
The Challenging Riddle about the Janus-Type Role of Hsp60 and Related Extracellular Vesicles and miRNAs in Carcinogenesis and the Promises of Its Solution

 , , and
, , and
Abstract
:1. Stress Responses and the Chaperoning System
Hsp60, One of the Most Ancient Anti-Stress Molecules
2. Hsp60 Chaperonopathies
3. Hsp60 in Carcinogenesis

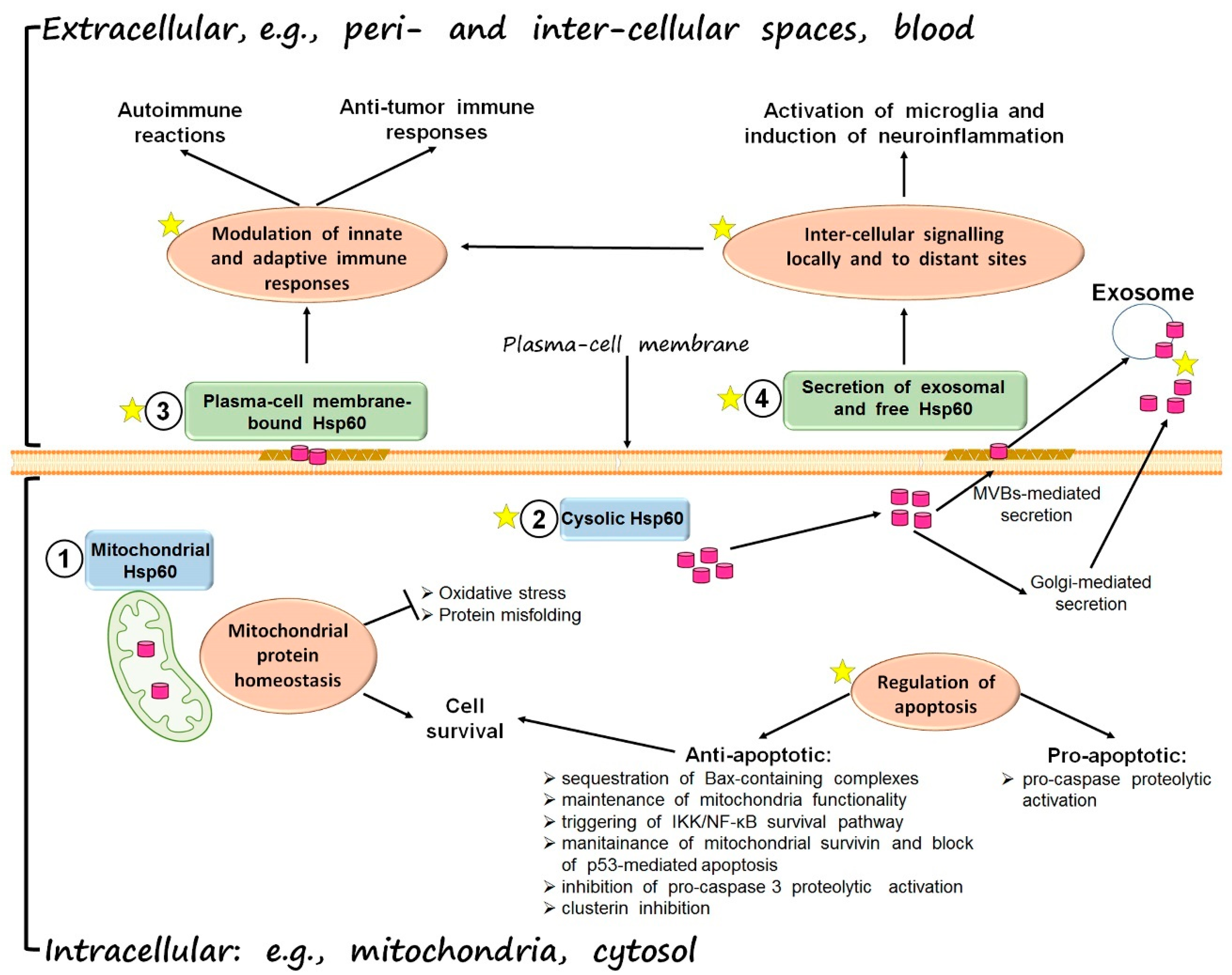{kind=link}
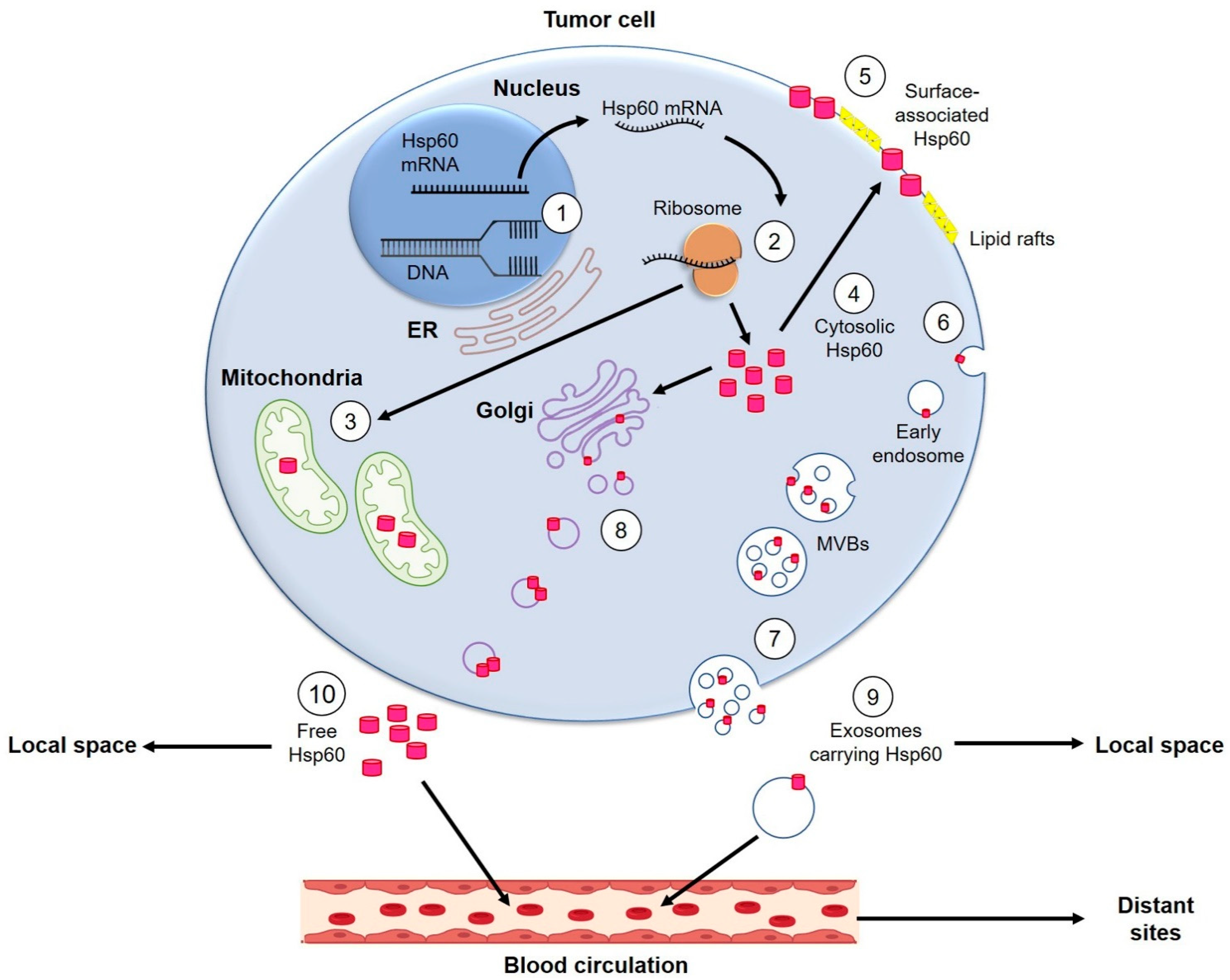{kind=link}
| Cancer | Hsp60 Level, Location and/or Status | Effect | Reference |
|---|---|---|---|
| Lung carcinoma | Presence of Hsp60 on cancer-cell plasma-cell membrane and on membrane of cancer cell-derived exosomes | Possible involvement in cell-to-cell communication and anti-tumor immune response stimulation | [47,48] |
| Decrease of intracellular Hsp60 level and increase of Hsp60 acetylation level after doxorubicin treatment | Hsp60/p53 complex dissociation and restoration of cancer-cells replicative senescence | [69] | |
| Decrease of intracellular Hsp60 level after CubipyOXA treatment | Dissociation of the Hsp60/pro-Caspase-3 complex and cancer-cell apoptosis | [116] | |
| Increased Hsp60 level | Positive correlation with cancer progression and poor prognosis | [94,96] | |
| Oral cancer | Presence of Hsp60 on cell surface | Interaction with gamma-delta T cells and transduction of anti-cancer immune response | [52] |
| Osteosarcoma | Hyperacetylation and loss of mitochondrial Hsp60 after Geldanamycin treatment | Decreased viability and augmented cancer-cell death | [68] |
| Breast cancer | Increase of phosphorylated surface Hsp60 | α3β1 integrin activation and enhancement of cancer cells motility and adhesion | [70] |
| Increased cytosolic Hsp60 | Enhanced cancer-cell proliferation and reduced apoptosis; positive correlation with worse disease-free survival and poor prognosis | [91,92] | |
| Bronchial carcinoma | Decreased Hsp60 level | Positive correlation with bronchial cancer development and progression | [85,119] |
| Cervical cancer | Increased Hsp60 level | Positive correlation with cancer progression and malignancy | [86,87,88,89,90,91,92,93,94,95,96,97,98,99,100,101,102,103,104,105,106,107,108,109,110] |
| Ovarian cancer | Increased Hsp60 level | Positive correlation with cancer progression and severity (poor prognosis and resistance to anti-cancer treatment) | [89,90,105] |
| Colorectal cancer | Increased Hsp60 level | Positive correlation with cancer progression and malignancy | [93,120] |
| IGFBP7-dependent down-regulation of intracellular and extracellular Hsp60 level | Involvement in tumor suppressive activity of IGFBP7 | [108] | |
| Prostate cancer | Increased Hsp60 level | Positive correlation with tumor progression and hormone resistance | [96] |
| Gastric cancers | Increased Hsp60 level | Positive correlation with cancer progression, invasiveness, and poor overall survival. | [97,98] |
| Leukemia | Presence of Hsp60 on the cell surface | Activation/maturation of dendritic cells and generation of potent anti-tumor T-cell response | [53] |
| Increased Hsp60 level | Positive correlation with lower complete remission rate and shorter survival | [101] | |
| Glioblastoma multiforme | Increased Hsp60 level | Cytoprotective and pro-survival role | [102] |
| Decreased Hsp60 level | Reduced cancer cell proliferation and tumor growth | [109,112] | |
| Hepatocellular carcinoma | Decreased Hsp60 level | Positive correlation with cancer progression and poor prognosis | [118] |
| Increased exosomal release of Hsp60 after anti-cancer treatment | Activation of anti-tumor immune response | [121] | |
| Pancreatic cancer | Increased Hsp60 level | Positive correlation with cancer-cell proliferation and tumor growth and progression | [111] |
