1. Introduction
In the physiological setting, the inflammatory responses in the human body are activated to protect the body from internal or external infection, preventing tissue damage. During this process, an intricate set of physiological reactions occur: inflammatory enzymes are activated, inflammatory cytokines released, and inflammatory cells translocated to the site of infection [
1,
2]. However, excessive inflammation results in the destruction of the tissue, accompanied by symptoms of fever, edema, erythema, and pain [
1]. In response to excessive inflammation, macrophages are activated, and they induce high expression and production of inflammatory mediators, such as cycloxygenase-2 (COX-2), inducible nitric oxide synthase (iNOS) nitric oxide (NO), tumor necrosis factor alpha (TNF-α), interleukin 6 (IL-6) and interleukin 1-beta (IL-1β) [
2,
3]. The overproduction of reactive oxygen species (ROS) results in oxidative stress, a known contributor in the development of inflammation, which could lead to various diseases such as unsuccessful aging, cancer, heart conditions, arteriosclerosis, and inflammation [
4].
Okra (
Abelmoschus esculentus L.) is a plant belonging to the Malvaceae family, with its native habitat in Africa. The plant is cultivated in tropical regions in the U.S., Europe, and Asia [
5,
6]. All parts of okra including the pod, the leaf, flowers, and branch, are edible, while the okra pod is especially enriched with polysaccharide, carbohydrates, dietary fibers, proteins, fatty acids, minerals, vitamins, and antioxidants, thereby earning the name “botanical vigor” or “green ginseng” [
5,
7]. Okra pods are known to contain high levels of antioxidant activity, including plentiful phenolic compounds with important physiological activities, including quartering, flavonol induction, and catechin oligomer and hydroxycinnamic acid induction. They also serve as a source of polysaccharides and provide plant-based health benefits including dietary fiber, proteins, fatty acids, minerals, and antioxidants [
7,
8]. Colloids in okra primarily include polysaccharides, flavonoids, polyphenols, alkaloids, and active ingredients for ameliorating fatigue and hypoglycemia, and stimulating immunity [
9]. Okra polysaccharides are mainly comprised of pectins and gum Arabic with rhamnogalacturonan type I pectin as the dominant ingredient [
9]. As they contain high levels of glycoprotein mucins, they facilitate protein digestion, reduce cholesterol, protect the stomach wall, and help neutralize and clear toxins. Soluble fibers in the form of gum and pectin in okra help to reduce serum cholesterol, thereby decreasing the risk of cardiovascular disease, while insoluble fibers help to keep intestinal blood vessels healthy [
8,
10] The okra pod is also known to have beneficial effects against diabetes, colitis, hepatitis, gastric ulcer, as well as neurodegenerative and various chronic diseases [
5,
11,
12]. Despite pieces of evidence on anti-inflammatory effects in vivo or in vitro of raw okra—okra extract in the lipopolysaccharide (LPS)-induced BV2 cell inhibits NO and ROS, the polysaccharide-2 of okra-induced cytokine, okra polysaccharide decrease in cytokines in mouse bone marrow hematopoietic cells [
10,
13,
14]—there are only a few studies on the effects of processing conditions on its anti-inflammatory and antioxidant (ROS assay) properties.
Natural plant extracts have recently received interest for their preventive effects in chronic diseases, especially cardiovascular disease, diabetes, and inflammatory disease [
15,
16,
17], and vegetables and fruits contain a large amount of anti-inflammatory plant chemicals—phytochemicals such as phenolic, triterpenoids, lectin and peptides [
18]. Meanwhile, okra may be eaten raw, alongside other foods as part of salad or soup, or cooked in different ways, such as blanching, pan-frying, and deep-frying [
5]. The nutrients and functional compounds in foods are known to change according to the cooking method used, such as moist, or dry heat cooking [
19,
20]. In line with this, the objective of this study was based on the hypothesis that raw and multimethod cooked okra might have different biological (anti-inflammatory and antioxidant) activities, and four different ethanolic extracts of raw, blanched, steamed, and sous-vide cooked okra were used to investigate the anti-inflammatory and antioxidant effects on lipopolysaccharide- (LPS) or hydrogen peroxide (H
2O
2)-treated RAW264.7 macrophages, by measuring the expression and production of COX-2, iNOS, TNF-α, IL-6, IL-1β, NO, and ROS. Therefore, this study will suggest the different cooking methods on okra and examine suitable cooking methods for consuming okra.
