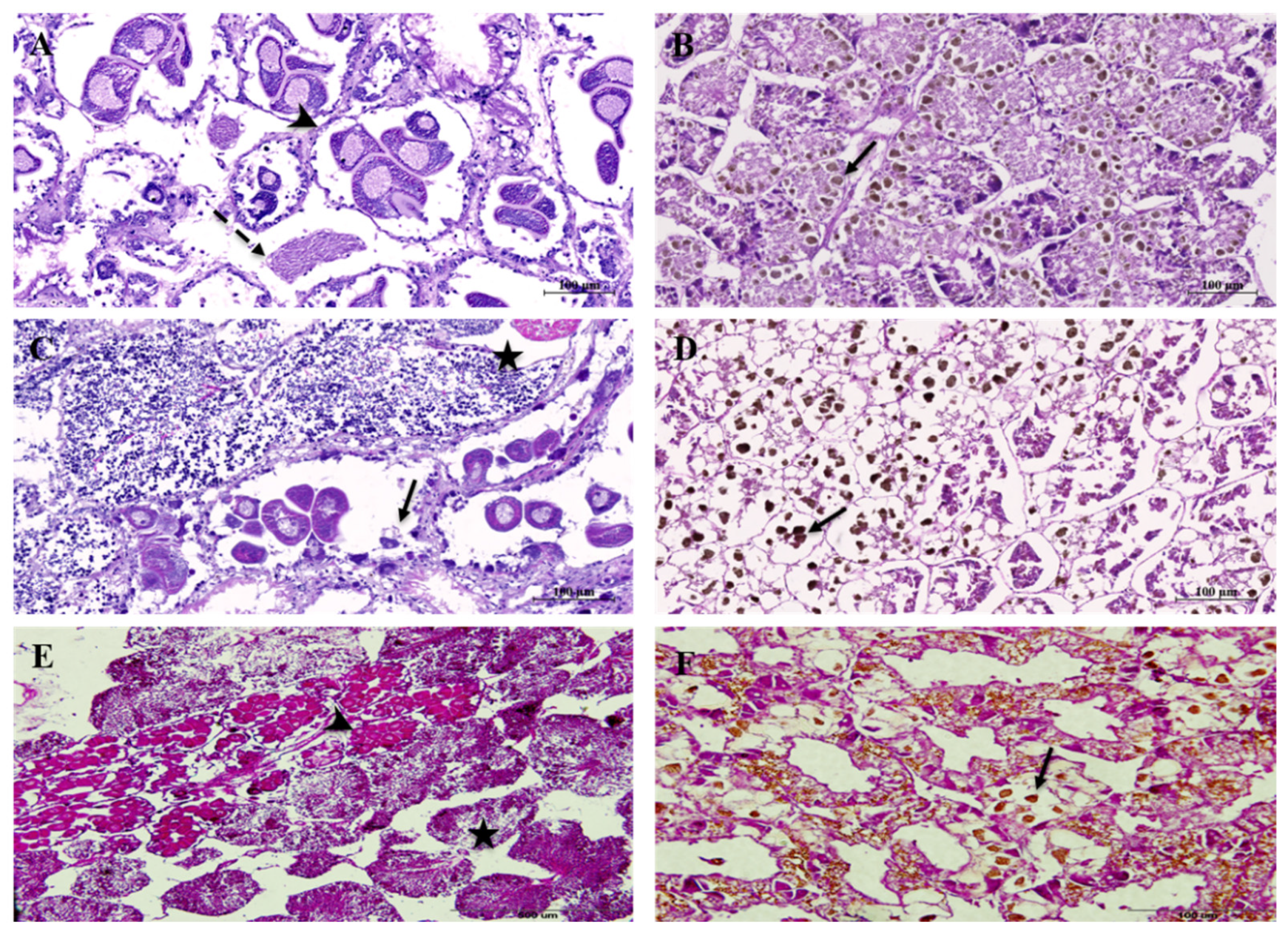
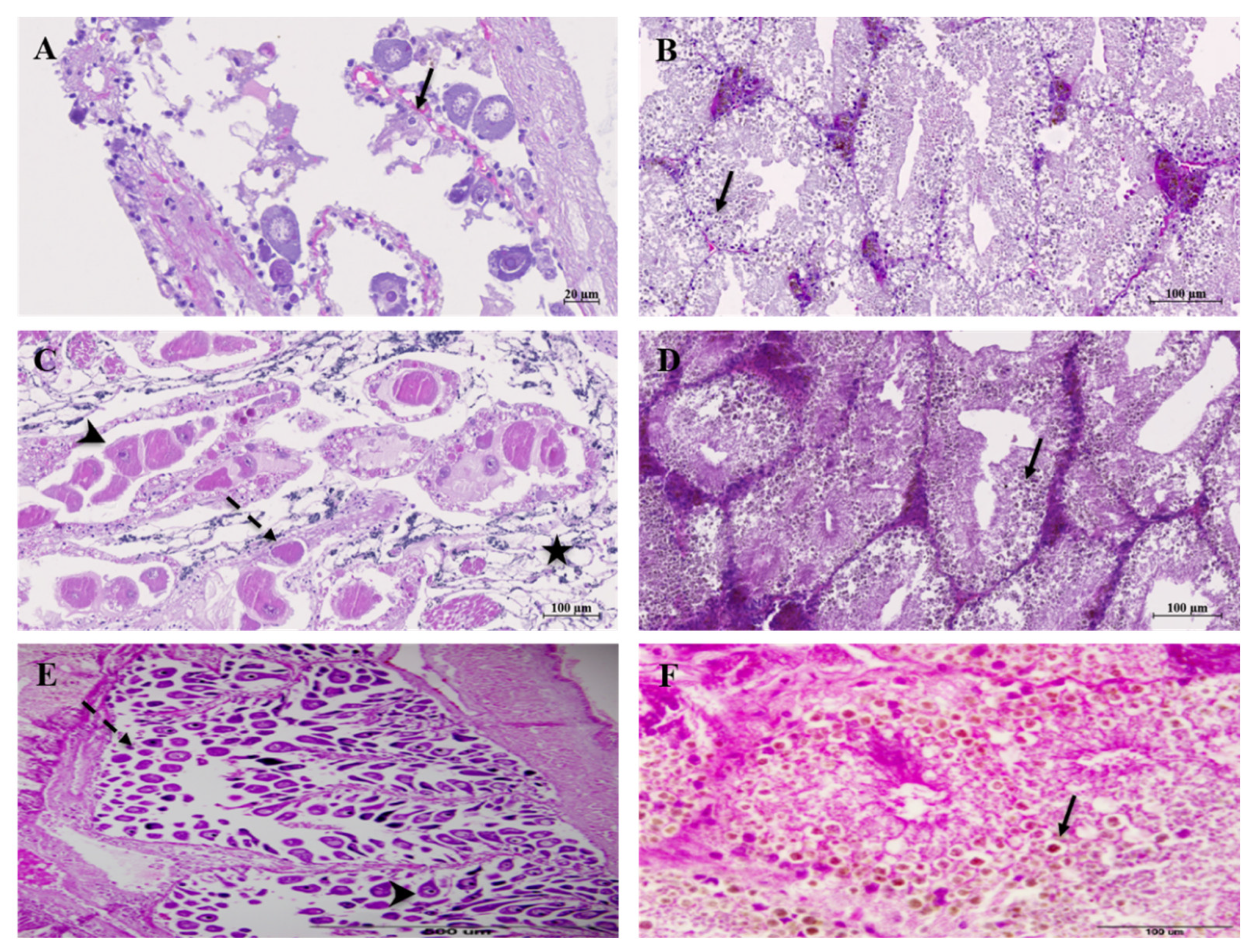
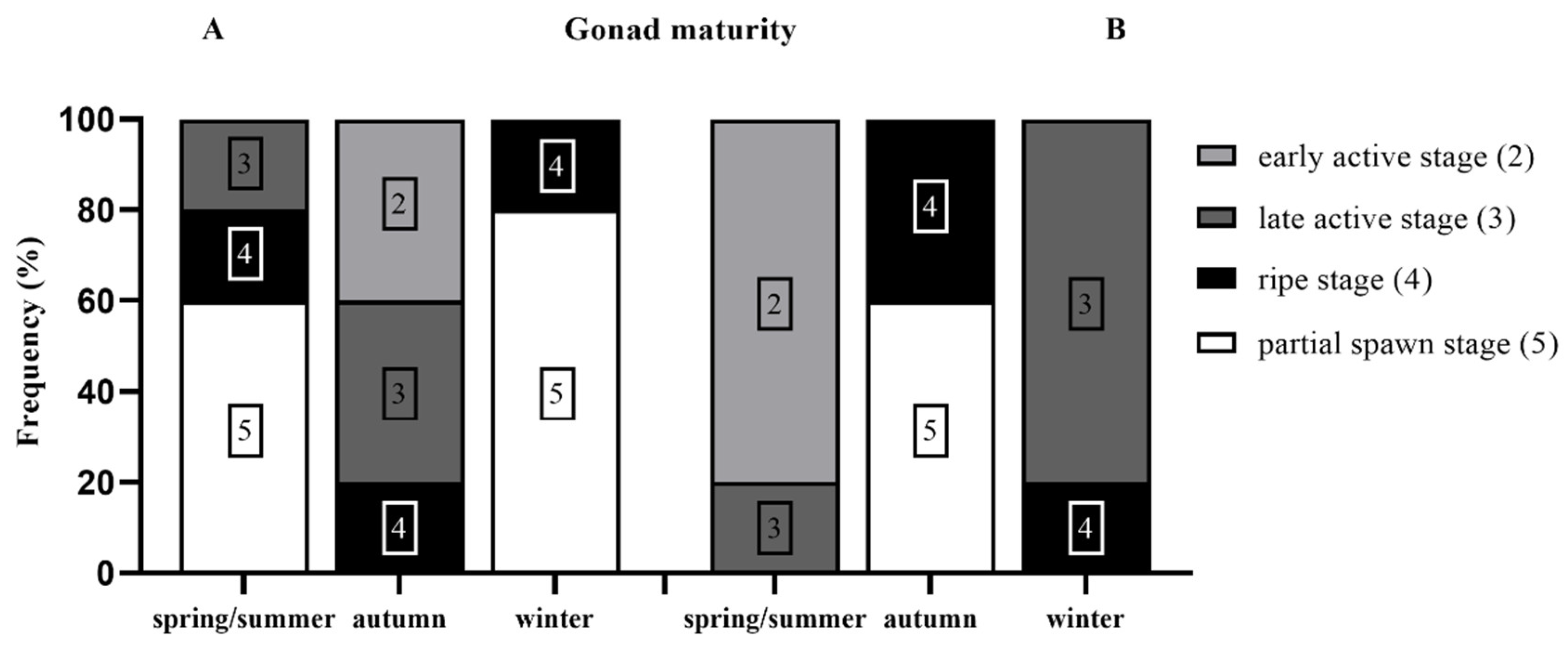
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Eddy, T.D.; Lotze, H.K.; Fulton, E.A.; Coll, M.; Ainsworth, C.H.; De Araújo, J.N.; Bulman, C.M.; Bundy, A.; Christensen, V.; Field, J.C.; et al. Ecosystem effects of invertebrate fisheries. Fish Fish. 2016, 18, 40–53. [Google Scholar] [CrossRef]
- Nerlović, V. Exploitation of scallop Pecten jacobaeus (Linneaus, 1758) in the north western coastal region of Istria. In Proceedings of the 39th Croatian Symposium on Agriculture with International Participation; Žimbrek, T., Ed.; Faculty of Agriculture, University of Zagreb: Zagreb, Croatia, 2004; pp. 606–608. ISBN 953-6135-40-X. [Google Scholar]
- Mgaya, Y.D.; Mercer, J.P. A review of the biology, ecology, fisheries and mariculture of the European abalone Haliotis tuberculata Linnaeus 1758 (Gastropoda: Haliotidae). Biol. Environ. 1994, 94B, 285–304. [Google Scholar]
- Marguš, D.; Teskeredžić, E. The reception of larvae, survival and growth of juvenile Mediterranean scallops (Pecten jacobaeus Linnaeus, 1758) in controlled breeding in the bay of Šarina Draga—The mouth of the Krka River. Croat. J. Fish. 2005, 63, 1–14. (In Croatian) [Google Scholar]
- Mattei, N.; Pellizzato, M. A population study on three stocks of a commercial Adriatic pectinid (Pecten jacobaeus). Fish. Res. 1996, 26, 49–65. [Google Scholar] [CrossRef]
- Peharda, M.; Soldo, A.; Pallaoro, A.; Matić, S.; Cetinić, P. Age and growth of the Mediterranean scallop Pecten jacobaeus (Linnaeus 1758) in the Northern Adriatic Sea. J. Shellfish Res. 2003, 22, 639–642. [Google Scholar]