| Bladder carcinoma | Increased Hsp60 level | Positive correlation with resistance to anti-cancer treatment | [105] |
| Decreased Hsp60 level | Positive correlation with higher tumor stage and cancer recurrence | [122] | |
| Renal cell carcinoma | Decreased Hsp60 | Disruption of mitochondria homeostasis and positive correlation with cancer progression | [117] |
| Large bowel cancer | Increased intracellular and exosomal Hsp60 level | Positive correlation with tumor development and progression | [123] |
| Thyroid cancers | Increased intracellular and exosomal Hsp60 level | Positive correlation with tumor progression | [99,100] |
3.1. Hsp60 in Extracellular Vesicles in Carcinogenesis
3.2. Hsp60 and miRNAs Correlations and Implications for Carcinogenesis
| Tissue | miRNAs and miRNAs Status | Effect | Reference |
|---|---|---|---|
| Rat cardiomyocytes | miR-1 and miR-206 high-glucose-dependent up-regulation | Increased cell apoptosis induced by Hsp60 and IGF-1 down-regulation and IGF-1/IGF-1R/PI3 K/Akt pathway inhibition | [158,159] |
| Breast cancer | miR-29a in vitro down-regulation | Increased cancer cell apoptosis and sensitization to anti-cancer treatment induced by Hsp60 up-regulation | [155] |
| Gastric Lymphoma | miR-17 higher level | Increased malignancy via regulation of Hsp60/TNFR2 pathway | [160] |
| Hepatocellular carcinoma | miR-644a in vitro up-regulation | Increased cancer-cell apoptosis induced by Hsp60 inhibition | [156] |
4. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Brock, T.D.; Freeze, H. Thermus aquaticus gen. n. and sp. n., a nonsporulating extreme thermophile. J. Bacteriol. 1969, 98, 289–297. [Google Scholar] [CrossRef] [Green Version]
- Moseley, B.E.B. Photobiology and Radiobiology of Micrococcus (Deinococcus) radiodurans. In Photochemical and Photobiological Reviews; Springer: Boston, MA, USA, 1983; pp. 223–274. [Google Scholar]
- Herbert, R.A. A perspective on the biotechnological potential of extremophiles. Trends Biotechnol. 1992, 10, 395–402. [Google Scholar] [CrossRef]
- Minton, K.W. DNA repair in the extremely radioresistant bacterium Deinococcus radiodurans. Mol. Microbiol. 1994, 13, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Minton, K.W.; Daly, M.J. A model for repair of radiation-induced DNA double-strand breaks in the extreme radiophile Deinococcus radiodurans. BioEssays 1995, 17, 457–464. [Google Scholar] [CrossRef] [PubMed]
- Gilichinsky, D.; Vishnivetskaya, T.; Petrova, M.; Spirina, E.; Mamykin, V.; Rivkina, E. Bacteria in permafrost. In Psychrophiles: From Biodiversity to Biotechnology; Springer: Berlin/Heidelberg, Germany, 2008; pp. 83–102. ISBN 9783540743347. [Google Scholar]
- Sorokin, D.Y.; Kuenen, J.G.; Muyzer, G. The microbial sulfur cycle at extremely haloalkaline conditions of soda lakes. Front. Microbiol. 2011, 2, 44. [Google Scholar] [CrossRef] [Green Version]
- Dhakar, K.; Pandey, A. Wide pH range tolerance in extremophiles: Towards understanding an important phenomenon for future biotechnology. Appl. Microbiol. Biotechnol. 2016, 100, 2499–2510. [Google Scholar] [CrossRef] [PubMed]
- Macario, A.J.L.; Conway de Macario, E. Stress genes: An introductory overview. Stress 1997, 1, 123–134. [Google Scholar] [CrossRef]
- Hartl, F.U.; Bracher, A.; Hayer-Hartl, M. Molecular chaperones in protein folding and proteostasis. Nature 2011, 475, 324–332. [Google Scholar] [CrossRef]
- Macario, A.J.L.; Conway de Macario, E. Chapter 12—Chaperone proteins and chaperonopathies. In Handbook of Stress, Physiology, Biochemistry, and Pathology; Fink, G., Ed.; Elsevier/Academic Press: Cambridge, MS, USA, 2019; Volume 3, pp. 135–152. Available online: https://doi.org/10.1016/B978-0-12-813146-6.00012-6 (accessed on 18 December 2020).