2. Materials and Methods
2.1. Sample Preparation
The raw plant of okra (Abelmoschus esculentus L.) was purchased in Dangjin, Chungcheongnam-do, South Korea. After storage at 5 °C, okra plants of similar sizes were selected for subsequent analyses. The plants were washed in running water and rinsed in distilled water, after which the stalk was removed. For cooking, the methods were selected after a preliminary study with varying cooking times and temperatures.
2.1.1. Blanching
Distilled water (500 mL) was boiled in a pot, and 100 g fresh okra was added and blanched for 2 min. Afterwards, the samples were placed in a plastic bag for rapid cooling under running water (2 min), and the moisture was removed using paper towels. The samples were stored at 5 °C.
2.1.2. Steaming
Distilled water (500 mL) was boiled in a steam cooker (Tefal, Model S07, Groupe SEB-France), to which 100 g okra was added for steaming with a closed lid, and then steamed for 2 min. Afterwards, the samples were treated in the same way as for blanching.
2.1.3. Sous-Vide
Okra (100 g) was placed in a vacuum-sealed polyethylene bag to be sous-vide cooked for 2 min in a bath cooker (Fusion chef, Germany) at 80 °C [
21]. The cooked samples were rapidly cooled under running water in the same ways as for blanching.
2.1.4. Preparation of Okra Extract
Ethanolic extraction of raw, blanched, steamed and sous-vide cooked okra was performed as described by [
5]. Each specimen was frozen and dried for 48 h at −75 to −80 °C with MCFD 8505 (Ilsin Lab Co., Seoul, Korea), and then ground into 30 mesh. The samples (10 g) were extracted by stirring with 100 mL of methanol at 25 °C and 150 rpm for 24 h using a temperature shaker incubator (SI-900R, JEIO TECH, Kimpo, Korea) and then filtered through a Whatman No. 4 paper. The residue was treated again with an additional 100 mL portion of ethanol as described above. The combined ethanolic extracts were evaporated to dryness at 40 °C using a rotary evaporator (NVC-2100, EYELA, Tokyo, Japan) and redissolved in ethanol at a concentration of 50 mg/mL and stored at 4 °C for further use.
2.1.5. Chemicals and Antibodies
Dulbecco’s Modified Eagle’s Medium (DMEM, LM001-05), fetal bovine serum (FBS, S001-01), and a cocktail of penicillin/streptomycin antibiotics (LS202-02) were obtained from WelGENE (Daegu, Korea). An anti-COX-2 antibody (160106) was purchased from Cayman Chemical (Ann Arbor, MI, USA). An anti-iNOS antibody (sc-651) was bought from Santa Cruz Biotechnology (Delaware, CA, USA). An antiactin antibody (A5441) and lipopolysaccharide (LPS) were purchased from Sigma (St. Louis, MO, USA). Western Bright TM enhanced chemiluminescence (ECL) was purchased from Advansta Corporation (San Jose, CA, USA). Plasticwares of 6-, 24-, and 96-well plates, and 60 and 100 mm of cell culture dishes were obtained from SPL Life Sciences (Gyeonggi-do, Korea).
2.2. RAW264.7 Cell Culture
In this study, RAW264.7 macrophages were used. Murine RAW264.7 macrophage (ATCC® TIB71TM, Manasssas, VA, USA) cells were grown and maintained in DMEM media supplemented with 10% heat-inactivated FBS (HI-FBS) and 1% penicillin/streptomycin at 37 °C in a humidified condition of 95% air and 5% CO2. Macrophages secrete various cytokines in the process of phagocytosis and removal of bacteria or foreign substances to regulate immune phenomena, and cells that play a pivotal role in immune action against antigens.