- Popović, N.T.; Ljubić, B.B.; Strunjak-Perović, I.; Babić, S.; Lorencin, V.; Jadan, M.; Čižmek, L.; Matulić, D.; Bojanić, K.; Čož-Rakovac, R. Seasonal antioxidant and biochemical properties of the Northern Adriatic Pecten jacobaeus. PLoS ONE 2020, 15, e0230539. [Google Scholar] [CrossRef] [PubMed]
- Clavier, J.; Richard, O. Growth of juvenile Haliotis tuberculata (Mollusca: Gastropoda) in their natural environment. J. Mar. Biol. Assoc. UK 1986, 66, 497–503. [Google Scholar] [CrossRef]
- Morash, A.J.; Alter, K. Effects of environmental and farm stress on abalone physiology: Perspectives for abalone aquaculture in the face of global climate change. Rev. Aquac. 2015, 7, 1–27. [Google Scholar] [CrossRef]
- Sokolova, I.M.; Pörtner, H.-O. Metabolic plasticity and critical temperatures for aerobic scope in a eurythermal marine invertebrate (Littorina saxatilis, Gastropoda: Littorinidae) from different latitudes. J. Exp. Biol. 2003, 206, 195–207. [Google Scholar] [CrossRef] [Green Version]
- Clavier, J.; Chardy, P. Investigation into the ecology of the ormer (Haliotis tuberculata L.), factors influencing spatial distribution. Aquat. Living Resour. 1989, 2, 191–197. [Google Scholar] [CrossRef] [Green Version]
- Polimene, L.; Pinardi, N.; Zavatarelli, M.; Colella, S. The Adriatic Sea ecosystem seasonal cycle: Validation of a three-dimensional numerical model. J. Geophys. Res. Space Phys. 2006, 112, C03S19. [Google Scholar] [CrossRef] [Green Version]
- Viličić, D. Specific oceanological characteristics of the Croatian part of the Adriatic. Hrvat. Vode 2014, 22, 297–314. (In Croatian) [Google Scholar]
- Mgaya, Y.D. Synopsis of Biological Data on the European abalone (ormer), Haliotis Tuberculata Linnaeus, 1758 (Gastropoda: Haliotidae); FAO Fisheries Synopsis. No 156; FAO: Rome, Italy, 1995; p. 28. [Google Scholar]