- Lindquist, S.; Craig, E.A. The heat-shock proteins. Annu. Rev. Genet. 1988, 22, 631–677. [Google Scholar] [CrossRef]
- Becker, J.; Craig, E.A. Heat-shock proteins as molecular chaperones. Eur. J. Biochem. 1994, 219, 11–23. [Google Scholar] [CrossRef]
- Nollen, E.A.A.; Morimoto, R.I. Chaperoning signaling pathways: Molecular chaperones as stress-sensing “heat shock” proteins. J. Cell Sci. 2002, 115, 2809–2816. [Google Scholar]
- Jacob, P.; Hirt, H.; Bendahmane, A. The heat-shock protein/chaperone network and multiple stress resistance. Plant Biotechnol. J. 2017, 15, 405–414. [Google Scholar] [CrossRef] [PubMed]
- Hendrick, J.P.; Hartl, F.U. Molecular chaperone functions of heat-shock proteins. Annu. Rev. Biochem. 1993, 62, 349–384. [Google Scholar] [CrossRef] [PubMed]
- Welch, W.J. Heat shock proteins functioning as molecular chaperones: Their roles in normal and stressed cells. Philos. Trans. R. Soc. B Biol. Sci. 1993, 339, 327–333. [Google Scholar]
- Kampinga, H.H.; Hageman, J.; Vos, M.J.; Kubota, H.; Tanguay, R.M.; Bruford, E.A.; Cheetham, M.E.; Chen, B.; Hightower, L.E. Guidelines for the nomenclature of the human heat shock proteins. Cell Stress Chaperones 2009, 14, 105–111. [Google Scholar] [CrossRef] [Green Version]
- Gupta, R.S. Evolution of the chaperonin families (HSP60, HSP 10 and TCP-1) of proteins and the origin of eukaryotic cells. Mol. Microbiol. 1995, 15, 1–11. [Google Scholar] [CrossRef]
- Rowland, S.E.; Robb, F.T. Structure, Function and Evolution of the Hsp60 Chaperonins; Springer: Singapore, 2017; pp. 3–20. [Google Scholar]
- Vilasi, S.; Bulone, D.; Caruso Bavisotto, C.; Campanella, C.; Marino Gammazza, A.; San Biagio, P.L.; Cappello, F.; Conway de Macario, E.; Macario, A.J.L. Chaperonin of Group I: Oligomeric spectrum and biochemical and biological implications. Front. Mol. Biosci. 2018, 4, 99. [Google Scholar] [CrossRef]
- Georgopoulos, C.P.; Hohn, B. Identification of a host protein necessary for bacteriophage morphogenesis (the groE gene product). Proc. Natl. Acad. Sci. USA 1978, 75, 131–135. [Google Scholar] [CrossRef] [Green Version]
- Barraclough, R.; Ellis, R.J. Protein synthesis in chloroplasts IX. Assembly of newly-synthesized large subunits into ribulose bishopshate carboxylase in isolated intact pea chloroplasts. Biochim. Biophys. Acta 1980, 608, 19–31. [Google Scholar] [CrossRef]
- Hemmingsen, S.M.; Ellis, R.J. Purification and Properties of Ribulosebisphosphate Carboxylase Large Subunit Binding Protein. Plant Physiol. 1986, 80, 269–276. [Google Scholar] [CrossRef] [Green Version]
- Hemmingsen, S.M.; Woolford, C.; Van Der Vies, S.M.; Tilly, K.; Dennis, D.T.; Georgopoulos, C.P.; Hendrix, R.W.; Ellis, R.J. Homologous plant and bacterial proteins chaperone oligomeric protein assembly. Nature 1988, 333, 330–334. [Google Scholar] [CrossRef] [PubMed]
- Horwich, A.L.; Fenton, W.A.; Chapman, E.; Farr, G.W. Two families of chaperonin: Physiology and mechanism. Annu. Rev. Cell Dev. Biol. 2007, 23, 115–145. [Google Scholar] [CrossRef] [PubMed]
- Techtmann, S.M.; Robb, F.T. Archaeal-like chaperonins in bacteria. Proc. Natl. Acad. Sci. USA 2010, 107, 20269–20274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- An, Y.J.; Rowland, S.E.; Na, J.H.; Spigolon, D.; Hong, S.K.; Yoon, Y.J.; Lee, J.H.; Robb, F.T.; Cha, S.S. Structural and mechanistic characterization of an archaeal-like chaperonin from a thermophilic bacterium. Nat. Commun. 2017, 8, 827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spiess, C.; Meyer, A.S.; Reissmann, S.; Frydman, J. Mechanism of the eukaryotic chaperonin: Protein folding in the chamber of secrets. Trends Cell Biol. 2004, 14, 598–604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leitner, A.; Joachimiak, L.A.; Bracher, A.; Mönkemeyer, L.; Walzthoeni, T.; Chen, B.; Pechmann, S.; Holmes, S.; Cong, Y.; Ma, B.; et al. The molecular architecture of the eukaryotic chaperonin TRiC/CCT. Structure 2012, 20, 814–825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willison, K.R. The structure and evolution of eukaryotic chaperonin-containing TCP-1 and its mechanism that folds actin into a protein spring. Biochem. J. 2018, 475, 3009–3034. [Google Scholar] [CrossRef] [PubMed]
- McMullin, T.W.; Hallberg, R.L. A highly evolutionarily conserved mitochondrial protein is structurally related to the protein encoded by the Escherichia coli groEL gene. Mol. Cell. Biol. 1988, 8, 371–380. [Google Scholar] [CrossRef] [Green Version]
- Sigler, P.B.; Xu, Z.; Rye, H.S.; Burston, S.G.; Fenton, W.A.; Horwich, A.L. Structure and function in GroEL-mediated protein folding. Annu. Rev. Biochem. 1998, 67, 581–608. [Google Scholar] [CrossRef] [Green Version]
- Xu, Z.; Sigler, P.B. GroEL/GroES: Structure and function of a two-stroke folding machine. J. Struct. Biol. 1998, 124, 129–141. [Google Scholar] [CrossRef]
- Levy-Rimler, G.; Viitanen, P.; Weiss, C.; Sharkia, R.; Greenberg, A.; Niv, A.; Lustig, A.; Delarea, Y.; Azem, A. The effect of nucleotides and mitochondrial chaperonin 10 on the structure and chaperone activity of mitochondrial chaperonin 60. Eur. J. Biochem. 2001, 268, 3465–3472. [Google Scholar] [CrossRef] [PubMed]
- Vilasi, S.; Carrotta, R.; Mangione, M.R.; Campanella, C.; Librizzi, F.; Randazzo, L.; Martorana, V.; Marino Gammazza, A.; Ortore, M.G.; Vilasi, A.; et al. Human Hsp60 with its mitochondrial import signal occurs in solution as heptamers and tetradecamers remarkably stable over a wide range of concentrations. PLoS ONE 2014, 9, e97657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Enriquez, A.S.; Rojo, H.M.; Bhatt, J.M.; Molugu, S.K.; Hildenbrand, Z.L.; Bernal, R.A. The human mitochondrial Hsp60 in the APO conformation forms a stable tetradecameric complex. Cell Cycle 2017, 16, 1309–1319. [Google Scholar] [CrossRef] [PubMed]
- Viitanen, P.V.; Lorimer, G.; Bergmeier, W.; Weiss, C.; Kessel, M.; Goloubinoff, P. Purification of mammalian mitochondrial chaperonin 60 through in vitro reconstitution of active oligomers. Methods Enzymol. 1998, 290, 203–217. [Google Scholar]
- Nielsen, K.L.; Cowan, N.J. A single ring is sufficient for productive chaperonin-mediated folding in vivo. Mol. Cell 1998, 2, 93–99. [Google Scholar] [CrossRef]
- Nisemblat, S.; Parnas, A.; Yaniv, O.; Azem, A.; Frolow, F. Crystallization and structure determination of a symmetrical “football” complex of the mammalian mitochondrial Hsp60-Hsp10 chaperonins. Acta Crystallogr. Sect. F Struct. Biol. Commun. 2014, 70, 116–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nisemblat, S.; Yaniv, O.; Parnas, A.; Frolow, F.; Azem, A. Crystal structure of the human mitochondrial chaperonin symmetrical football complex. Proc. Natl. Acad. Sci. USA 2015, 112, 6044–6049. [Google Scholar] [CrossRef] [Green Version]
- Gomez-Llorente, Y.; Jebara, F.; Patra, M.; Malik, R.; Nisemblat, S.; Chomsky-Hecht, O.; Parnas, A.; Azem, A.; Hirsch, J.A.; Ubarretxena-Belandia, I. Structural basis for active single and double ring complexes in human mitochondrial Hsp60-Hsp10 chaperonin. Nat. Commun. 2020, 11, 1916. [Google Scholar] [CrossRef] [Green Version]
- Soltys, B.J.; Gupta, R.S. Immunoelectron microscopic localization of the 60-kDa heat shock chaperonin protein (Hsp60) in mammalian cells. Exp. Cell Res. 1996, 222, 16–27. [Google Scholar] [CrossRef]
- Soltys, B.J.; Gupta, R.S. Cell surface localization of the 60 kDa heat shock chaperonin protein (hsp60) in mammalian cells. Cell Biol. Int. 1997, 21, 315–320. [Google Scholar] [CrossRef]
- Soltys, B.J.; Gupta, R.S. Mitochondrial-matrix proteins at unexpected locations: Are they exported? Trends Biochem. Sci. 1999, 24, 174–177. [Google Scholar] [CrossRef]