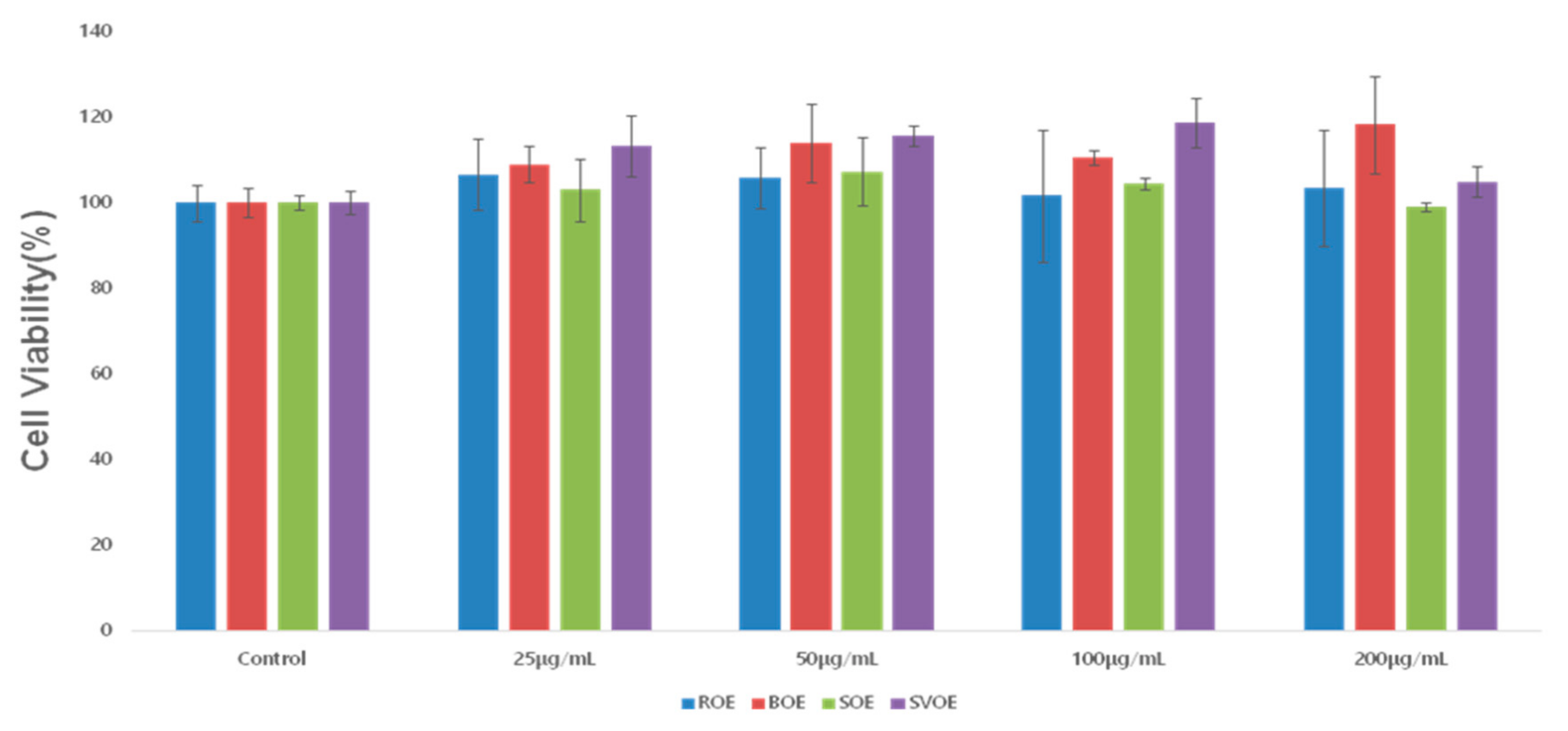
2.3. Cell Viability Assay
To assess the cytotoxicity of the raw okra extract (ROE) and the okra extracts cooked in different ways (BOE for blanching, SOE for steaming, and SVOE for sous-vide cooking) in RAW264.7 cells, the methods of Green et al. [
22]. RAW264.7 macrophage cells were treated with vehicle control (DMSO; 0.1%) or okra ethanol extracts at the indicated concentrations for 24 h. In total, 20 μL of 3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium (MTS) solution was added to each well, and the plates were incubated at 37 °C for 1 h at room temperature (RT). The absorbance of each well was measured at 490 nm using a microplate reader (SPECTRA max 340PC; Molecular Devices, LLC, Silicon Valley, CA, USA).