- Matozzo, V.; Chinellato, A.; Munari, M.; Bressan, M.; Marin, M.G. Can the combination of decreased pH and increased temperature values induce oxidative stress in the clam Chemelea gallina and the mussel Mytilus galloprovincialis? Mar. Pollut. Bull. 2013, 72, 34–40. [Google Scholar] [CrossRef]
- Velez, C.; Figueira, E.; Soares, A.M.; Freitas, R. Native and introduced clams biochemical responses to salinity and pH changes. Sci. Total Environ. 2016, 566–567, 260–268. [Google Scholar] [CrossRef] [PubMed]
- Basuyaux, O.; Blin, J.-L.; Costil, K.; Richard, O.; Lebel, J.-M.; Serpentini, A. Assessing the impacts of several algae-based diets on cultured European abalone (Haliotis tuberculata). Aquat. Living Resour. 2018, 31, 28. [Google Scholar] [CrossRef]
- Lau, P.; Wong, H. Effect of size, tissue parts and location on six biochemical markers in the green-lipped mussel, Perna viridis. Mar. Pollut. Bull. 2003, 46, 1563–1572. [Google Scholar] [CrossRef]
- López-Bera, J.; Pueyo, C. Mutagen content and metabolic activation of promutagens by molluscs as biomarkers of marine pollution. Mutat. Res. 1998, 399, 3–15. [Google Scholar] [CrossRef]
- Babić, S.; Barišić, J.; Malev, O.; Klobučar, G.; Topić Popović, N.; Strunjak-Perović, I.; Krasnići, N.; Čož-Rakovac, R.; Sauerborn Klobučar, R. Sewage sludge toxicity assessment using earthworm Eisenia fetida: Can biochemical and histopathological analysis provide fast and accurate insight? Environ. Sci. Pollut. Res. 2016, 23, 12150–12163. [Google Scholar] [CrossRef]
- Narvarte, M.; Kroeck, M. Intraspecific variation in the reproductive cycle of the tehuelchus scallop Aequipecten tehuelchus (Pelecypoda, Pectinidae), in San Matias Gulf, Patagonia, Argentina. J. Shellfish Res. 2002, 21, 571–576. [Google Scholar]
- Najmudeen, T.M. Variation in biochemical composition during gonad maturation of the tropical abaloneHaliotis variaLinnaeus 1758 (Vetigastropoda: Haliotidae). Mar. Biol. Res. 2007, 3, 454–461. [Google Scholar] [CrossRef] [Green Version]