- Cechetto, J.D.; Soltys, B.J.; Gupta, R.S. Localization of mitochondrial 60-kD heat shock chaperonin protein (Hsp60) in pituitary growth hormone secretory granules and pancreatic zymogen granules. J. Histochem. Cytochem. 2000, 48, 45–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campanella, C.; Bucchieri, F.; Merendino, A.M.; Fucarino, A.; Burgio, G.; Corona, D.F.V.; Barbieri, G.; David, S.; Farina, F.; Zummo, G.; et al. The odyssey of Hsp60 from tumor cells to other destinations includes plasma membrane-associated stages and Golgi and exosomal protein-trafficking modalities. PLoS ONE 2012, 7, e42008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Merendino, A.M.; Bucchieri, F.; Campanella, C.; Marcianò, V.; Ribbene, A.; David, S.; Zummo, G.; Burgio, G.; Corona, D.F.V.; Conway de Macario, E.; et al. Hsp60 is actively secreted by human tumor cells. PLoS ONE 2010, 5, e9247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cappello, F.; Conway de Macario, E.; Marasà, L.; Zummo, G.; Macario, A.J.L. Hsp60 expression, new locations, functions and perspectives for cancer diagnosis and therapy. Cancer Biol. Ther. 2008, 7, 801–809. [Google Scholar] [CrossRef] [PubMed]
- Henderson, B.; Fares, M.A.; Lund, P.A. Chaperonin 60: A paradoxical, evolutionarily conserved protein family with multiple moonlighting functions. Biol. Rev. 2013, 88, 955–987. [Google Scholar] [CrossRef]
- Caruso Bavisotto, C.; Alberti, G.; Vitale, A.M.; Paladino, L.; Campanella, C.; Rappa, F.; Gorska, M.; Conway de Macario, E.; Cappello, F.; Macario, A.J.L.; et al. Hsp60 Post-translational modifications: Functional and pathological consequences. Front. Mol. Biosci. 2020, 7, 95. [Google Scholar] [CrossRef]
- Laad, A.D.; Thomas, M.L.; Fakih, A.R.; Chiplunkar, S.V. Human gamma delta T cells recognize heat shock protein-60 on oral tumor cells. Int. J. Cancer 1999, 80, 709–714. [Google Scholar] [CrossRef]
- Feng, H.; Zeng, Y.; Graner, M.W.; Katsanis, E. Stressed apoptotic tumor cells stimulate dendritic cells and induce specific cytotoxic T cells. Blood 2002, 100, 4108–4115. [Google Scholar] [CrossRef] [Green Version]
- Osterloh, A.; Meier-Stiegen, F.; Veit, A.; Fleischer, B.; Von Bonin, A.; Breloer, M. Lipopolysaccharide-free heat shock protein 60 activates T cells. J. Biol. Chem. 2004, 279, 47906–47911. [Google Scholar] [CrossRef] [Green Version]
- Chandra, D.; Choy, G.; Tang, D.G. Cytosolic accumulation of HSP60 during apoptosis with or without apparent mitochondrial release: Evidence that its pro-apoptotic or pro-survival functions involve differential interactions with caspase-3. J. Biol. Chem. 2007, 282, 31289–31301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samali, A.; Cai, J.; Zhivotovsky, B.; Jones, D.P.; Orrenius, S. Presence of a pre-apoptotic complex of pro-caspase-3, Hsp60 and Hsp10 in the mitochondrial fraction of Jurkat cells. EMBO J. 1999, 18, 2040–2048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xanthoudakis, S.; Sophie, R.; Rasper, D.; Hennessey, T.; Aubin, Y.; Cassady, R.; Tawa, P.; Ruel, R.; Rosen, A.; Nicholson, D.W. Hsp60 accelerates the maturation of pro-caspase-3 by upstream activator proteases during apoptosis. EMBO J. 1999, 18, 2049–2056. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shan, Y.X.; Liu, T.J.; Su, H.F.; Samsamshariat, A.; Mestril, R.; Wang, P.H. Hsp10 and Hsp60 modulate Bcl-2 family and mitochondria apoptosis signaling induced by doxorubicin in cardiac muscle cells. J. Mol. Cell. Cardiol. 2003, 35, 1135–1143. [Google Scholar] [CrossRef]
- Gupta, S.; Knowlton, A.A. HSP60, Bax, apoptosis and the heart. J. Cell. Mol. Med. 2005, 9, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Lin, K.M.; Lin, B.; Lian, I.Y.; Mestril, R.; Scheffler, I.E.; Dillmann, W.H. Combined and individual mitochondrial HSP60 and HSP10 expression in cardiac myocytes protects mitochondrial function and prevents apoptotic cell deaths induced by simulated ischemia-reoxygenation. Circulation 2001, 103, 1787–1792. [Google Scholar] [CrossRef] [Green Version]
- Chun, J.N.; Choi, B.; Lee, K.W.; Lee, D.J.; Kang, D.H.; Lee, J.Y.; Song, I.S.; Kim, H.I.; Lee, S.H.; Kim, H.S.; et al. Cytosolic Hsp60 Is Involved in the NF-kB-Dependent Survival of Cancer Cells via IKK Regulation. PLoS ONE 2010, 5, e9422. [Google Scholar] [CrossRef]
- Ghosh, J.C.; Dohi, T.; Byoung, H.K.; Altieri, D.C. Hsp60 regulation of tumor cell apoptosis. J. Biol. Chem. 2008, 283, 5188–5194. [Google Scholar] [CrossRef] [Green Version]
- Cappello, F.; Marino Gammazza, A.; Palumbo Piccionello, A.; Campanella, C.; Pace, A.; Conway de Macario, E.; Macario, A.J.L. Hsp60 chaperonopathies and chaperonotherapy: Targets and agents. Expert Opin. Ther. Targets 2014, 18, 185–208. [Google Scholar] [CrossRef]
- Macario, A.J.L.; Conway de Macario, E. Sick chaperones, cellular stress, and disease. N. Engl. J. Med. 2005, 353, 1489–1501. [Google Scholar] [CrossRef]
- Macario, A.J.L.; Conway de Macario, E. Chaperonopathies by defect, excess, or mistake. Ann. N. Y. Acad. Sci. 2007, 1113, 178–191. [Google Scholar] [CrossRef] [PubMed]
- Macario, A.J.L.; Conway de Macario, E.; Cappello, F. The Chaperonopathies. Diseases with Defective Molecular Chaperones; Springer: Dordrecht, The Netherlands; Heidelberg, Germany; New York, NY, USA; London, UK, 2013; Available online: https://www.springer.com/us/book/9789400746664 (accessed on 18 December 2020).
- Baron, B. Role of the Post-translational Modifications of HSP60 in Disease. In Heat Shock Protein 60 in Human Diseases and Disorder; Springer: Cham, Switzerland, 2019; Volume 18, pp. 69–94. [Google Scholar]
- Gorska, M.; Marino Gammazza, A.; Zmijewski, M.A.; Campanella, C.; Cappello, F.; Wasiewicz, T.; Kuban-Jankowska, A.; Daca, A.; Sielicka, A.; Popowska, U.; et al. Geldanamycin-Induced Osteosarcoma Cell Death Is Associated with Hyperacetylation and Loss of Mitochondrial Pool of Heat Shock Protein 60 (Hsp60). PLoS ONE 2013, 8, e71135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marino Gammazza, A.; Campanella, C.; Barone, R.; Caruso Bavisotto, C.; Gorska, M.; Wozniak, M.; Carini, F.; Cappello, F.; D’Anneo, A.; Lauricella, M.; et al. Doxorubicin anti-tumor mechanisms include Hsp60 post-translational modifications leading to the Hsp60/p53 complex dissociation and instauration of replicative senescence. Cancer Lett. 2017, 385, 75–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barazi, H.O.; Zhou, L.; Templeton, N.S.; Krutzsch, H.C.; Roberts, D.D. Identification of heat shock protein 60 as a molecular mediator of alpha 3 beta 1 integrin activation. Cancer Res. 2002, 62, 1541–1548. [Google Scholar]
- Leung, W.-H.; Vong, Q.P.; Lin, W.; Bouck, D.; Wendt, S.; Sullivan, E.; Li, Y.; Bari, R.; Chen, T.; Leung, W. PRL-3 Mediates the Protein Maturation of ULBP2 by Regulating the Tyrosine Phosphorylation of HSP60. J. Immunol. 2015, 194, 2930–2941. [Google Scholar] [CrossRef] [Green Version]
- Direskeneli, H.; Ekşioǧlu-Demiralp, E.; Yavuz, Ş.; Ergun, T.; Shinnick, T.; Lehner, T.; Akoǧlu, T. T cell responses to 60/65 kDa heat shock protein derived peptides in Turkish patients with Behcet’s disease. J. Rheumatol. 2000, 27, 708–713. [Google Scholar]
- Shimizu, J.; Izumi, T.; Suzuki, N. Aberrant activation of heat shock protein 60/65 reactive T cells in patients with Behcet’s disease. Autoimmune Dis. 2012, 1, 105205. [Google Scholar] [CrossRef] [Green Version]
- Horváth, L.; Cervenak, L.; Oroszlán, M.; Prohászka, Z.; Uray, K.; Hudecz, F.; Baranyi, É.; Madácsy, L.; Singh, M.; Romics, L.; et al. Antibodies against different epitopes of heat-shock protein 60 in children with type 1 diabetes mellitus. Immunol. Lett. 2002, 80, 155–162. [Google Scholar] [CrossRef]
- Dieudé, M.; Senécal, J.L.; Raymond, Y. Induction of endothelial cell apoptosis by heat-shock protein 60-reactive antibodies from anti-endothelial cell autoantibody-positive systemic lupus erythematosus patients. Arthritis Rheum. 2004, 50, 3221–3231. [Google Scholar] [CrossRef]