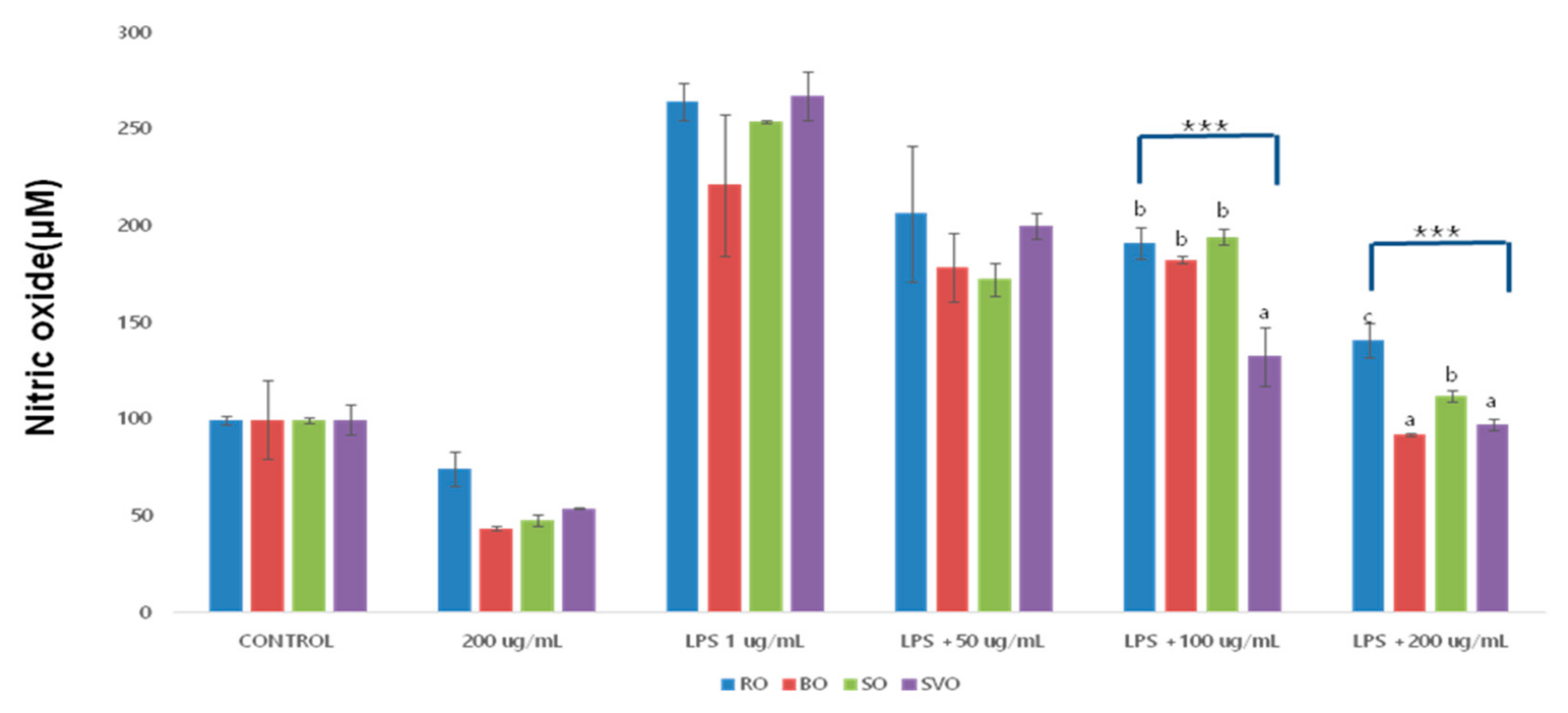
2.4. Nitric Oxide (NO) Assay
To measure the NO production, the methods of Griess Reagent System was used. RAW264.7 macrophage cells were seeded in a 6-well plate (1 × 105 cells/2 mL/ well) overnight. Cells were treated without or with LPS at 1 μg/mL in the presence or absence of differently cooked (raw, blanching, steaming, and sous-vide cooking) okra ethanol extracts at the indicated concentration (50–200 μg/mL) for 8 h. The supernatant from each well was mixed with Griess reagent at 1:1 ratio (0.1% N-1-naphthylenediamine dihydrochloride, 1% sulfanilamide, and 2.5% phosphoric acid in water). After 30 min of incubation at RT, the optical density was measured at 540 nm using a microplate reader (SPECTRA max 340PC; Molecular Devices, LLC, Silicon Valley, CA, USA).