- Borja, A.; Franco, J.; Pérez, V. A Marine Biotic Index to Establish the Ecological Quality of Soft-Bottom Benthos within European Estuarine and Coastal Environments. Mar. Pollut. Bull. 2000, 40, 1100–1114. [Google Scholar] [CrossRef]
- Gallmetzer, I.; Haselmair, A.; Tomašových, A.; Stachowitsch, M.; Zuschin, M. Responses of molluscan communities to centuries of human impact in the northern Adriatic Sea. PLoS ONE 2017, 12, e0180820. [Google Scholar] [CrossRef]
- Abele, D.; Heise, K.; Pörtner, H.O.; Puntarulo, S. Temperature-dependence of mitochondrial function and production of reactive oxygen species in the intertidal mud clam Mya arenaria. J. Exp. Biol. 2002, 205, 1831–1841. [Google Scholar] [PubMed]
- Peck, L.S. Feeding, growth and temperature in the ormer Haliotis tuberculata L. Prog. Underw. Sci. 1989, 14, 95–107. [Google Scholar]
- Viarengo, A.; Canesi, L.; Pertica, M.; Livingstone, D. Seasonal variations in the antioxidant defence systems and lipid peroxidation of the digestive gland of mussels. Comp. Biochem. Physiol. Part C Comp. Pharmacol. 1991, 100, 187–190. [Google Scholar] [CrossRef]
- Xie, J.J.; Chen, X.; Guo, T.Y.; Xie, S.W.; Fang, H.H.; Liu, Z.L.; Zhang, Y.M.; Tian, L.X.; Liu, Y.J.; Niu, J. Dietary values of Forsythia suspensa extract in Penaeus mondon under normal rearing and Vibrio parahaemolyticus 3HP (VP3HP) challenge conditions: Effect on growth, intestinal barrier function, immune response and immune related gene expression. Fish Shellfish Immun. 2018, 75, 316–326. [Google Scholar] [CrossRef]
- Wang, J.; Dong, B.; Yu, Z.-X.; Yao, C.-L. The impact of acute thermal stress on green mussel Perna viridis: Oxidative damage and responses. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2018, 222, 7–15. [Google Scholar] [CrossRef]