- Jamin, C.; Dugué, C.; Alard, J.É.; Jousse, S.; Saraux, A.; Guillevin, L.; Piette, J.C.; Youinou, P. Induction of endothelial cell apoptosis by the binding of anti-endothelial cell antibodies to Hsp60 in vasculitis-associated systemic autoimmune diseases. Arthritis Rheum. 2005, 52, 4028–4038. [Google Scholar] [CrossRef]
- Alard, J.E.; Dueymes, M.; Youinou, P.; Jamin, C. Modulation of endothelial cell damages by anti-Hsp60 autoantibodies in systemic autoimmune diseases. Autoimmun. Rev. 2007, 6, 438–443. [Google Scholar] [CrossRef] [PubMed]
- Alard, J.E.; Dueymes, M.; Youinou, P.; Jamin, C. HSP60 and anti-HSP60 antibodies in vasculitis: They are two of a kind. Clin. Rev. Allergy Immunol. 2008, 35, 66–71. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Quyyumi, A.A.; Rott, D.; Csako, G.; Wu, H.; Halcox, J.; Epstein, S.E. Antibodies to human heat-shock protein 60 are associated with the presence and severity of coronary artery disease: Evidence for an autoimmune component of atherogenesis. Circulation 2001, 103, 1071–1075. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benagiano, M.; D’Elios, M.M.; Amedei, A.; Azzurri, A.; van der Zee, R.; Ciervo, A.; Rombolà, G.; Romagnani, S.; Cassone, A.; Del Prete, G. Human 60-kDa Heat Shock Protein Is a Target Autoantigen of T Cells Derived from Atherosclerotic Plaques. J. Immunol. 2005, 174, 6509–6517. [Google Scholar] [CrossRef] [Green Version]
- Mandal, K.; Foteinos, G.; Jahangiri, M.; Xu, Q. Role of antiheat shock protein 60 autoantibodies in atherosclerosis. Lupus 2005, 14, 742–746. [Google Scholar] [CrossRef]
- Bodolay, E.; Prohászka, Z.; Paragh, G.; Csipő, I.; Nagy, G.; Laczik, R.; Demeter, N.; Zöld, E.; Nakken, B.; Szegedi, G.; et al. Increased levels of anti-heat-shock protein 60 (anti-Hsp60) indicate endothelial dysfunction, atherosclerosis and cardiovascular diseases in patients with mixed connective tissue disease. Immunol. Res. 2014, 60, 50–59. [Google Scholar] [CrossRef]
- Mantej, J.; Polasik, K.; Piotrowska, E.; Tukaj, S. Autoantibodies to heat shock proteins 60, 70, and 90 in patients with rheumatoid arthritis. Cell Stress Chaperones 2019, 24, 283–287. [Google Scholar] [CrossRef]
- Meng, Q.; Li, B.X.; Xiao, X. Toward developing chemical modulators of Hsp60 as potential therapeutics. Front. Mol. Biosci. 2018, 5, 35. [Google Scholar] [CrossRef] [Green Version]
- Cappello, F.; Di Stefano, A.; D’Anna, S.E.; Donner, C.F.; Zummo, G. Immunopositivity of heat shock protein 60 as a biomarker of bronchial carcinogenesis. Lancet Oncol. 2005, 6, 816. [Google Scholar] [CrossRef] [Green Version]
- Cappello, F.; Bellafiore, M.; Palma, A.; Marciano, V.; Martorana, G.; Belfiore, P.; Martorana, A.; Farina, F.; Zummo, G.; Bucchieri, F. Expression of 60-kD Heat Shock Protein Increases during Carcinogenesis in the Uterine Exocervix. Pathobiology 2002, 70, 83–88. [Google Scholar] [CrossRef]
- Castle, P.E.; Ashfaq, R.; Ansari, F.; Muller, C.Y. Immunohistochemical evaluation of heat shock proteins in normal and preinvasive lesions of the cervix. Cancer Lett. 2005, 229, 245–252. [Google Scholar] [CrossRef] [PubMed]
- Hwang, Y.J.; Lee, S.P.; Kim, S.Y.; Choi, Y.H.; Kim, M.J.; Lee, C.H.; Lee, J.Y.; Kim, D.Y. Expression of heat shock protein 60 kDa is upregulated in cervical cancer. Yonsei Med. J. 2009, 50, 399–406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hjerpe, E.; Egyhazi, S.; Carlson, J.; Stolt, M.F.; Schedvins, K.; Johansson, H.; Shoshan, M.; Åvall-Lundqvist, E. HSP60 predicts survival in advanced serous ovarian cancer. Int. J. Gynecol. Cancer 2013, 23, 448–455. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Li, X.; Zhang, W.; Chen, Y.; Zhu, S.; Chen, L.; Xu, R.; Lv, Y.; Wu, D.; Guo, M.; et al. HSP60-regulated Mitochondrial Proteostasis and Protein Translation Promote Tumor Growth of Ovarian Cancer. Sci. Rep. 2019, 9, 12628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bodoor, K.; Abu-Sheikha, A.; Matalka, I.; Alzou’bi, H.; Batiha, O.; Abu-Awad, A.; Jalboush, S.A.; Fayyad, L.M.; Qadiri, E.; Jarun, Y.; et al. Immunohistochemical analysis of heat shock proteins in triple negative breast cancer: HSP60 expression is a marker of poor prognosis. Eur. J. Gynaecol. Oncol. 2018, 39, 926–934. [Google Scholar]
- Kim, S.K.; Kim, K.; Ryu, J.W.; Ryu, T.Y.; Lim, J.H.; Oh, J.H.; Min, J.K.; Jung, C.R.; Hamamoto, R.; Son, M.Y.; et al. The novel prognostic marker, EHMT2, is involved in cell proliferation via HSPD1 regulation in breast cancer. Int. J. Oncol. 2019, 54, 65–76. [Google Scholar] [CrossRef] [Green Version]
- Cappello, F.; Bellafiore, M.; Palma, A.; David, S.; Marcianò, V.; Bartolotta, T.; Sciumè, C.; Modica, G.; Farina, F.; Zummo, G.; et al. 60 KDa chaperonin (HSP60) is over-expressed during colorectal carcinogenesis. Eur. J. Histochem. 2003, 47, 105–109. [Google Scholar] [CrossRef]
- Xu, X.; Wang, W.; Shao, W.; Yin, W.; Chen, H.; Qiu, Y.; Mo, M.; Zhao, J.; Deng, Q.; He, J. Heat shock protein-60 expression was significantly correlated with the prognosis of lung adenocarcinoma. J. Surg. Oncol. 2011, 104, 598–603. [Google Scholar] [CrossRef]
- Aǧababaoǧlu, I.; Önen, A.; Demir, A.B.; Aktaş, S.; Altun, Z.; Ersöz, H.; Şanlii, A.; Özdemir, N.; Akkoçlu, A. Chaperonin (HSP60) and annexin-2 are candidate biomarkers for non-small cell lung carcinoma. Medicine 2017, 96, e5903. [Google Scholar] [CrossRef]
- Castilla, C.; Congregado, B.; Conde, J.M.; Medina, R.; Torrubia, F.J.; Japn, M.A.; Sáez, C. Immunohistochemical expression of Hsp60 correlates with tumor progression and hormone resistance in prostate cancer. Urology 2010, 76, 1017.e1–1017.e6. [Google Scholar] [CrossRef]
- Giaginis, C.; Daskalopoulou, S.S.; Vgenopoulou, S.; Sfiniadakis, I.; Kouraklis, G.; Theocharis, S.E. Heat Shock Protein-27, -60 and -90 expression in gastric cancer: Association with clinicopathological variables and patient survival. BMC Gastroenterol. 2009, 9, 14. [Google Scholar] [CrossRef] [Green Version]
- Li, X.S.; Xu, Q.; Fu, X.Y.; Luo, W.S. Heat shock protein 60 overexpression is associated with the progression and prognosis in gastric cancer. PLoS ONE 2014, 9, e107507. [Google Scholar] [CrossRef] [PubMed]
- Pitruzzella, A.; Paladino, L.; Vitale, A.M.; Martorana, S.; Cipolla, C.; Graceffa, G.; Cabibi, D.; David, S.; Fucarino, A.; Bucchieri, F.; et al. Quantitative immunomorphological analysis of heat shock proteins in thyroid follicular adenoma and carcinoma tissues reveals their potential for differential diagnosis and points to a role in carcinogenesis. Appl. Sci. 2019, 9, 4324. [Google Scholar] [CrossRef] [Green Version]
- Caruso Bavisotto, C.; Cipolla, C.; Graceffa, G.; Barone, R.; Bucchieri, F.; Bulone, D.; Cabibi, D.; Campanella, C.; Marino Gammazza, A.; Pitruzzella, A.; et al. Immunomorphological pattern of molecular chaperones in normal and pathological thyroid tissues and circulating exosomes: Potential use in clinics. Int. J. Mol. Sci. 2019, 20, 4496. [Google Scholar] [CrossRef] [Green Version]
- Thomas, X.; Campos, L.; Mounier, C.; Cornillon, J.; Flandrin, P.; Le, Q.H.; Piselli, S.; Guyotat, D. Expression of heat-shock proteins is associated with major adverse prognostic factors in acute myeloid leukemia. Leuk. Res. 2005, 29, 1049–1058. [Google Scholar] [CrossRef]
- Ghosh, J.C.; Siegelin, M.D.; Dohi, T.; Altieri, D.C. Heat shock protein 60 regulation of the mitochondrial permeability transition pore in tumor cells. Cancer Res. 2010, 70, 8988–8993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caruso Bavisotto, C.; Graziano, F.; Rappa, F.; Marino Gammazza, A.; Logozzi, M.; Fais, S.; Maugeri, R.; Bucchieri, F.; Conway de Macario, E.; Macario, A.J.L.; et al. Exosomal chaperones and miRNAs in gliomagenesis: State-of-art and theranostics perspectives. Int. J. Mol. Sci. 2018, 19, 2626. [Google Scholar] [CrossRef] [Green Version]