2.5. Preparation of Protein Samples
RAW264.7 macrophage cells were seeded in a 6-well plate (1 × 105 cells/2 mL/well) overnight. Cells were treated with differently cooked (raw, blanching, steaming, and sous-vide cooking) okra ethanol extracts at the designated concentrations (50–200 μg/mL) in the presence or absence of LPS (1 μg/mL) for 8 h. After treatment, cells were washed twice with PBS, and proteins were extracted using modified RIPA buffer [50 mM Tris-Cl (pH 7.4), 150 mM NaCl, 0.1% sodium dodecyl sulfate (SDS), 0.25% sodium deoxycholate, 1% Triton X-100, 1% Nonidet P-40, 1 mM EDTA, 1 mM EGTA and protease inhibitor cocktail (PIC) (1X)]. Whole cell lysates were collected and centrifuged at 12,074× g for 20 min at 4 °C. The supernatant was collected, and its protein concentration was determined by using a bicinchoninic acid protein assay kit (Thermo Fisher Scientific, Inc., Waltham, MA, USA).
2.6. Western Blot Analysis
To determine the effects of the ROE as well as BOE, SOE, and SVOE on COX-2 and iNOS expression, a Western blot analysis was carried out. Proteins (30 μg) were loaded and run in 10 % of SDS-polyacrylamide gel electrophoresis (SDS-PAGE). After separation of proteins, transferred onto polyvinylidene difluoride membrane (PVDF, Millipore, Bedford, MA, USA) and then blocked with 5% (w/v) skim milk in Tris Buffered Saline (TBS) containing 0.1% Tween 20 (TBST) for 1 h at RT. Membranes were probed overnight with antibodies against COX-2 (1:2000), iNOS (1:2000), or actin (1:10,000) at 4 °C, followed with antigoat IgG or antimouse IgG or antirabbit IgG coupled with horseradish peroxidase for 2 h at RT. Later, membranes were rinsed three times with TBST and developed with ECL reagents. Actin expression levels were used as an equal protein loading control.
2.7. Reverse-Transcription-Polymerase Chain Reaction (RT-PCR) Analysis
Total cellular RNA from conditioned RAW264.7 macrophage cells was isolated using TRIzol® reagent (Thermo Fisher Scientific, Inc., Waltham, MA, USA) according to the manufacturer’s protocol and RT PCR was performed. Briefly, equal amounts of total RNA (5 μg) were reverse transcribed in a 40 μL reaction mixture containing 8 μL Molony Murine Leu-kemia Virus Reverse Transcriptase (M MLV RT) 5× buffer, 3 μL 10 mM dNTPs, 0.45 μL 40 U/μL RNase inhibitor, 0.3 μL 200 U/μL M MLV RT (Promega Corporation) and 3.75 μL 20 μM oligo dT (Bioneer Corporation, Oakland, CA, USA). Single-stranded cDNA was amplified by PCR using 4 μL 5× Green Go Taq® Flexi reaction buffer, 0.4 μM 10 mM dNTPs, 0.1 μL 5 U/μL Taq polymerase, 1.2 μL 25 mM MgCl2 (Promega Corporation), and 0.4 μL primer (20 pM/μL). The following primer pairs were used: COX-2 sense, 5′ TTGAAGACCAGGAGTACAGC 3′ and antisense, 5′ GGTACAGTTCCATGACATCG 3′; iNOS sense, 5′ GACAAGCTGCATGTGACATC 3′ and antisense, 5′ GCTGGTAGGTTCCTGTTGTT 3′; TNF-α sense, 5′ GGCAGGTCTACTTTGGAGTCATTGC 3′; antisense, 5′ ACATTCGAGGCTCCAGTGAATTCG 3′; IL-6 sense, 5′ GTCGGAGGCTTAATTACACATGTTC 3′ and antisense, 5′ ACTCCTTCTGTGACTCCAGCTTATC 3′; IL-1β sense, 5′ TGCAGAGTTCCCCAACTGGTACATC 3′ and antisense, 5′ GTGCTGCCTAATGTCCCCTTGAAT 3′; actin sense, 5′ TCATGAAGTGTGACGTTGACATCCGT 3′ and antisense, 5′ CCTAGAAGCATTTGCGGTGCACGATG 3′. The PCR conditions were as follows: For COX-2: 30 cycles of denaturation at 95 °C for 1 min, annealing at 63 °C for 1 min and extension at 72 °C for 1 min; for iNOS: 30 cycles of denaturation at 95 °C for 1 min, annealing at 50 °C for 1 min and extension at 72 °C for 1 min; for TNF-α: 30 cycles of denaturation at 95 °C for 30 s, annealing at 58 °C for 1 min and extension at 72 °C for 30 s; for IL-6: 30 cycles of denaturation at 95 °C for 30 s, annealing at 59 °C for 1 min and extension at 72 °C for 1 min; for IL-1β: 30 cycles of denaturation at 95 °C for 30 s, annealing at 60 °C for 30 s and extension at 72 °C for 30 s; β-actin: 25 cycles of denaturation at 95 °C for 30 s, annealing at 63 °C for 30 s and extension at 72 °C for 1 min. β-actin was used as an internal control to evaluate the relative expression of iNOS, TNF-α, IL-6, and IL-1β.