- Freas, W.; Grollman, S. Ionic and osmotic influence on prostaglandin release from the gill tissue of a marine bivalve, Modiolus demissus. J. Exp. Biol. 1980, 84, 169–185. [Google Scholar]
- Deridovich, I.; Reunova, O. Prostaglandins: Reproduction control in bivalve molluscs. Comp. Biochem. Physiol. Part A: Physiol. 1993, 104, 23–27. [Google Scholar] [CrossRef]
- Carregosa, V.; Velez, C.; Soares, A.M.V.M.; Figueira, E.; Freitas, R. Physiological and biochemical responses of three Veneridae clams exposed to salinity changes. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2014, 177–178, 1–9. [Google Scholar] [CrossRef]
- Brucka-Jastrzebska, E.; Kawczuga, D.; Grzelak, A.; Bartosz, G. Magnesium content, total antioxidant status and lipid peroxidation in rainbow trout (Onchorhynchus mykiss Walbaum). Magnes. Res. 2009, 22, 273–279. [Google Scholar]
- Chantler, P.D. Scallop Adductor Muscles: Structure and Function. In Scallops: Biology, Ecology, Aquaculture, and Fisheries, 3rd ed.; Shumway, S.E., Parsons, G.J., Eds.; Elsevier: Amsterdam, The Netherlands, 2016; pp. 161–218. [Google Scholar]
- Palacios, E.; Racotta, I.S.; Arjona, O.; Marty, Y.; Le Coz, J.R.; Moal, J.; Samain, J.F. Lipid composition of the pacific lion-paw scallop, Nodipecten subnodosus, in relation to gametogenesis 2. Lipid classes and sterols. Aquaculture 2007, 266, 266–273. [Google Scholar] [CrossRef] [Green Version]
- Pernet, F.; Tremblay, R.; Comeau, L.; Guderley, H. Temperature adaptation in two bivalve species from different thermal habitats: Energetics and remodelling of membrane lipids. J. Exp. Biol. 2007, 210, 2999–3014. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Pita, I.; Sánchez-Lazo, C.; Ruíz-Jarabo, I.; Herrera, M.; Mancera, J.M. Biochemical composition, lipid classes, fatty acids and sexual hormones in the mussel Mytilus galloprovincialis from cultivated populations in south Spain. Aquaculture 2012, 358–359, 274–283. [Google Scholar] [CrossRef]