- Graziano, F.; Caruso Bavisotto, C.; Marino Gammazza, A.; Rappa, F.; Conway de Macario, E.; Macario, A.J.L.; Cappello, F.; Campanella, C.; Maugeri, R.; Iacopino, D.G. Chaperonology: The third eye on brain gliomas. Brain Sci. 2018, 8, 110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abu-Hadid, M.; Wilkes, J.D.; Elakawi, Z.; Pendyala, L.; Perez, R.P. Relationship between heat shock protein 60 (HSP6O) mRNA expression and resistance to platinum analogues in human ovarian and bladder carcinoma cell lines. Cancer Lett. 1997, 119, 63–70. [Google Scholar] [CrossRef]
- Tsai, Y.P.; Yang, M.H.; Huang, C.H.; Chang, S.Y.; Chen, P.M.; Liu, C.J.; Teng, S.C.; Wu, K.J. Interaction between HSP60 and β-catenin promotes metastasis. Carcinogenesis 2009, 30, 1049–1057. [Google Scholar] [CrossRef]
- Wu, J.; Liu, T.; Rios, Z.; Mei, Q.; Lin, X.; Cao, S. Heat Shock Proteins and Cancer. Trends Pharmacol. Sci. 2017, 38, 226–256. [Google Scholar] [CrossRef]
- Ruan, W.; Wang, Y.; Ma, Y.; Xing, X.; Lin, J.; Cui, J.; Lai, M. HSP60, a protein downregulated by IGFBP7 in colorectal carcinoma. J. Exp. Clin. Cancer Res. 2010, 29, 41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, H.; Li, J.; Liu, X.; Wang, G.; Luo, M.; Deng, H. Down-regulation of HSP60 Suppresses the Proliferation of Glioblastoma Cells via the ROS/AMPK/mTOR Pathway. Sci. Rep. 2016, 6, 28388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campanella, C.; D’Anneo, A.; Marino Gammazza, A.; Caruso Bavisotto, C.; Barone, R.; Emanuele, S.; Lo Cascio, F.; Mocciaro, E.; Fais, S.; Conway de Macario, E.; et al. The histone deacetylase inhibitor SAHA induces HSP60 nitration and its extracellular release by exosomal vesicles in human lung-derived carcinoma cells. Oncotarget 2016, 7, 28849–28867. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.; Sun, H.; Zheng, C.; Gao, J.; Fu, Q.; Hu, N.; Shao, X.; Zhou, Y.; Xiong, J.; Nie, K.; et al. Oncogenic HSP60 regulates mitochondrial oxidative phosphorylation to support Erk1/2 activation during pancreatic cancer cell growth. Cell Death Dis. 2018, 9, 161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caruso Bavisotto, C.; Marino Gammazza, A.; Lo Cascio, F.; Mocciaro, E.; Vitale, A.M.; Vergilio, G.; Pace, A.; Cappello, F.; Campanella, C.; Palumbo Piccionello, A. Curcumin affects Hsp60 folding activity and levels in neuroblastoma cells. Int. J. Mol. Sci. 2020, 21, 661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, W.; Ryu, J.; Kim, J.E. CCAR2/DBC1 and Hsp60 positively regulate expression of survivin in neuroblastoma cells. Int. J. Mol. Sci. 2019, 20, 131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaiwatanasirikul, K.A.; Sala, A. The tumour-suppressive function of CLU is explained by its localisation and interaction with HSP60. Cell Death Dis. 2011, 2, e219. [Google Scholar] [CrossRef] [Green Version]
- Campanella, C.; Bucchieri, F.; Ardizzone, N.M.; Marino Gammazza, A.; Montalbano, A.; Ribbene, A.; Di Felice, V.; Bellafiore, M.; David, S.; Rappa, F.; et al. Upon oxidative stress, the antiapoptotic Hsp60/procaspase-3 complex persists in mucoepidermoid carcinoma cells. Eur. J. Histochem. 2008, 52, 221–228. [Google Scholar] [CrossRef] [Green Version]
- Caruso Bavisotto, C.; Nikolic, D.; Marino Gammazza, A.; Barone, R.; Lo Cascio, F.; Mocciaro, E.; Zummo, G.; Conway de Macario, E.; Macario, A.J.L.; Cappello, F.; et al. The dissociation of the Hsp60/pro-Caspase-3 complex by bis(pyridyl)oxadiazole copper complex (CubipyOXA) leads to cell death in NCI-H292 cancer cells. J. Inorg. Biochem. 2017, 170, 8–16. [Google Scholar] [CrossRef] [Green Version]
- Tang, H.; Chen, Y.; Liu, X.; Wang, S.; Lv, Y.; Wu, D.; Wang, Q.; Luo, M.; Deng, H. Downregulation of HSP60 disrupts mitochondrial proteostasis to promote tumorigenesis and progression in clear cell renal cell carcinoma. Oncotarget 2016, 7, 38822–38834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Zhou, X.; Chang, H.; Huang, X.; Guo, X.; Du, X.; Tian, S.; Wang, L.; Lyv, Y.; Yuan, P.; et al. Hsp60 exerts a tumor suppressor function by inducing cell differentiation and inhibiting invasion in hepatocellular carcinoma. Oncotarget 2016, 7, 68976–68989. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cappello, F.; Di Stefano, A.; David, S.; Rappa, F.; Anzalone, R.; La Rocca, G.; D’Anna, S.E.; Magno, F.; Donner, C.F.; Balbi, B.; et al. Hsp60 and Hsp10 down-regulation predicts bronchial epithelial carcinogenesis in smokers with chronic obstructive pulmonary disease. Cancer 2006, 107, 2417–2424. [Google Scholar] [CrossRef]
- Hamelin, C.; Cornut, E.; Poirier, F.; Pons, S.; Beaulieu, C.; Charrier, J.P.; Haïdous, H.; Cotte, E.; Lambert, C.; Piard, F.; et al. Identification and verification of heat shock protein 60 as a potential serum marker for colorectal cancer. FEBS J. 2011, 278, 4845–4859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lv, L.H.; Wan, Y.L.; Lin, Y.; Zhang, W.; Yang, M.; Li, G.N.; Lin, H.M.; Shang, C.Z.; Chen, Y.J.; Min, J. Anticancer drugs cause release of exosomes with heat shock proteins from human hepatocellular carcinoma cells that elicit effective natural killer cell antitumor responses in vitro. J. Biol. Chem. 2012, 287, 15874–15885. [Google Scholar] [CrossRef] [Green Version]
- Lebret, T.; Watson, R.W.G.; Molinié, V.; O’Neill, A.; Gabriel, C.; Fitzpatrick, J.M.; Botto, H. Heat shock proteins HSP27, HSP60, HSP70, and HSP90: Expression in bladder carcinoma. Cancer 2003, 98, 970–977. [Google Scholar] [CrossRef]
- Campanella, C.; Rappa, F.; Sciumè, C.; Marino Gammazza, A.; Barone, R.; Bucchieri, F.; David, S.; Curcurù, G.; Caruso Bavisotto, C.; Pitruzzella, A.; et al. Heat shock protein 60 levels in tissue and circulating exosomes in human large bowel cancer before and after ablative surgery. Cancer 2015, 121, 3230–3239. [Google Scholar] [CrossRef]
- Théry, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K.; et al. Minimal information for studies of extracellular vesicles 2018 (MISEV2018): A position statement of the International Society for Extracellular Vesicles and update of the MISEV2014 guidelines. J. Extracell. Vesicles 2018, 7, 1535750. [Google Scholar] [CrossRef] [Green Version]
- Stoorvogel, W.; Kleijmeer, M.J.; Geuze, H.J.; Raposo, G. The biogenesis and functions of exosomes. Traffic 2002, 3, 321–330. [Google Scholar] [CrossRef]
- Camussi, G.; Deregibus, M.C.; Cantaluppi, V. Role of stem-cell-derived microvesicles in the paracrine action of stem cells. Biochem. Soc. Trans. 2013, 41, 283–287. [Google Scholar] [CrossRef]
- Liu, Q.; Rojas-Canales, D.M.; Divito, S.J.; Shufesky, W.J.; Stolz, D.B.; Erdos, G.; Sullivan, M.L.G.; Gibson, G.A.; Watkins, S.C.; Larregina, A.T.; et al. Donor dendritic cell-derived exosomes promote allograft-targeting immune response. J. Clin. Investig. 2016, 126, 2805–2820. [Google Scholar] [CrossRef] [PubMed]
- Long, Q.; Upadhya, D.; Hattiangady, B.; Kim, D.K.; An, S.Y.; Shuai, B.; Prockop, D.J.; Shetty, A.K. Intranasal MSC-derived A1-exosomes ease inflammation, and prevent abnormal neurogenesis and memory dysfunction after status epilepticus. Proc. Natl. Acad. Sci. USA 2017, 114, E3536–E3545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whiteside, T.L. Exosome and mesenchymal stem cell cross-talk in the tumor microenvironment. Semin. Immunol. 2018, 35, 69–79. [Google Scholar] [CrossRef] [PubMed]
- Tai, Y.L.; Chen, K.C.; Hsieh, J.T.; Shen, T.L. Exosomes in cancer development and clinical applications. Cancer Sci. 2018, 109, 2364–2374. [Google Scholar] [CrossRef] [Green Version]
- Caruso Bavisotto, C.; Marino Gammazza, A.; Rappa, F.; Fucarino, A.; Pitruzzella, A.; David, S.; Campanella, C. Exosomes: Can doctors still ignore their existence? EuroMediterr. Biomed. J. 2013, 8, 136–139. [Google Scholar]
- Vesiclepedia: Browse Results. Available online: http://microvesicles.org/browse_results?org_name=Homosapiens&cont_type=&tissue=&gene_symbol=&ves_type= (accessed on 10 January 2021).