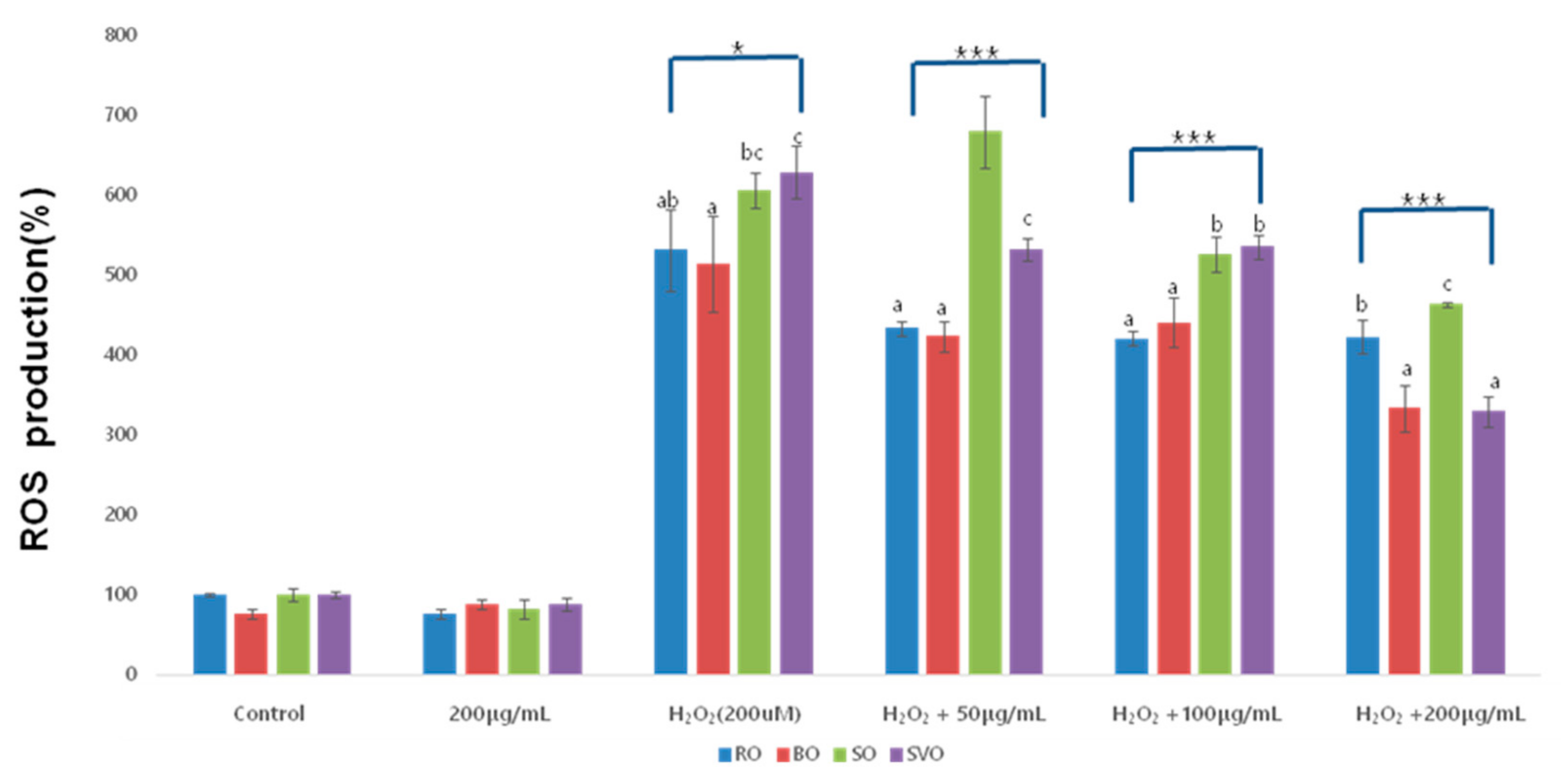
2.8. Reactive Oxygen Species (ROS) Assay
RAW264.7 macrophage cells were seeded in a black-clear bottom 96-well black clear bottom plate (2.5 × 104 cells/100 µL/well) overnight. Cells were incubated with diluted CM-H2DCFDA (10 µM, Abcam, Cambridge, MA, USA) solution for 30 min at 37 °C. After removing CM-H2DCFDA solution, cells were washed with phosphate-buffered saline (PBS), and treated with vehicle control (DMSO; 0.1%) or H2O2 (200 µM) in the presence or absence of differently cooked (raw, blanching, steaming, and sous-vide cooking) okra ethanol extracts at the designated concentrations for 30 min. Fluorescence intensity of DCF was quantified with excitation and emission at 488 and 520 nm, respectively, using Victor3 (Perkin Elmer, CT, USA).