- Barber, B.J.; Blake, N.J. Reproductive Physiology. In Scallops: Biology, Ecology, Aquaculture, and Fisheries, 3rd ed.; Shumway, S.E., Parsons, G.J., Eds.; Elsevier: Amsterdam, The Netherlands, 2016; pp. 253–299. [Google Scholar]
- Le Pennec, G.; Le Pennec, M.; Beninger, G. Seasonal digestive gland dynamics of the scallop Pecten maximus in the Bay of Brest (France). J. Mar. Biol. Assoc. UK 2001, 81, 663–671. [Google Scholar] [CrossRef]
- Besnard, J.Y.; Lubet, P.; Nouvelot, A. Seasonal variations of the fatty acid content of the neutral lipids and phospholipids in the female gonad of Pecten maximus L. Comp. Biochem. Physiol. 1989, 93B, 21–26. [Google Scholar] [CrossRef]
- Josip, B.; Čož-Rakovac, R.; Delaš, I.; Popović, N.T.; Gavrilović, A.; Jug-Dujaković, J.; Brailo, M.; Sauerborn-Klobučar, R.; Babić, S.; Strunjak-Perović, I. Predictive modeling of European flat oyster (Ostrea edulis L.) fatty acid composition. Aquac. Int. 2017, 25, 805–825. [Google Scholar] [CrossRef]
- Ding, J.; Li, L.; Wu, F.; Zhang, G. Effect of chronic temperature exposure on the immunity of abalone, Haliotis discus hannai. Aquac. Res. 2015, 47, 2861–2873. [Google Scholar] [CrossRef]
- MacDonald, B.A.; Bricelj, V.M.; Shumway, S.E. Physiology: Energy Acquisition and Utilisation. In Scallops: Biology, Ecology, Aquaculture, and Fisheries, 3rd ed.; Shumway, S.E., Parsons, G.J., Eds.; Elsevier: Amsterdam, The Netherlands, 2016; pp. 301–353. [Google Scholar]
- Marchais, V.; Jolivet, A.; Hervé, S.; Roussel, S.; Schöne, B.R.; Grall, J.; Chauvaud, L.; Clavier, J. New tool to elucidate the diet of the ormer Haliotis tuberculata (L.): Digital shell color analysis. Mar. Biol. 2017, 164, 213. [Google Scholar] [CrossRef]