- ExoCarta: Browse Results. Available online: http://exocarta.org/browse_results?org_name=Homosapiens&cont_type=&tissue=&gene_symbol= (accessed on 10 January 2021).
- Maacha, S.; Bhat, A.A.; Jimenez, L.; Raza, A.; Haris, M.; Uddin, S.; Grivel, J.C. Extracellular vesicles-mediated intercellular communication: Roles in the tumor microenvironment and anti-cancer drug resistance. Mol. Cancer 2019, 18, 55. [Google Scholar] [CrossRef] [Green Version]
- Dickman, C.T.; Lawson, J.; Jabalee, J.; MacLellan, S.A.; LePard, N.E.; Bennewith, K.L.; Garnis, C. Selective extracellular vesicle exclusion of miR-142-3p by oral cancer cells promotes both internal and extracellular malignant phenotypes. Oncotarget 2017, 8, 15252–15266. [Google Scholar] [CrossRef] [Green Version]
- Lawson, J.; Dickman, C.; MacLellan, S.; Towle, R.; Jabalee, J.; Lam, S.; Garnis, C. Selective secretion of microRNAs from lung cancer cells via extracellular vesicles promotes CAMK1D-mediated tube formation in endothelial cells. Oncotarget 2017, 8, 83913–83924. [Google Scholar] [CrossRef] [Green Version]
- Fais, S.; O’Driscoll, L.; Borras, F.E.; Buzas, E.; Camussi, G.; Cappello, F.; Carvalho, J.; Da Silva, A.C.; Del Portillo, H.; El Andaloussi, S.; et al. Evidence-Based Clinical Use of Nanoscale Extracellular Vesicles in Nanomedicine. ACS Nano 2016, 10, 3886–3899. [Google Scholar] [CrossRef]
- Campanella, C.; Caruso Bavisotto, C.; Logozzi, M.; Marino Gammazza, A.; Mizzoni, D.; Cappello, F.; Fais, S. On the Choice of the Extracellular Vesicles for Therapeutic Purposes. Int. J. Mol. Sci. 2019, 20, 236. [Google Scholar] [CrossRef] [Green Version]
- Caruso Bavisotto, C.; Cappello, F.; Macario, A.J.L.; Conway de Macario, E.; Logozzi, M.; Fais, S.; Campanella, C. Exosomal HSP60: A potentially useful biomarker for diagnosis, assessing prognosis, and monitoring response to treatment. Expert Rev. Mol. Diagn. 2017, 17, 815–822. [Google Scholar] [CrossRef] [PubMed]
- Bartel, D.P. MicroRNAs: Genomics, Biogenesis, Mechanism, and Function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef] [Green Version]
- Lewis, B.P.; Burge, C.B.; Bartel, D.P. Conserved seed pairing, often flanked by adenosines, indicates that thousands of human genes are microRNA targets. Cell 2005, 120, 15–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friedman, R.C.; Farh, K.K.H.; Burge, C.B.; Bartel, D.P. Most mammalian mRNAs are conserved targets of microRNAs. Genome Res. 2009, 19, 92–105. [Google Scholar] [CrossRef] [Green Version]
- Kumar, M.S.; Lu, J.; Mercer, K.L.; Golub, T.R.; Jacks, T. Impaired microRNA processing enhances cellular transformation and tumorigenesis. Nat. Genet. 2007, 39, 673–677. [Google Scholar] [CrossRef]
- Liu, M.; Zhu, K.; Qian, X.; Li, W. Identification of miRNA/mRNA-negative regulation pairs in nasopharyngeal carcinoma. Med. Sci. Monit. 2016, 22, 2215–2234. [Google Scholar] [CrossRef] [Green Version]
- Peng, Y.; Croce, C.M. The role of microRNAs in human cancer. Signal Transduct. Target. Ther. 2016, 1, 15004. [Google Scholar] [CrossRef] [Green Version]
- Svoronos, A.A.; Engelman, D.M.; Slack, F.J. OncomiR or tumor suppressor? The duplicity of MicroRNAs in cancer. Cancer Res. 2016, 76, 3666–3670. [Google Scholar] [CrossRef] [Green Version]
- Lan, H.; Lu, H.; Wang, X.; Jin, H. MicroRNAs as potential biomarkers in cancer: Opportunities and challenges. BioMed Res. Int. 2015, 2015, 125094. [Google Scholar] [CrossRef] [Green Version]
- Callegari, E.; Elamin, B.K.; Giannone, F.; Milazzo, M.; Altavilla, G.; Fornari, F.; Giacomelli, L.; D’Abundo, L.; Ferracin, M.; Bassi, C.; et al. Liver tumorigenicity promoted by microRNA-221 in a mouse transgenic model. Hepatology 2012, 56, 1025–1033. [Google Scholar] [CrossRef]
- Gwak, J.M.; Kim, H.J.; Kim, E.J.; Chung, Y.R.; Yun, S.; Seo, A.N.; Lee, H.J.; Park, S.Y. MicroRNA-9 is associated with epithelial-mesenchymal transition, breast cancer stem cell phenotype, and tumor progression in breast cancer. Breast Cancer Res. Treat. 2014, 147, 39–49. [Google Scholar] [CrossRef]
- Tanaka, R.; Tomosugi, M.; Horinaka, M.; Sowa, Y.; Sakai, T. Metformin causes G1-phase arrest via down-regulation of MIR-221 and enhances TRAIL sensitivity through DR5 up-regulation in pancreatic cancer cells. PLoS ONE 2015, 10, e0125779. [Google Scholar] [CrossRef] [Green Version]
- Munoz, J.L.; Rodriguez-Cruz, V.; Rameshwar, P. High expression of miR-9 in CD133 + glioblastoma cells in chemoresistance to temozolomide. J. Cancer Stem Cell Res. 2015, 3, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, C.-W.; Yu, J.-C.; Hsieh, Y.-H.; Liao, W.-L.; Shieh, J.-C.; Yao, C.-C.; Lee, H.-J.; Chen, P.-M.; Wu, P.-E.; Shen, C.-Y. Increased Cellular Levels of MicroRNA-9 and MicroRNA-221 Correlate with Cancer Stemness and Predict Poor Outcome in Human Breast Cancer. Cell. Physiol. Biochem. 2018, 48, 2205–2218. [Google Scholar] [CrossRef] [PubMed]
- Kao, C.J.; Martiniez, A.; Shi, X.B.; Yang, J.; Evans, C.P.; Dobi, A.; Devere White, R.W.; Kung, H.J. MiR-30 as a tumor suppressor connects EGF/Src signal to ERG and EMT. Oncogene 2014, 33, 2495–2503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ozgur, A.; Tutar, L.; Tutar, Y. Regulation of Heat Shock Proteins by miRNAs in Human Breast Cancer. MicroRNA 2014, 3, 118–135. [Google Scholar] [CrossRef]
- Choghaei, E.; Khamisipour, G.; Falahati, M.; Naeimi, B.; Mossahebi-Mohammadi, M.; Tahmasebi, R.; Hasanpour, M.; Shamsian, S.; Hashemi, Z.S. Knockdown of microRNA-29a changes the expression of heat shock proteins in breast carcinoma MCF-7 cells. Oncol. Res. 2016, 23, 69–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, W.; Liao, Y.; Li, Z.; Wang, Y.; Zheng, S.; Xu, X.; Ran, F.; Tang, B.; Wang, Z. MicroRNA-644a promotes apoptosis of hepatocellular carcinoma cells by downregulating the expression of heat shock factor 1. Cell Commun. Signal. 2018, 16, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ban, H.S.; Han, T.S.; Hur, K.; Cho, H.S. Epigenetic alterations of Heat Shock Proteins (HSPs) in cancer. Int. J. Mol. Sci. 2019, 20, 4758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shan, Z.X.; Lin, Q.X.; Deng, C.Y.; Zhu, J.N.; Mai, L.P.; Liu, J.L.; Fu, Y.H.; Liu, X.Y.; Li, Y.X.; Zhang, Y.Y.; et al. MiR-1/miR-206 regulate Hsp60 expression contributing to glucose-mediated apoptosis in cardiomyocytes. FEBS Lett. 2010, 584, 3592–3600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, Y.; Chen, X.; Li, X.; Li, Z.; Diao, H.; Liu, L.; Zhang, J.; Ju, J.; Wen, L.; Liu, X.; et al. MicroRNA-1 downregulation induced by carvedilol protects cardiomyocytes against apoptosis by targeting heat shock protein 60. Mol. Med. Rep. 2019, 49, 3527–3536. [Google Scholar] [CrossRef]