2.9. Statistical Analysis
The results were analyzed using the SPSS 24 (SPSS Inc., Chicago, IL, USA) and one-way ANOVA was performed to analyze the difference in mean values among the experimental groups. Duncan’s multiple range test was used to analyze the inter-variable differences, while the statistical significance was tested at p < 0.05, p < 0.01, and p < 0.001.
4. Discussion
Excessive inflammation and oxidative stress is closely linked to the development of many human chronic illnesses. Up to date, little is known about okra’s anti-inflammatory and antioxidative effects and modes of action. This study investigated the effects of four different okra ethanolic extracts (ROE, BOE, SOE, and SVOE) on regulation of LPS-induced inflammatory enzyme and cytokine expression and H2O2-induced ROS generation in RAW264.7 cells. We report that ROE, BOE, SOE, and SVOE strongly inhibit not only LPS-induced expression of iNOS, TNF-α, IL-1β, and IL-6, but also H2O2-induced ROS generation in RAW264.7 cells.
It has been previously reported that ethanolic extracts of raw okra at ≤200 μg/mL concentrations inhibit LPS-induced expression of TNF-a and IL-1β in BV2 microglia cells with no cytotoxicity, addressing its potential utility against neuro-inflammatory illnesses [
14]. Macrophages are key defensive cells in response to inflammation, producing NO and cytokines to serve as first-line protection against infection. LPSs found in the cell membranes of macrophages have been used to induce inflammation in related experiments [
2,
3]. Nitric oxide (NO), as an indicator of inflammatory response, is produced mainly by inducible nitric oxide synthase (iNOS) and is associated with the virulence of inflammatory diseases [
2,
4,
23]. In this study, we have demonstrated that ethanolic extracts of raw okra (ROE) at ≤200 μg/mL largely suppresses not only LPS-induced expression of iNOS, TNF-α, IL-1β, and IL-6, but also LPS-induced NO production in RAW264.7 cells with no cytotoxicity, supporting ROE’s anti-inflammatory effect. As previously mentioned, the nutrients and functional compounds in foods changed according to the cooking method used, such as moist, or dry heat cooking [
19,
20,
24], which may thus influence their biological activity. Of importance, we herein have shown that other ethanolic extracts of okra (BOE, SOE, and SVOE) prepared from blanching, steaming, and sous-vide cooking methods, respectively, are also able to greatly inhibit LPS-induced iNOS expression and NO production in RAW264.7 cells. These results point out that BOE, SOE, and SVOE also have strong anti-inflammatory activities. Given that SVOE has higher anti-inflammatory (and antioxidant) activities than those of ROE herein, it is suggested that the sous-vide cooking method is useful in not only preserving the nutrients and functional compounds in okra but also improving okra’s biological (anti-inflammatory) activity. There is a wealth of information illustrating presence of a variety of active components in okra (extracts), including polysaccharide [
8,
9,
13]. Because both ROE and SVOE have strong anti-inflammatory (and antioxidant) activities herein, it will be interesting to identify and compare, in future, functional components in ROE and SVOE and elucidate how they trigger anti-inflammatory (antioxidant) activities.