- Jeffries, M.A.; Lee, C.M. A climatology of the northern Adriatic Sea’s response to bora and river forcing. J. Geophys. Res. 2007, 112, 1–18. [Google Scholar] [CrossRef]
- Furlan, E.; Torresan, S.; Critto, A.; Lovato, T.; Solidoro, C.; Lazzari, P.; Marcomini, A. Cumulative Impact Index for the Adriatic Sea: Accounting for interactions among climate and anthropogenic pressures. Sci. Total Environ. 2019, 670, 379–397. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ormer Sampling (Measured at 7 m Depth) | Scallop Sampling (Measured at 15 m Depth) | |||||
|---|---|---|---|---|---|---|
| Fall | Winter | Spring/Summer | Fall | Winter | Spring/Summer | |
| Temperature (°C) | 19.30 ± 3.21 | 11.33 ± 2.11 | 21.20 ± 1.41 | 20.90 ± 1.98 | 12.55 ± 4.88 | 16.87 ± 3.05 |
| Salinity (psu) | 37.97 ± 0.25 | 38.30 ± 0.30 | 34.90 ± 1.56 | 38.10 ± 0.14 | 38.15 ± 0.64 | 36.07 ± 2.02 |
| Dissolved oxygen (mg/L) | 5.15 ± 0.31 | 6.02 ± 0.26 | 5.99 ± 1.31 | 4.99 ± 0.17 | 5.89 ± 0.60 | 5.41 ± 0.32 |
| Green Ormer | Mediterranean Scallop | |||||
|---|---|---|---|---|---|---|
| Fall | Winter | Spring/Summer | Fall | Winter | Spring/Summer | |
| Soft body mass (g) | 17.27 ± 7.32 | 12.58 ± 6.70 | 12.94 ± 8.96 | 21.12 ± 2.55 | 23.17 ± 2.80 | 38.00 ± 8.24 |
| Shell length (mm) | 41.70 ± 6.99 | 41.63 ± 7.54 | 41.71 ± 7.70 | 96.84 ± 9.66 | 102.96 ± 5.12 | 113.93 ± 7.84 |
| Shell mass (g) | 4.88 ± 2.38 | 3.99 ± 2.28 | 4.79 ± 2.35 | 85.41 ± 26.39 | 92.32 ± 10.19 | 97.67 ± 26.48 |
| Green Ormer | Mediterranean Scallop | |||||
|---|---|---|---|---|---|---|
| Fall | Winter | Spring/Summer | Fall | Winter | Spring/Summer | |
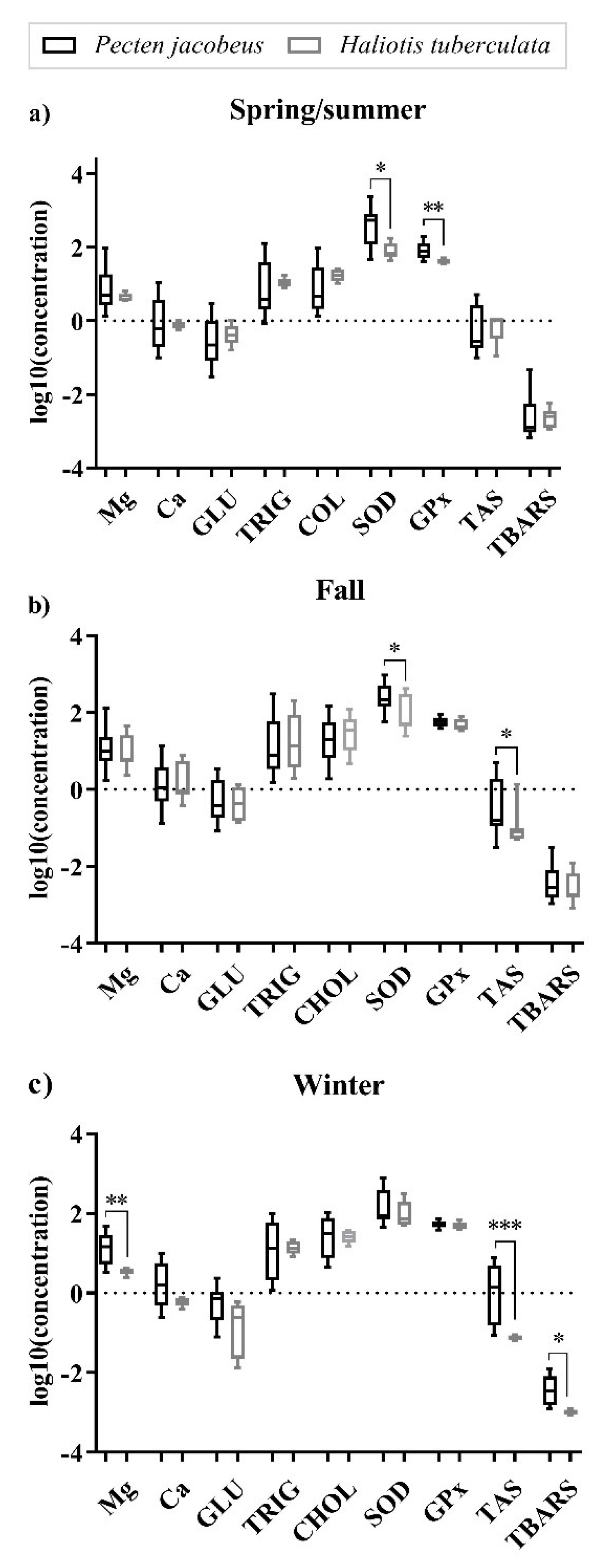
| Mg (mg/g) | 13.89 ± 15.82 | 3.63 ± 0.68 | 4.35 ± 1.15 | 18.00 ± 23.15 | 18.42 ± 13.98 | 13.58 ± 21.03 |
| Ca (mg/g) | 2.44 ± 2.84 | 0.61 ± 0.14 | 0.76 ± 0.19 | 2.93 ± 3.54 | 3.14 ± 3.06 | 2.39 ± 3.11 |