- Wang, L.L.; Dong, J.J.; An, B.Z.; Liang, J.; Cai, K.R.; Jin, Z.S.; Jin, H.S.; Hu, J.P. Has-miR-17 increases the malignancy of gastric lymphoma by HSP60/TNFR2 pathway. J. Biol. Regul. Homeost. Agents 2020, 34, 1317–1324. [Google Scholar] [PubMed]
- Ciocca, D.R.; Calderwood, S.K. Heat shock proteins in cancer: Diagnostic, prognostic, predictive, and treatment implications. Cell Stress Chaperones 2005, 10, 86–103. [Google Scholar] [CrossRef]
- Lianos, G.D.; Alexiou, G.A.; Mangano, A.; Mangano, A.; Rausei, S.; Boni, L.; Dionigi, G.; Roukos, D.H. The role of heat shock proteins in cancer. Cancer Lett. 2015, 360, 114–118. [Google Scholar] [CrossRef]
- Qian-Lin, Z.; Ting-Feng, W.; Qi-Feng, C.; Min-Hua, Z.; Ai-Guo, L. Inhibition of cytosolic chaperonin CCTζ-1 expression depletes proliferation of colorectal carcinoma in vitro. J. Surg. Oncol. 2010, 102, 419–423. [Google Scholar] [CrossRef]
- Dong, D.; Stapleton, C.; Luo, B.; Xiong, S.; Ye, W.; Zhang, Y.; Jhaveri, N.; Zhu, G.; Ye, R.; Liu, Z.; et al. A critical role for GRP78/BiP in the tumor microenvironment for neovascularization during tumor growth and metastasis. Cancer Res. 2011, 71, 2848–2857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Cui, S.; Zhang, X.; Wu, Y.; Tang, H. High Expression of Heat Shock Protein 90 Is Associated with Tumor Aggressiveness and Poor Prognosis in Patients with Advanced Gastric Cancer. PLoS ONE 2013, 8, e62876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jagadish, N.; Agarwal, S.; Gupta, N.; Fatima, R.; Devi, S.; Kumar, V.; Suri, V.; Kumar, R.; Suri, V.; Sadasukhi, T.C.; et al. Heat shock protein 70-2 (HSP70-2) overexpression in breast cancer. J. Exp. Clin. Cancer Res. 2016, 35, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.-Y.; Wei, P.-L.; Chen, W.-Y.; Chang, W.-C.; Chang, Y.-J. Silencing Heat Shock Protein 27 Inhibits the Progression and Metastasis of Colorectal Cancer (CRC) by Maintaining the Stability of Stromal Interaction Molecule 1 (STIM1) Proteins. Cells 2018, 7, 262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rappa, F.; Farina, F.; Zummo, G.; David, S.; Campanella, C.; Carini, F.; Tomasello, G.; Damiani, P.; Cappello, F.; Conway de Macario, E.; et al. HSP-molecular chaperones in cancer biogenesis and tumor therapy: An overview. AntiCancer Res. 2012, 32, 5139–5150. [Google Scholar]
- Chatterjee, S.; Burns, T.F. Targeting heat shock proteins in cancer: A promising therapeutic approach. Int. J. Mol. Sci. 2017, 18, 1978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Desmetz, C.; Bibeau, F.; Boissière, F.; Bellet, V.; Rouanet, P.; Maudelonde, T.; Mangé, A.; Solassol, J. Proteomics-based identification of HSP60 as a tumor-associated antigen in early-stage breast cancer and ductal carcinoma in situ. J. Proteome Res. 2008, 7, 3830–3837. [Google Scholar] [CrossRef] [PubMed]
- Abdalla, M.A.K.; Haj-Ahmad, Y. Promising urinary protein biomarkers for the early detection of hepato-cellular carcinoma among high-risk hepatitis C virus egyptian patients. J. Cancer 2012, 3, 390–403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, C.-L.; Hsu, Y.-T.; Wu, C.-C.; Yang, Y.-C.; Wang, C.; Wu, T.-C.; Hung, C.-F. Immune Mechanism of the Antitumor Effects Generated by Bortezomib. J. Immunol. 2012, 189, 3209–3220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murshid, A.; Gong, J.; Stevenson, M.A.; Calderwood, S.K. Heat shock proteins and cancer vaccines: Developments in the past decade and chaperoning in the decade to come. Expert Rev. Vaccines 2011, 10, 1553–1568. [Google Scholar] [CrossRef] [Green Version]
- Wiechmann, K.; Müller, H.; König, S.; Wielsch, N.; Svatoš, A.; Jauch, J.; Werz, O. Mitochondrial Chaperonin HSP60 Is the Apoptosis-Related Target for Myrtucommulone. Cell Chem. Biol. 2017, 24, 614–623.e6. [Google Scholar] [CrossRef] [Green Version]


| Chaperonopathies | Mechanism | Example | References |
|---|---|---|---|
| Excess | Quantitative variation, in which a gene is dysregulated or overexpressed. Qualitative variation, with a gain of function. | e.g., Alzheimer’s disease, Parkinson’s disease, Huntington’s disease. | [65] |
| Defect | Quantitative variation with gene downregulation. Qualitative variation, due to structural defect (genetic or acquired) | e.g., Charcot–Marie–Tooth disease, Spastic paraplegia, Hypo-myelinating leukodystrophy | [65] |
| Mistake or collaborationism | The chaperone is normal but the pathway in which it is involved may promote cell pathology. | e.g., certain tumor types, autoimmune conditions, prion disease | [65] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
David, S.; Vitale, A.M.; Fucarino, A.; Scalia, F.; Vergilio, G.; Conway de Macario, E.; Macario, A.J.L.; Caruso Bavisotto, C.; Pitruzzella, A. The Challenging Riddle about the Janus-Type Role of Hsp60 and Related Extracellular Vesicles and miRNAs in Carcinogenesis and the Promises of Its Solution. Appl. Sci. 2021, 11, 1175. https://doi.org/10.3390/app11031175
David S, Vitale AM, Fucarino A, Scalia F, Vergilio G, Conway de Macario E, Macario AJL, Caruso Bavisotto C, Pitruzzella A. The Challenging Riddle about the Janus-Type Role of Hsp60 and Related Extracellular Vesicles and miRNAs in Carcinogenesis and the Promises of Its Solution. Applied Sciences. 2021; 11(3):1175. https://doi.org/10.3390/app11031175
Chicago/Turabian StyleDavid, Sabrina, Alessandra Maria Vitale, Alberto Fucarino, Federica Scalia, Giuseppe Vergilio, Everly Conway de Macario, Alberto J. L. Macario, Celeste Caruso Bavisotto, and Alessandro Pitruzzella. 2021. "The Challenging Riddle about the Janus-Type Role of Hsp60 and Related Extracellular Vesicles and miRNAs in Carcinogenesis and the Promises of Its Solution" Applied Sciences 11, no. 3: 1175. https://doi.org/10.3390/app11031175