COX-2 and iNOS are absent in normal cells, but can be induced by LPS or inflammatory cytokines, resulting in inflammatory tissue damage, gene mutation, and nerve damage [
25,
26]. The present decrease in COX-2 and iNOS expression by okra extracts from all cooking methods is indicative of okra’s anti-inflammatory effects. LPS induced high iNOS mRNA expression in RAW264.7 cells, pointing out that the iNOS proteins induced by LPS in these cells were due to its transcriptional upregulation. In addition, a concentration-dependent inhibitory effect of okra extracts on the production of iNOS and inflammatory cytokines (TNF-α, IL-6, and IL-1β) was shown across all four cooking methods. TNF-α is a cytokine produced by white blood cells, including lymphocytes and macrophages. It is generated upon stimulation of macrophages, by substances such as LPS, but is not produced in a normal state [
25,
26]. TNF-α plays a critical role in natural immune responses and is associated with the onset and progress of acute and chronic inflammatory diseases [
27,
28]. IL-6 has a crucial role in the early immune response and overexpression could cause various diseases such as infectious diseases, malignant tumors, or autoimmune diseases; hence, IL-6 is always increased in various inflammatory diseases. IL-6 is also important in the functional regulation of T cells and B cells [
29,
30]. IL-1β, on the other hand, promotes T cell activation, B cell maturation, and natural killer cell activity [
31]. The infiltration of inflammatory cells such as lymphocytes and neutrophils is therefore increased in the inflammatory area [
32] and, when facilitated with IL-1β overexpression, results in autoimmune abnormality, pain or fever, suggestive of IL-1β’s crucial role in inflammation. In this study, the okra extracts prepared using all four different cooking methods were shown to have effectively inhibited the production of iNOS, TNF-α, IL-6 and IL-1β, the main inflammatory cytokines, when their levels were measured after treating RAW264.7 cells simultaneously with LPS and different okra extracts. Therefore, they were effective in treating inflammation by effectively controlling cytokines, which play a leading role in the inflammation process. As the okra extracts may affect other molecular pathways, their detail mechanism should be studied further. This effect should be analyzed in various cell lines.
Increase intracellular ROS production in response to inflammation result in apoptosis of causing feedback-inhibition of inflammation; however, in chronic inflammation, prolonged inflammation and oxidative stress a toxic environment exacerbating conditions such as asthma, arthritis, inflammatory bowel disease, and bronchitis. In addition, a high level of ROS causes oxidative stress inducing damage to the lipids, proteins, and DNA [
33,
34]. Therefore, natural material that inhibits high ROS production could be used as potential anti-inflammatory and antioxidant drug or agent. Previous studies have demonstrated okra’s antioxidant property [
9,
10,
11]. In the current study, we have shown the ability of ROE, BOE, SOE, and SVOE to largely lower intracellular ROS levels in LPS-stimulated RAW264.7 cells, illustrating their antioxidant effects. ROE, BOE, SOE, and SVOE have the ability to greatly inhibit LPS-induced NO production in RAW264.7 cells; herein, it is further conceivable that the antioxidant effects of ROE, BOE, SOE, and SVOE are due to the reduced NO production through iNOS downregulation. It is worth stating that among the extracts tested, SVOE—especially at 200 μg/mL—had the highest ROS scavenging activity.
In summary, we have demonstrated firstly that ROE, BOE, SOE, and SVOE at nontoxic concentrations (≤200 μg/mL) have strong anti-inflammatory and antioxidant effects on LPS- or H2O2-treated RAW264.7 cells, and these effects are mediated through the reduced expression and production of iNOS, TNF-α, IL-6, NO, and ROS. This works advocates use of ROE, BOE, SOE, and SVOE as potential anti-inflammatory and antioxidant foods or agents, and sous-vide cooking is the recommended method to prepare okra and related products with improved anti-inflammatory and antioxidant properties.
5. Conclusions
In this study, the anti-inflammatory and antioxidant effects of raw and cooked okra extracts (blanching, steaming, and sous-vide cooking), was verified using LPS- and H2O2-induced RAW264.7 cells. Okra treatment (≤200 μg/mL) did not affect cell viability, confirming it has no cytotoxic effects. It also significantly inhibited both NO production, COX-2, iNOS expression, and the cytokines, TNF-α, IL-6 and IL-1β, confirming its anti-inflammatory properties, with sous-vide cooking having the most pronounced effect. Okra treatment (≤200 μg/mL) also significantly inhibited ROS production, with the sous-vide cooked extract again showing the greatest antioxidant effect. These results point out that BOE, SOE, and SVOE also have strong anti-inflammatory activities. Given that SVOE has higher anti-inflammatory activities and inhibitory ROS production in cells than those of ROE herein, it is suggested that the sous-vide cooking method is useful not only preserving the functional compounds in okra but also improving okra’s biological (anti-inflammatory and antioxidant) activities. In addition, continuous research on the recommendation of appropriate cooking methods and the possibility of food development is needed.








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}