| GLU (mg/g) | 0.62 ± 0.48 | 0.26 ± 0.25 | 0.50 ± 0.33 | 1.00 ± 1.06 | 0.80 ± 0.65 | 0.64 ± 0.77 |
| TRIG (mg/g) | 48.45 ± 75.49 | 14.62 ± 5.20 | 11.34 ± 3.49 | 43.43 ± 67.83 | 31.80 ± 32.98 | 22.54 ± 32.33 |
| CHOL (mg/g) | 44.94 ± 40.95 | 27.44 ± 9.35 | 18.13 ± 6.54 | 31.78 ± 34.06 | 43.07 ± 37.41 | 19.30 ± 24.24 |
| SOD (U/g) | 135.33 ± 159.47 | 121.38 ± 108.55 | 88.31 ± 51.35 | 300.01 ± 236.93 | 208.60 ± 240.49 | 542.20 ± 520.96 |
| GPx (U/g) | 49.01 ± 17.68 | 48.38 ± 11.50 | 41.16 ± 5.80 | 60.69 ± 16.05 | 53.33 ± 10.57 | 93.10 ± 52.38 |
| TAS (mmol/g) | 0.29 ± 0.53 | 0.08 ± 0.01 | 0.76 ± 0.52 | 1.07 ± 1.39 | 2.36 ± 2.70 | 0.52 ± 1.56 |
| TBARS (ng/g) | 4.22 ± 3.79 | 1.05 ± 0.15 | 2.69 ± 1.75 | 5.56 ± 6.67 | 4.99 ± 3.84 | 5.33 ± 8.66 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Topić Popović, N.; Krbavčić, M.; Barišić, J.; Beer Ljubić, B.; Strunjak-Perović, I.; Babić, S.; Lorencin, V.; Matulić, D.; Tomljanović, T.; Čož-Rakovac, R. Comparative Tissue Responses of Marine Mollusks on Seasonal Changes in the Northern Adriatic Sea. Appl. Sci. 2021, 11, 2874. https://doi.org/10.3390/app11062874
Topić Popović N, Krbavčić M, Barišić J, Beer Ljubić B, Strunjak-Perović I, Babić S, Lorencin V, Matulić D, Tomljanović T, Čož-Rakovac R. Comparative Tissue Responses of Marine Mollusks on Seasonal Changes in the Northern Adriatic Sea. Applied Sciences. 2021; 11(6):2874. https://doi.org/10.3390/app11062874
Chicago/Turabian StyleTopić Popović, Natalija, Martina Krbavčić, Josip Barišić, Blanka Beer Ljubić, Ivančica Strunjak-Perović, Sanja Babić, Vanesa Lorencin, Daniel Matulić, Tea Tomljanović, and Rozelindra Čož-Rakovac. 2021. "Comparative Tissue Responses of Marine Mollusks on Seasonal Changes in the Northern Adriatic Sea" Applied Sciences 11, no. 6: 2874. https://doi.org/10.3390/app11062874